
MiR-30b Is Involved in the Homocysteine-Induced Apoptosis in Human Coronary Artery Endothelial Cells by Regulating the Expression of Caspase 3

Abstract
:1. Introduction
2. Results
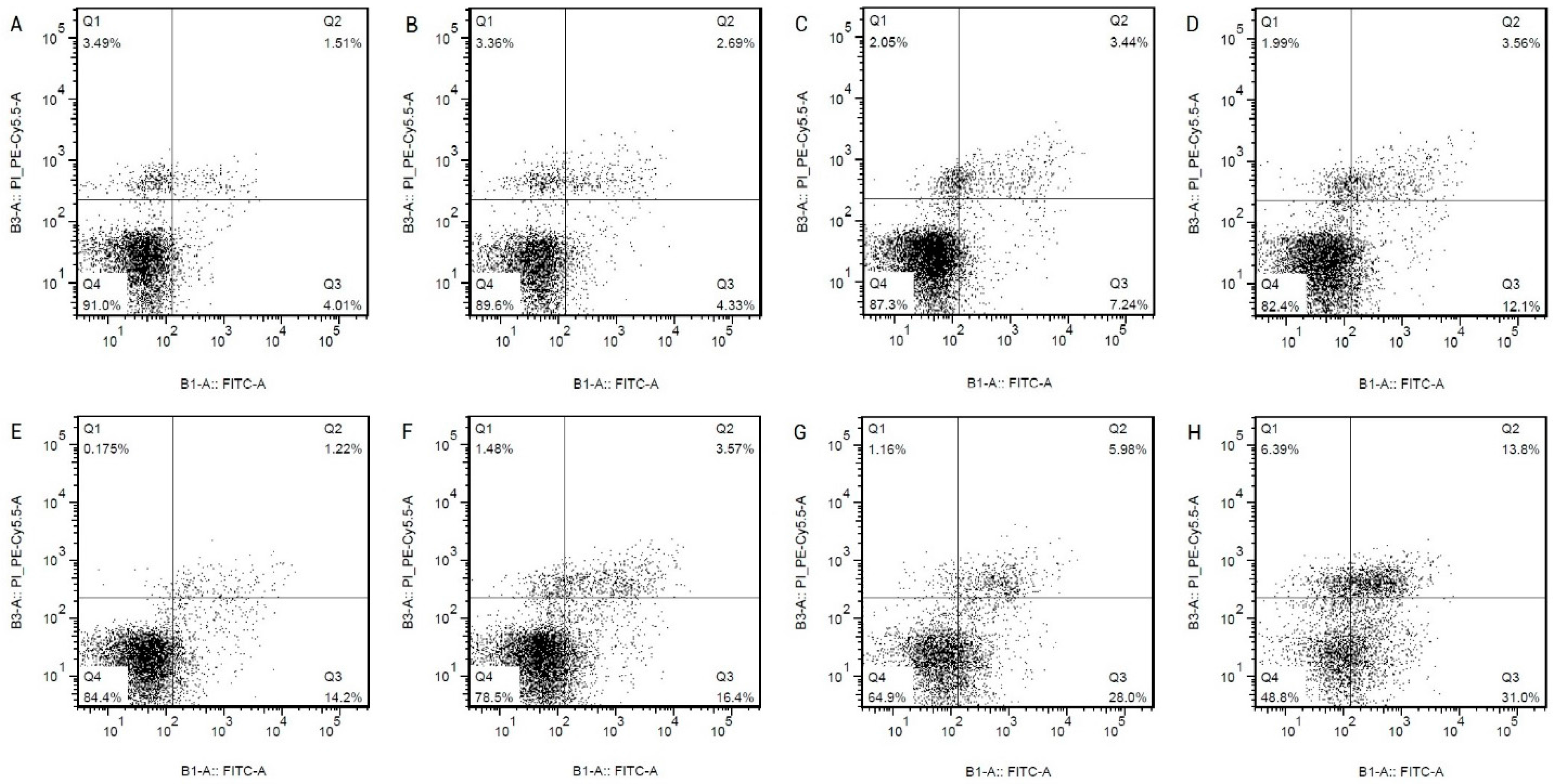
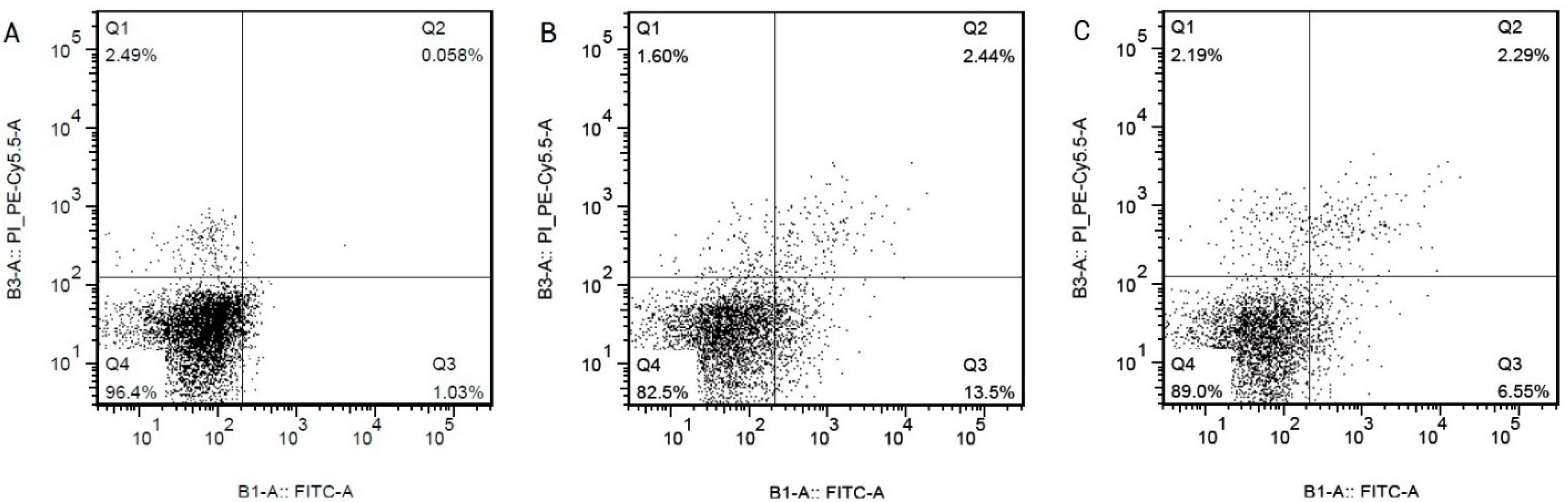
2.1. Hcy Induce Apoptosis in HCAECs by Upregulating the Expression of Caspase-3 in a Dose-Dependent Manner



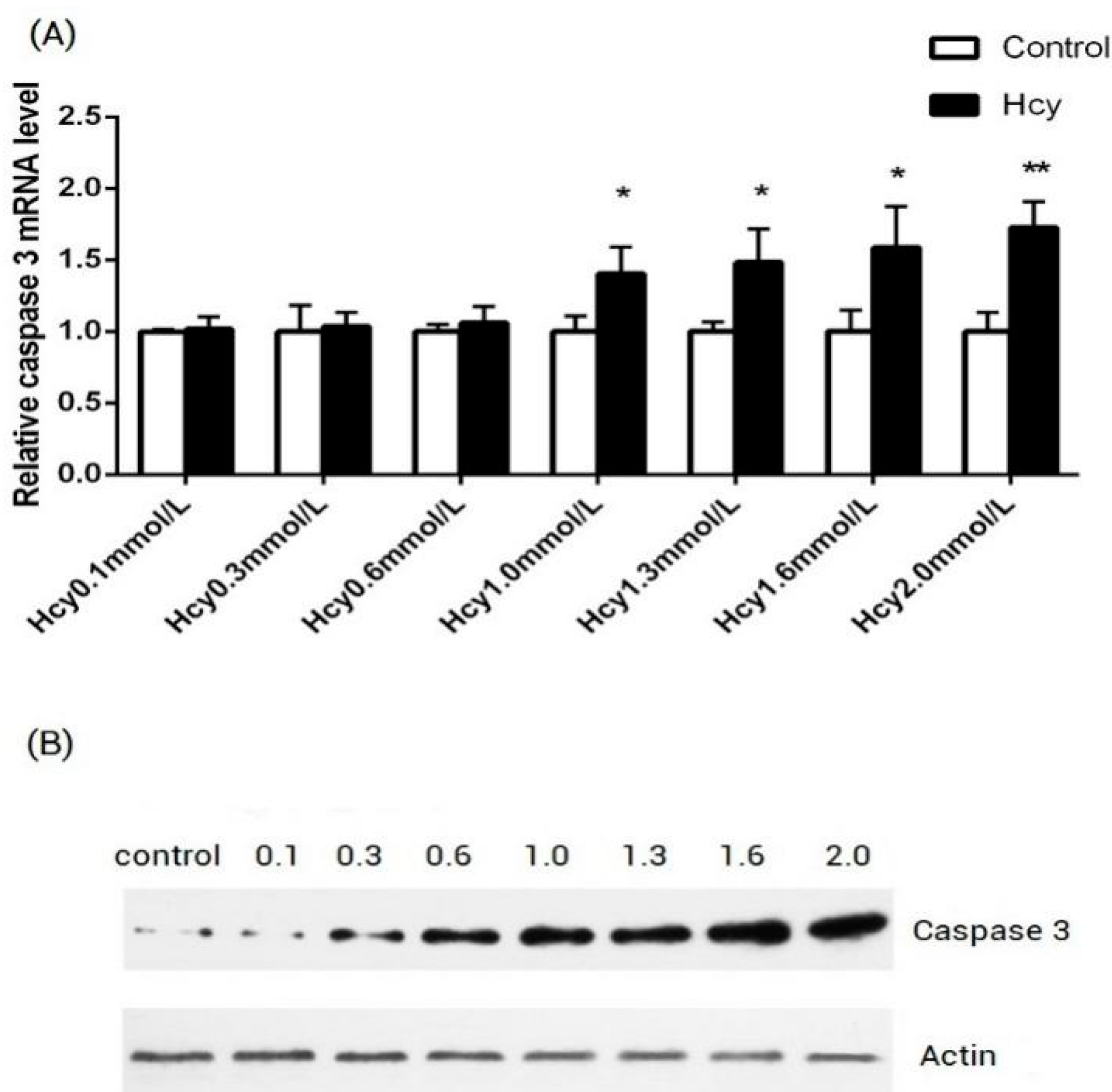{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (n = 3) | Apoptosis Rate % (LR + UR) |
|---|---|
| control | 5.92 ± 0.34 |
| Hcy 0.1 mmol/L | 6.5 ± 0.32 |
| Hcy 0.3 mmol/L | 9.88 ± 0.86 * |
| Hcy 0.6 mmol/L | 15.35 + 0.41 ** |
| Hcy 1.0 mmol/L | 16.28 ± 0.44 ** |
| Hcy 1.3 mmol/L | 19.72 ± 0.46 ** |
| Hcy 1.6 mmol/L | 32.9 ± 0.54 ** |
| Hcy 2.0 mmol/L | 48.76 ± 3.08 ** |
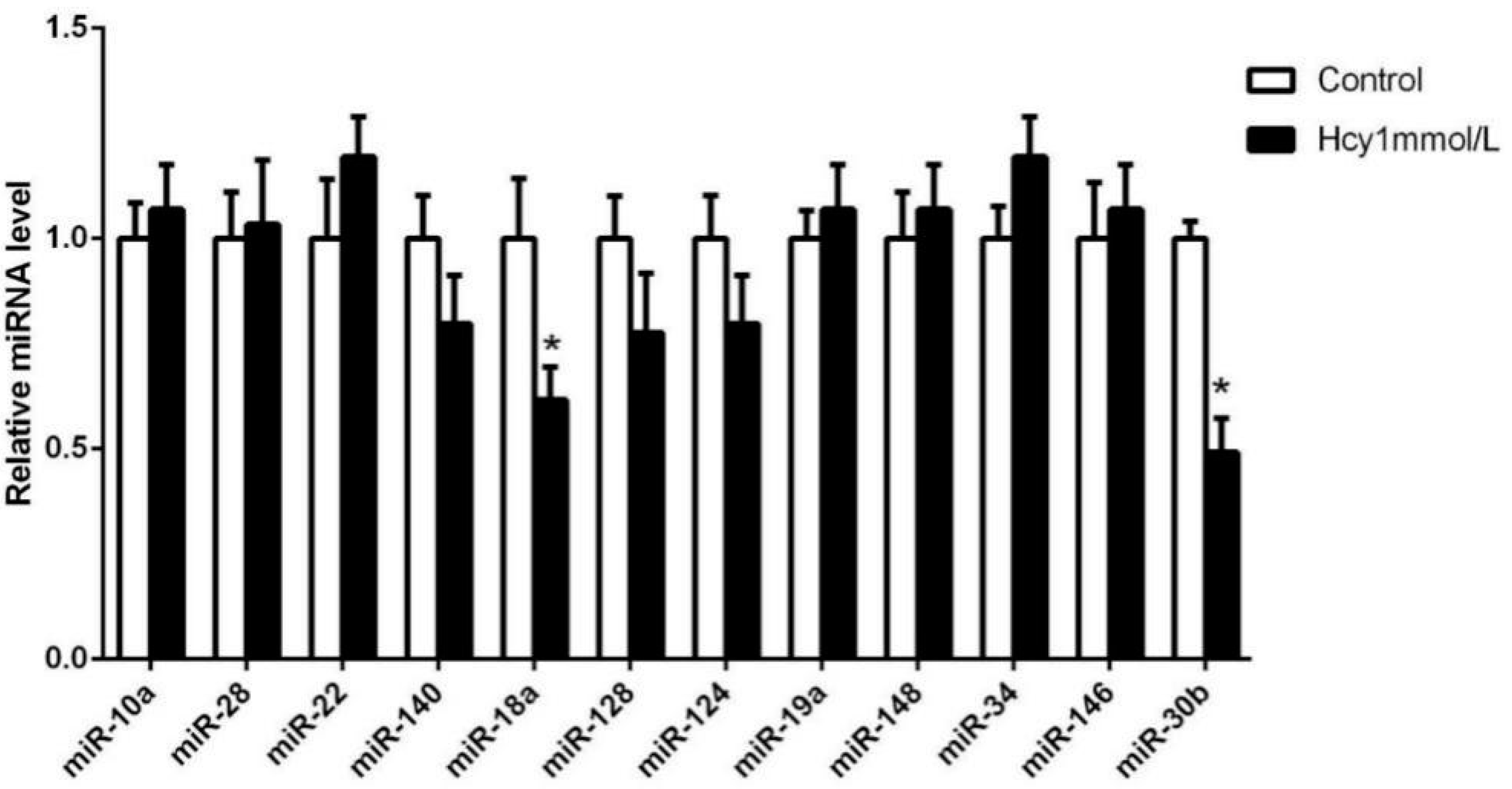
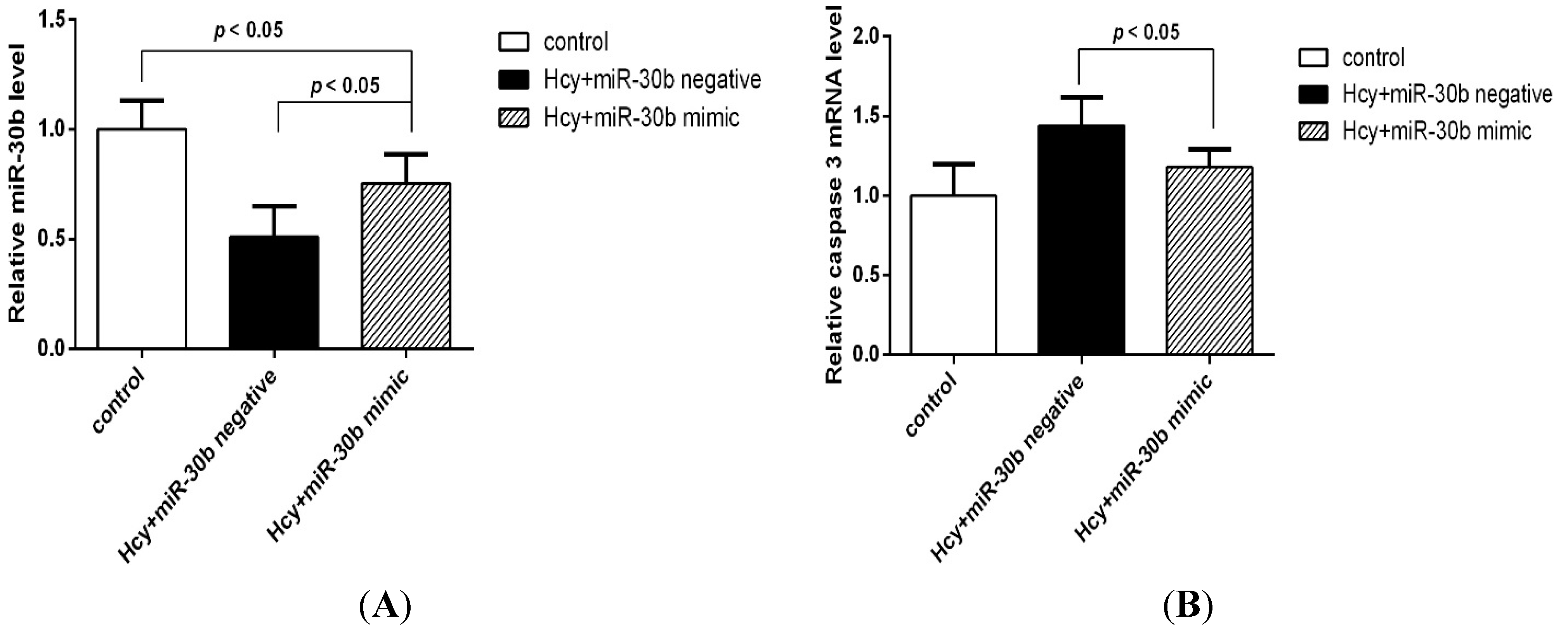
2.2. MiR-30b Is Downregulated during Hcy-Induced Apoptosis in HCAECs

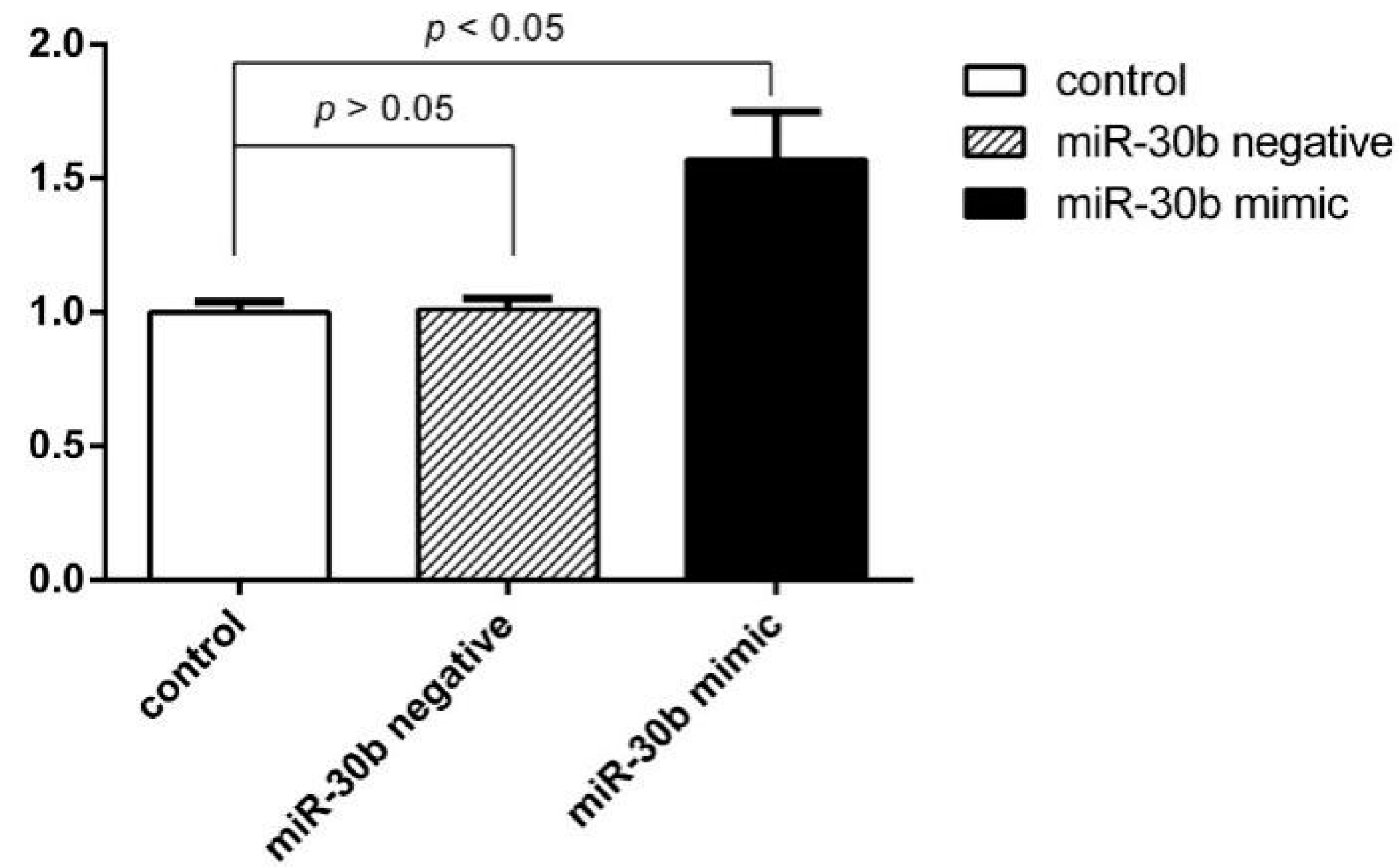
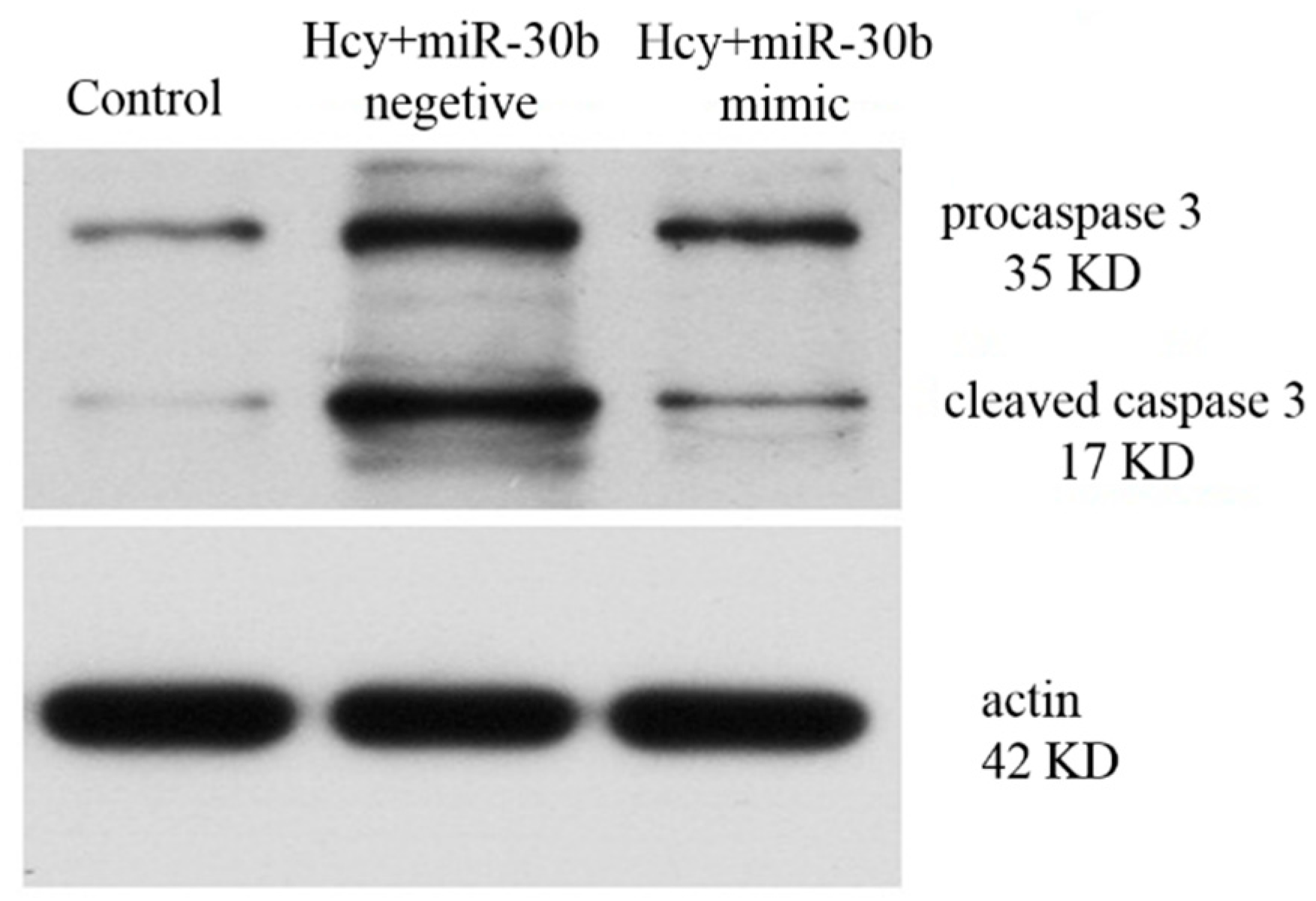
2.3. Overexpression of miR-30b Inhibits the Apoptosis Induced by Hcy in HCAECs


| Group (n = 4) | Apoptosis Rate % (LR + UR) |
|---|---|
| Control group | 4.77 ± 1.23 |
| Hcy negative control group | 15.83 ± 0.51 * |
| Hcy miR-30b overexpressed group | 10.59 ± 0.6 *,# |


3. Discussion
4. Experimental Section
4.1. Reagents and Cell Cultures
4.2. Apoptosis Assays
4.3. RNA Isolation, Reverse Transcription, Real-Time qPCR
| Primer Name | Primer Sequence |
|---|---|
| hsa-miR-10a-5p RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCACAAATT-3′ |
| hsa-miR-28-5P RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTCAAT-3′ |
| hsa-miR-22-5p RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTAAAGC-3′ |
| hsa-miR-140-5p RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTACCA-3′ |
| hsa-miR-18a-5p RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTATCT-3′ |
| hsa-miR-128 RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAGAG-3′ |
| hsa-miR-124a RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATAC-3′ |
| hsa-miR-19a RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAGTTTT-3′ |
| hsa-miR-148b RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACAAAGTT-3′ |
| hsa-miR-34b RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCAATCA-3′ |
| hsa-miR-146a RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAACCCA-3′ |
| hsa-miR-30b-5p RT primer | 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCTGA-3′ |
| Primer Name | Primer Sequence |
|---|---|
| hsa-miR-10a-5p Forward primer | 5′-ACGGGTACCCTGTAGATCCG-3′ |
| hsa-miR-28-5P Forward primer | 5′-GCCAAAGGAGCTCACAGTCT-3′ |
| hsa-miR-22-5p Forward primer | 5′-AGGCAGTTCTTCAGTGGCAA-3′ |
| hsa-miR-140-5p Forward primer | 5′-ACGGGCAGTGGTTTTACCCTA-3′ |
| hsa-miR-18a-5p Forward primer | 5′-AGGGCTAAGGTGCATCTAGTGC-3′ |
| hsa-miR-128 Forward primer | 5′-ATCCTTCACAGTGAACCGGT-3′ |
| hsa-miR-124 Forward primer | 5′-GTGAATTAAGGCACGCGGTG-3′ |
| hsa-miR-19a Forward primer | 5′-ACGCCTGTGCAAATCTATGC-3′ |
| hsa-miR-148b Forward primer | 5′-CGGCTCAGTGCATCACAG-3′ |
| hsa-miR-34b Forward primer | 5′-ACGGGTAGGCAGTGTCATTAGC-3′ |
| hsa-miR-146a Forward primer | 5′-AGCCGTGAGAACTGAATTCCA-3′ |
| hsa-miR-30b-5p Forward primer | 5′-ACGGGCTGTAAACATCCTACAC-3′ |
| has-miR Reverse primer | 5′-CAGTGCAGGGTCCGAGGTAT-4′ |
| caspase 3 Forward primer | 5′-TGTGAGGCGGTTGTGGAAGAGT-3′ |
| caspase 3 Reverse primer | 5′-AATGGGGGAAGAGGCAGGTGCA-3′ |
| U6 Forward primer | 5′-CTCGCTTCGGCAGCACA-3′ |
| U6 Reverse primer | 5′-AACGCTTCACGAATTTGCGT-3′ |
4.4. Western Blot
4.5. Overexpression of miR-30b
4.6. Statistical Methods
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schaffer, A.; Verdoia, M.; Cassetti, E.; Marino, P.; Suryapranata, H.; de Luca, G. Relationship between homocysteine and coronary artery disease. Results from a large prospective cohort study. Thromb. Res. 2014, 134, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Jia, X.; Gao, J.; Mou, W.; Tong, H.; Wen, X.; Tian, Y. Association of serum homocysteine levels with the severity and calcification of coronary atherosclerotic plaques detected by coronary CT angiography. Int. Angiol. 2014, 33, 316–323. [Google Scholar] [PubMed]
- Ma, Y.; Li, L.; Geng, X.B.; Hong, Y.; Shang, X.M.; Tan, Z.; Song, Y.X.; Zhao, G.Y.; Zhao, B.Q.; Tian, M.R. Correlation between hyperhomocysteinemia and outcomes of patients With acute myocardial infarction. Am. J. Ther. 2014. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, Y.; Chen, Y.; Feng, L.; Nie, Z. Homocysteine level and risk of different stroke types: A meta-analysis of prospective observational studies. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Pate, M.; Damarla, V.; Chi, D.S.; Negi, S.; Krishnaswamy, G. Endothelial cell biology: Role in the inflammatory response. Adv. Clin. Chem. 2010, 52, 109–130. [Google Scholar] [PubMed]
- Sipkens, J.A.; Hahn, N.; van den Brand, C.S.; Meischl, C.; Cillessen, S.A.; Smith, D.E.; Juffermans, L.J.; Musters, R.J.; Roos, D.; Jakobs, C.; et al. Homocysteine-induced apoptosis in endothelial cells coincides with nuclear NOX2 and peri-nuclear NOX4 activity. Cell. Biochem. Biophys. 2013, 67, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Hou, D.D.; Zhao, Q.; Liu, W.; Zhen, P.P.; Xu, J.P.; Wang, K.; Huang, H.X.; Li, X.; Zhang, H.; et al. Phytoestrogen ɑ-Zearalanol attenuates homocysteine-induced apoptosis in human umbilical vein endothelial cells. Biomed. Res. Int. 2013, 2013, 813450. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.J.; Lai, Y.Q.; Zhao, M.; Gong, T.; Zhang, B.K. Homocysteine-induced hypermethylation of DDAH2 promoter contributes to apoptosis of endothelial cells. Pharmazie 2013, 68, 282–286. [Google Scholar] [PubMed]
- Saki, N.; Abroun, S.; Soleimani, M.; Hajizamani, S.; Shahjahani, M.; Kast, R.E.; Mortazavi, Y. Involvement of MicroRNA in T-Cell Differentiation and Malignancy. Int. J. Hematol. Oncol. Stem Cell Res. 2015, 9, 33–49. [Google Scholar] [PubMed]
- Asada, S.; Takahashi, T.; Isodono, K.; Adachi, A.; Imoto, H.; Ogata, T.; Ueyama, T.; Matsubara, H.; Oh, H. Downregulation of Dicer expression by serum withdrawal sensitizes human endothelial cells to apoptosis. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2512–H2521. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pan, Q.; Zhao, Y.; He, C.; Bi, K.; Chen, Y.; Zhao, B.; Chen, Y.; Ma, X. MicroRNA-155 regulates ROS production, no generation, apoptosis and multiple functions of human brain microvessel endothelial cells under physiological and pathological conditions. J. Cell. Biochem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Cao, Y.; Yang, H.; Xiao, B.; Lu, Z. MicroRNA-221/222 regulate ox-LDL-induced endothelial apoptosis via Ets-1/p21 inhibition. Mol. Cell. Biochem. 2015, 405, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.L.; Zhang, Y.F.; Xia, Q.Q.; Zhu, J.; Yu, X.; Fan, T.; Wang, F. MicroRNA-19a regulates lipopolysaccharide-induced endothelial cell apoptosis through modulation of apoptosis signal-regulating kinase 1 expression. BMC Mol. Biol. 2015, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qin, W.; Zhang, L.; Wu, X.; Du, N.; Hu, Y.; Li, X.; Shen, N.; Xiao, D.; Zhang, H.; et al. MicroRNA-26a prevents endothelial cell apoptosis by directly targeting TRPC6 in the setting of atherosclerosis. Sci. Rep. 2015, 5, 9401. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, X.L.; Yan, C.H.; Li, Y.; Tian, X.X.; Zhu, N.; Rong, J.J.; Peng, C.F.; Han, Y.L. MicroRNA-495 regulates the proliferation and apoptosis of human umbilical vein endothelial cells by targeting chemokine CCL2. Thromb. Res. 2015, 135, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zhang, W.; Liu, Q. Human cytomegalovirus-encoded miR-US25–1 aggravates the oxidised low density lipoprotein-induced apoptosis of endothelial cells. Biomed. Res. Int. 2014, 2014, 531979. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Liang, H.; Liu, H.; Li, D.; Chen, X.; Li, L.; Zhang, C.Y.; Zen, K. Platelet-secreted microRNA-223 promotes endothelial cell apoptosis induced by advanced glycation end products via targeting the insulin-like growth factor 1 receptor. J. Immunol. 2014, 192, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, N.; Zhang, J.; Tong, Y. Hsa-let-7g miRNA targets caspase-3 and inhibits the apoptosis induced by ox-LDL in endothelial cells. Int. J. Mol. Sci. 2013, 14, 22708–22720. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.Q.; Xu, Z.M.; Xie, M.B.; Pei, D.A. Resveratrol inhibits hydrogen peroxide-induced apoptosis in endothelial cells via the activation of PI3K/Akt by miR-126. J. Atheroscler. Thromb. 2014, 21, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Xiong, Y.; Li, G.; Liu, M.; He, T.; Tang, Y.; Chen, Y.; Cai, L.; Jiang, R.; Tao, J. MiR-21 is overexpressed in response to high glucose and protects endothelial cells from apoptosis. Exp. Clin. Endocrinol. Diabetes 2013, 121, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Xiao, B.; Jiang, T.; Yang, H. Effects of miR-590–5p on ox-LDL-induced endothelial cells apoptosis and LOX-1 expression. J. Cent. South Univ. Med. Sci. 2012, 37, 675–681. [Google Scholar]
- Shin, S.; Moon, K.C.; Park, K.U.; Ha, E. MicroRNA-513a-5p mediates TNF-α and LPS induced apoptosis via downregulation of X-linked inhibitor of apoptotic protein in endothelial cells. Biochimie 2012, 94, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Xu, J.M.; Li, S.B.; Yuan, L.Q.; Dai, R.P. Effects of down-regulation of microRNA-23a on TNF-α-induced endothelial cell apoptosis through caspase-dependent pathways. Cardiovasc. Res. 2012, 93, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Xiao, B.; Liang, D.; Xia, J.; Li, Y.; Yang, H. MicroRNAs expression in ox-LDL treated HUVECs: MiR-365 modulates apoptosis and Bcl-2 expression. Biochem. Biophys. Res. Commun. 2011, 410, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Tyagi, N.; Kundu, S.; Tyagi, S.C. MicroRNAs are involved in homocysteine-induced cardiac remodeling. Cell Biochem. Biophys. 2009, 55, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kalani, A.; Kamat, P.K.; Tyagi, S.C.; Tyagi, N. Synergy of homocysteine, microRNA, and epigenetics: A novel therapeutic approach for stroke. Mol. Neurobiol. 2013, 48, 157–168. [Google Scholar] [CrossRef] [PubMed]
- McCully, K.S. Vascular pathology of homocysteinemia: Implications for the pathogenesis of arteriosclerosis. Am. J. Pathol. 1969, 56, 111–128. [Google Scholar] [PubMed]
- Kerkeni, M.; Tnani, M.; Chuniaud, L.; Miled, A.; Maaroufi, K.; Trivin, F. Comparative study on in vitro effects of homocysteine thiolactone and homocysteine on HUVEC cells: Evidence for a stronger proapoptotic and proinflammative homocysteine thiolactone. Mol. Cell. Biochem. 2006, 291, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Sun, Z.L.; Xie, Q.Y. Protective effect of microRNA-30b on hypoxia/reoxygenation-induced apoptosis in H9C2 cardiomyocytes. Gene 2015, 561, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Song, C.L.; Liu, B.; Wang, J.P.; Zhang, B.L.; Zhang, J.C.; Zhao, L.Y.; Shi, Y.F.; Li, Y.X.; Wang, G.; Diao, H.Y.; et al. Anti-apoptotic Effect of MicroRNA-30b in Early Phase of Rat Myocardial Ischemia-Reperfusion Injury Model. J. Cell. Biochem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhu, E.D.; Li, N.; Li, B.S.; Li, W.; Zhang, W.J.; Mao, X.H.; Guo, G.; Zou, Q.M.; Xiao, B. miR-30b, down-regulated in gastric cancer, promotes apoptosis and suppresses tumor growth by targeting plasminogen activator inhibitor-1. PLoS ONE 2014, 9, e106049. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.T.; Ye, Y.P.; Zhang, N.J.; Li, T.T.; Wang, S.Y.; Cui, Y.M.; Qi, L.; Wu, P.; Jiao, H.L.; Xie, Y.J.; et al. MicroRNA-30b functions as a tumour suppressor in human colorectal cancer by targeting KRAS, PIK3CD and BCL2. J. Pathol. 2014, 232, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Fujiya, M.; Konishi, H.; Mohamed Kamel, M.K.; Ueno, N.; Inaba, Y.; Moriichi, K.; Tanabe, H.; Ikuta, K.; Ohtake, T.; Kohgo, Y.; et al. MicroRNA-18a induces apoptosis in colon cancer cells via the autophagolysosomal degradation of oncogenic heterogeneous nuclear ribonucleoprotein A1. Oncogene 2014, 33, 4847–4856. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.I.; Hsu, C.H.; Lee, K.H.; Lin, J.T.; Chen, C.S.; Chang, K.C.; Su, C.Y.J.; Hsiao, M.; Lu, P.J. MicroRNA-18a is elevated in prostate cancer and promotes tumorigenesis through suppressing STK4 in vitro and in vivo. Oncogenesis 2014, 3, e99. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, P.; Zhao, W.; Liu, X.; Ma, J.; Xue, Y.; Liu, Y. MiR-18a regulates the proliferation, migration and invasion of human glioblastoma cell by targeting neogenin. Exp. Cell Res. 2014, 324, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Mercié, P.; Garnier, O.; Lascoste, L.; Renard, M.; Closse, C.; Durrieu, F.; Marit, G.; Boisseau, R.M.; Belloc, F. Homocysteine-thiolactone induces caspase-independent vascular endothelial cell death with apoptotic features. Apoptosis 2000, 5, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Gurda, D.; Handschuh, L.; Kotkowiak, W.; Jakubowski, H. Homocysteine thiolactone and N-homocysteinylated protein induce pro-atherogenic changes in gene expression in human vascular endothelial cells. Amino Acids 2015, 47, 1319–1339. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Chen, Q.; Song, X.; Zhou, L.; Zhang, J. MiR-30b Is Involved in the Homocysteine-Induced Apoptosis in Human Coronary Artery Endothelial Cells by Regulating the Expression of Caspase 3. Int. J. Mol. Sci. 2015, 16, 17682-17695. https://doi.org/10.3390/ijms160817682
Li F, Chen Q, Song X, Zhou L, Zhang J. MiR-30b Is Involved in the Homocysteine-Induced Apoptosis in Human Coronary Artery Endothelial Cells by Regulating the Expression of Caspase 3. International Journal of Molecular Sciences. 2015; 16(8):17682-17695. https://doi.org/10.3390/ijms160817682
Chicago/Turabian StyleLi, Feng, Qi Chen, Xiaowei Song, Lei Zhou, and Jianliang Zhang. 2015. "MiR-30b Is Involved in the Homocysteine-Induced Apoptosis in Human Coronary Artery Endothelial Cells by Regulating the Expression of Caspase 3" International Journal of Molecular Sciences 16, no. 8: 17682-17695. https://doi.org/10.3390/ijms160817682
APA StyleLi, F., Chen, Q., Song, X., Zhou, L., & Zhang, J. (2015). MiR-30b Is Involved in the Homocysteine-Induced Apoptosis in Human Coronary Artery Endothelial Cells by Regulating the Expression of Caspase 3. International Journal of Molecular Sciences, 16(8), 17682-17695. https://doi.org/10.3390/ijms160817682