
Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk


Abstract
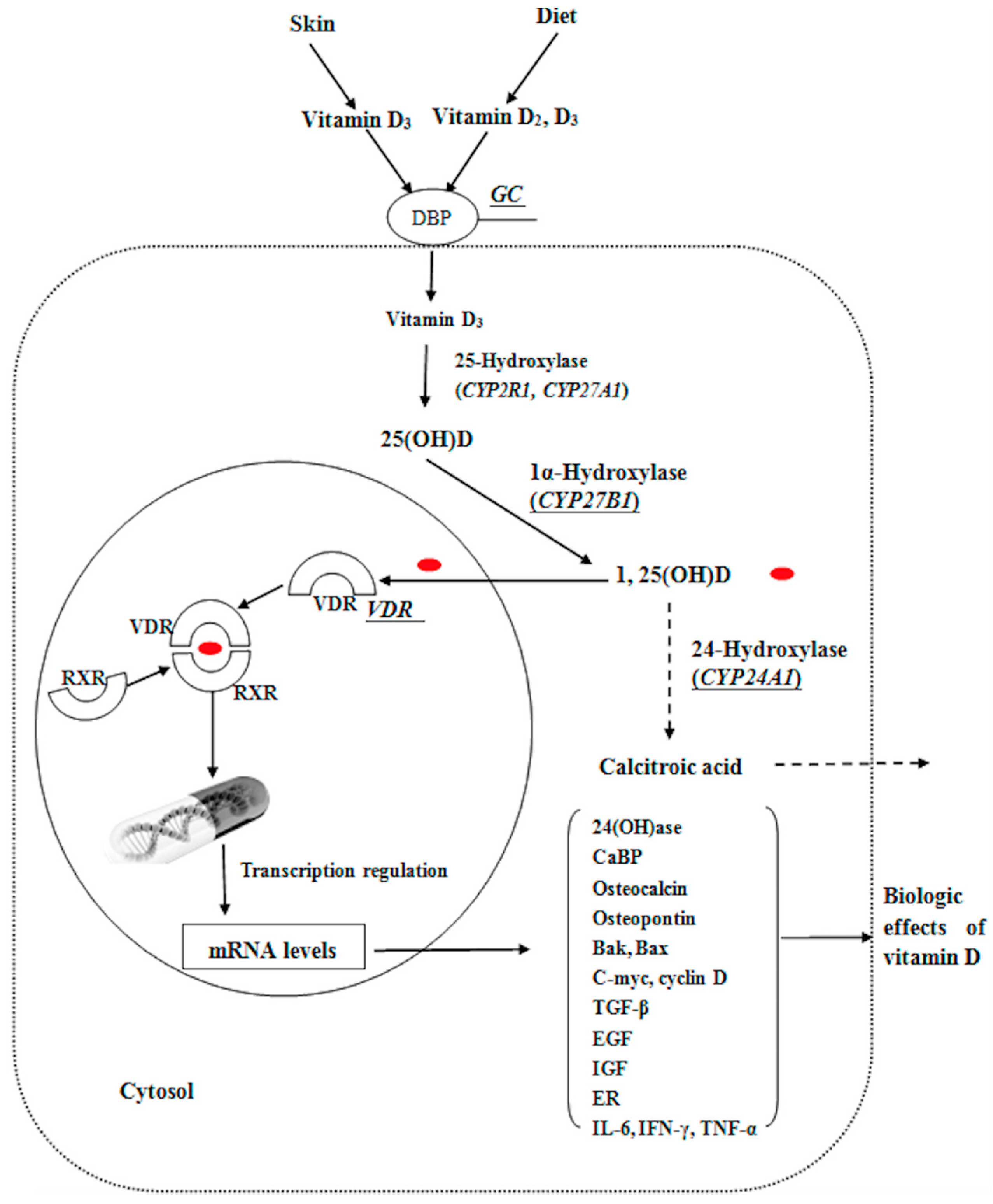
:1. Introduction
2. Results
2.1. Descriptive Characteristics
2.2. Relationship between SNPs, Smoking and NSCLC Risk
2.3. Plasma 25(OH)D Concentrations with Different Genotypes
2.4. Joint Association between Vitamin D-Related Polymorphisms and Plasma 25(OH)D on NSCLC Risk
2.5. Plasma 25(OH)D Concentrations in Smokers and Non-Smokers with Different Genotypes
2.6. Interaction between Gene and Smoking
3. Discussion
4. Materials and Methods
4.1. Case and Control Selection
4.2. Vitamin D Assay
4.3. SNP Selection and Genotyping Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA. Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.; Munden, R.F. Lung cancer epidemiology, risk factors, and prevention. Radiol. Clin. N. Am. 2012, 50, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Vaidya, V. Vitamin D and cancer. J. Cancer Res. Ther. 2007, 3, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D. The relationship between vitamin D and cancer. Clin. J. Oncol. Nurs. 2011, 15, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Trump, D.L.; Chadha, M.K.; Sunga, A.Y.; Fakih, M.G.; Ashraf, U.; Silliman, C.G.; Hollis, B.W.; Nesline, M.K.; Tian, L.; Tan, W.; et al. Vitamin D deficiency and insufficiency among patients with prostate cancer. BJU Int. 2009, 104, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Hines, S.L.; Jorn, H.K.; Thompson, K.M.; Larson, J.M. Breast cancer survivors and vitamin D: A review. Nutrition 2010, 26, 255–262. [Google Scholar] [CrossRef] [PubMed]
- LaPar, D.J.; Nagji, A.S.; Bhamidipati, C.M.; Kozower, B.D.; Lau, C.L.; Ailawadi, G.; Jones, D.R. Seasonal variation influences outcomes following lung cancer resections. Eur. J. Cardiothorac. Surg. 2011, 40, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D status: Measurement, interpretation, and clinical application. Ann. Epidemiol. 2009, 19, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Prosser, D.E.; Kaufmann, M. 25-Hydroxyvitamin d-24-hydroxylase (CYP24A1): Its important role in the degradation of vitamin D. Arch. Biochem. Biophys. 2012, 523, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Yin, X.Y.; Wu, X.H.; Zhang, L.; Pan, S.Y.; Zheng, Z.J.; Wang, J.G. Serum 25(OH)D and vitamin D status in relation to VDR, GC and CYP2R1 variants in Chinese. Endocr. J. 2014, 61, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, M.; Huang, G.; Delanghe, J.R.; Taes, Y.E. Biological and clinical aspects ofthe vitamin D binding protein (Gc-globulin) and its polymorphism. Clin. Chim. Acta 2006, 372, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Mawer, E.B.; Hayes, M.E.; Heys, S.E.; Davies, M.; White, A.; Stewart, M.F.; Smith, G.N. Constitutive synthesis of 1,25-dihydroxyvitamin D3 by a human small cell lung cancer cell line. J. Clin. Endocrinol. Metab. 1994, 79, 554–560. [Google Scholar] [PubMed]
- Zhou, W.; Heist, R.S.; Liu, G.; Asomaning, K.; Neuberg, D.S.; Hollis, B.W.; Wain, J.C.; Lynch, T.J.; Giovannucci, E.; Su, L.; et al. Circulating 25-hydroxyvitamin D levels predict survival in early-stage non-small-cell lung cancer patients. J. Clin. Oncol. 2007, 25, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, W.; Hu, Z.B.; Xu, L.; Shu, Y.Q.; Pan, S.Y.; Dai, J.C.; Jin, G.F.; Ma, H.X.; Shen, H.B.; et al. Plasma vitamin D levels and vitamin D receptor polymorphisms are associated with survival of non-small cell lung cancer. Chin. J. Cancer Res. 2011, 23, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Haddad, S.A.; Hu, Q.; Liu, S.; Lunetta, K.L.; Ruiz-Narvaez, E.A.; Hong, C.C.; Zhu, Q.; Sucheston-Campbell, L.; Cheng, T.Y.; et al. Genetic variations in vitamin D-related pathways and breast cancer risk in African American women in the AMBER consortium. Int. J. Cancer 2016, 138, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Arem, H.; Yu, K.; Xiong, X.; Moy, K.; Freedman, N.D.; Mayne, S.T.; Albanes, D.; Arslan, A.A.; Austin, M.; Bamlet, W.R.; et al. Vitamin D metabolic pathway genes and pancreatic cancer risk. PLoS ONE 2015, 10, e0117574. [Google Scholar] [CrossRef] [PubMed]
- Verone-Boyle, A.R.; Shoemaker, S.; Attwood, K.; Morrison, C.D.; Makowski, A.J.; Battaglia, S.; Hershberger, P.A. Diet-derived 25 hydroxyvitamin D3 activates vitamin D receptor target gene expression and suppresses EGFR mutant non-small cell lung cancer growth in vitro and in vivo. Oncotarget 2016, 7, 995–1013. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Goehe, R.W.; Di, X.; Hicks, M.A., 2nd; Torti, S.V.; Torti, F.M.; Harada, H.; Gewirtz, D.A. A novel cytostatic form of autophagy in sensitization of non-small cell lung cancer cells to radiation by vitamin D and the vitamin D analog, EB 1089. Autophagy 2014, 10, 2346–2361. [Google Scholar] [CrossRef] [PubMed]
- Turna, A.; Pekçolaklar, A.; Metin, M.; Yaylim, I.; Gurses, A. The effect of season of operation on the survival of patients with resected non-small cell lung cancer. Interact. Cardiovasc. Thorac. Surg. 2012, 14, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Porojnicu, A.C.; Robsahm, T.E.; Dahlback, A.; Berg, J.P.; Christiani, D.; Bruland, O.S.; Moan, J. Seasonal and geographical variations in lung cancer prognosis in Norway. Does vitamin D from the sun play a role? Lung Cancer 2007, 55, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kilkkinen, A.; Knekt, P.; Heliövaara, M.; Rissanen, H.; Marniemi, J.; Hakulinen, T.; Aromaa, A. Vitamin D status and the risk of lung cancer: A cohort study in Finland. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3274–3278. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J. Vitamin D deficiency in patients with inflammatory bowel disease. Br. J. Nurs. 2016, 25, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Brot, C.; Jorgensen, N.R.; Sorensen, O.H. The influence of smoking on vitamin D status and calcium metabolism. Eur. J. Clin. Nutr. 1999, 53, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Saleh, F.; Figenschau, Y.; Kamycheva, E.; Haug, E.; Sundsfjord, J. Serum parathyroid hormone (PTH) levels in smokers and non- smokers. The fifth Tromso study. Eur. J. Endocrinol. 2005, 152, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Need, A.G.; Kemp, A.; Giles, N.; Morris, H.A.; Horowitz, M.; Nordin, B.E. Relationships between intestinal calcium absorption, serum vitamin D metabolites and smoking in postmenopausal women. Osteoporos. Int. 2002, 13, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Stampfli, M.R.; Anderson, G.P. How cigarette smoke skews immune responses to promote infection, lung disease and cancer. Nat. Rev. Immunol. 2009, 9, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Mio, T.; Romberger, D.J.; Thompson, A.B.; Robbins, R.A.; Heires, A.; Rennard, S.I. Cigarette smoke induces interleukin-8 release from human bronchial epithelial cells. Am. J. Respir. Crit. Care Med. 1997, 155, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Hansdottir, S. Modulation of Lung Innate Immunity by Vitamin D and Cigarette Smoke. Ph.D. Thesis, University of Iceland, Reykjavík, Iceland, 2011; pp. 36–65. [Google Scholar]
- Haley, K.J.; Manoli, S.E.; Tantisira, K.G.; Litonjua, A.A.; Nguyen, P.; Kobzik, L.; Weiss, S.T. Maternal smoking causes abnormal expression of the vitamin D receptor. Am. J. Respir. Crit. Care Med. 2009, 179, A5874. [Google Scholar]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef]
- Joshi, A.D.; Andersson, C.; Buch, S.; Stender, S.; Noordam, R.; Weng, L.C.; Weeke, P.E.; Auer, P.L.; Boehm, B.; Chen, C.; et al. Four susceptibility loci for gallstone disease identified in a meta-analysis of genome-wide association studies. Gastroenterology 2016, 151, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Cheng, Y.; Dai, J.; Xie, L.; Jin, G.; Ma, H.; Hu, Z.; Shi, Y.; Lin, D.; Shen, H. Genome-wide association study based risk prediction model in predicting lung cancer risk in Chinese. Zhonghua Liu Xing Bing Xue Za Zhi 2015, 36, 1047–1052. [Google Scholar] [PubMed]
- Cargill, M.; Altshuler, D.; Ireland, J.; Sklar, P.; Ardlie, K.; Patil, N.; Shaw, N.; Lane, C.R.; Lim, E.P.; Kalyanaraman, N.; et al. Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat. Genet. 1999, 22, 231–238. [Google Scholar] [PubMed]
- Chen, G.; Kim, S.H.; King, A.N.; Zhao, L.; Simpson, R.U.; Christensen, P.J.; Wang, Z.; Thomas, D.G.; Giordano, T.J.; Lin, L.; et al. CYP24A1 is an independent prognostic marker of survival in patients with lung adenocarcinoma. Clin. Cancer Res. 2011, 17, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, N.; Nadal, E.; Jeon, C.K.; Sandoval, J.; Colacino, J.; Rozek, L.S.; Christensen, P.J.; Esteller, M.; Beer, D.G.; Kim, S.H. Epigenetic regulation of vitamin D metabolism in human lung adenocarcinoma. J. Thorac. Oncol. 2014, 9, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kanterewicz, B.; Buch, S.; Petkovich, M.; Parise, R.; Beumer, J.; Lin, Y.; Diergaarde, B.; Hershberger, P.A. CYP24 inhibition preserves 1α,25-dihydroxyvitamin D(3) anti-proliferative signaling in lung cancer cells. Mol. Cell. Endocrinol. 2012, 355, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Ramshaw, H.; Zhang, A.; Cook, R.; Byford, V.; White, J.; Petkovich, M. Expression and activity of vitamin D-metabolizing cytochrome P450s (CYP1alpha and CYP24) in human nonsmall cell lung carcinomas. Endocrinology 1999, 140, 3303–3310. [Google Scholar] [PubMed]
- Anderson, M.G.; Nakane, M.; Ruan, X.; Kroeger, P.E.; Wu-Wong, J.R. Expression of VDR and CYP24A1 mRNA in human tumors. Cancer Chemother. Pharmacol. 2006, 57, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, X.; Chen, X.; Liu, L.; Lu, C.; Tang, X.; Shi, J.; Li, M.; Zhou, M.; Zhang, Z.; et al. GC Glu416Asp and Thr420Lys polymorphisms contribute to gastrointestinal cancer susceptibility in a Chinese population. Int. J. Clin. Exp. Med. 2012, 5, 72–79. [Google Scholar] [PubMed]
- Hummel, D.M.; Fetahu, I.S.; Gröschel, C.; Manhardt, T.; Kállay, E. Role of proinflammatory cytokines on expression of vitamin D metabolism and target genes incolon cancer cells. J. Steroid Biochem. Mol. Biol. 2014, 144, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Shui, I.M.; Mondul, A.M.; Lindström, S.; Tsilidis, K.K.; Travism, R.C.; Gerke, T.; Albanes, D.; Mucci, L.A.; Giovannucci, E.; Kraft, P. Breast and prostate cancer cohort consortium group: Circulating vitamin D, vitamin D-related genetic variation, and risk of fatal prostate cancer in the national cancer institute breast and prostate cancer cohort consortium. Cancer 2015, 121, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.; Bonilla, C.; Metcalfe, C.; Lewis, S.; Evans, D.M.; Fraser, W.D.; Kemp, J.P.; Donovan, J.L.; Hamdy, F.C.; Neal, D.E.; et al. Associations of vitamin D pathway genes with circulating 25-hydroxyvitamin-D, 1,25-dihydroxyvitamin-D, and prostate cancer: A nested case-control study. Cancer Causes Control 2015, 26, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, A.K.; Bairati, I.; Qiu, X.; Huang, H.; Cheng, D.; Liu, G.; Meyer, F.; Adjei, A.; Xu, W. Genetic sequence variants in vitamin D metabolism pathway genes, serum vitamin D level andoutcome in head and neck cancer patients. Int. J. Cancer 2013, 132, 2520–2527. [Google Scholar] [CrossRef] [PubMed]
- Woodson, K.; Ratnasinghe, D.; Bhat, N.K.; Stewart, C.; Tangrea, J.A.; Hartman, T.J.; Stolzenberg-Solomon, R.; Virtamo, J.; Taylor, P.R.; Albanes, D. Prevalence of disease-related DNA polymorphisms among participants in a large cancer prevention trial. Eur. J. Cancer Prev. 1999, 8, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, N.; Daignault-Newton, S.; Dy, G.K.; Muindi, J.R.; Adjei, A.; Elingrod, V.L.; Kalemkerian, G.P.; Cease, K.B.; Stella, P.J.; Brenner, D.E.; et al. A phase I/II pharmacokinetic and pharmacogenomic study of calcitriol in combination with cisplatin and docetaxel in advanced non-small-cell lung cancer. Cancer Chemother. Pharmacol. 2013, 71, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Xu, F.; Qu, J.; Wang, Y.; Gao, M.; Yu, H.; Qian, B. Genetic polymorphisms in the vitamin D pathway in relation to lung cancer risk and survival. Oncotarget 2015, 6, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.L.; Lau, K.S.; Sham, P.C.; Tan, K.C.; Kung, A.W. Genetic variant in vitamin D binding protein is associated with serum 25-hydroxyvitamin D and vitamin D insufficiency in southern Chinese. J. Hum. Genet. 2013, 58, 749–751. [Google Scholar] [CrossRef] [PubMed]
- Anic, G.M.; Thompson, R.C.; Nabors, L.B.; Olson, J.J.; Browning, J.E.; Madden, M.H.; Murtagh, F.R.; Forsyth, P.A.; Egan, K.M. An exploratory analysis of common genetic variants in the vitamin D pathway including genome-wide associated variants in relation to glioma risk and outcome. Cancer Causes Control 2012, 23, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejuk, D.; Łaczmański, Ł.; Milewicz, A.; Kuliczkowska-Płaksej, J.; Lenarcik-Kabza, A.; Hirnle, L.; Zaleska-Dorobisz, U.; Lwow, F. Classic PCOS phenotype is not associated with deficiency of endogenous vitamin D and VDR gene polymorphisms rs731236 (Taq1), rs7975232 (Apa1), rs1544410 (Bsm1), rs10735810 (Fok1): A case-control study of lower Silesian women. Gynecol. Endocrinol. 2015, 30, 1–4. [Google Scholar]
- Kaabachi, W.; Kaabachi, S.; Rafrafi, A.; Amor, A.B.; Tizaoui, K.; Haj Sassi, F.; Hamzaoui, K. Association of vitamin D receptor Fok1 and Apa1 polymorphisms with lung cancer risk in Tunisian population. Mol. Biol. Rep. 2014, 41, 6545–6553. [Google Scholar] [CrossRef] [PubMed]
- Dogan, I.; Onen, H.I.; Yurdakul, A.S.; Konac, E.; Ozturk, C.; Varol, A.; Ekmekci, A. Polymorphisms in the vitamin D receptor gene and risk of lung cancer. Med. Sci. Monit. 2009, 15, BR232–BR242. [Google Scholar] [PubMed]
- Xiong, L.; Cheng, J.; Gao, J.; Wang, J.; Liu, X.; Wang, L. Vitamin D receptor genetic variants are associated with chemotherapy response and prognosis in patients with advanced non-small-cell lung cancer. Clin. Lung Cancer 2013, 14, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, J.; Zhang, Y. Polymorphisms in the vitamin D receptor gene and the lung cancer risk. Tumour Biol. 2014, 35, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Heist, R.S.; Zhou, W.; Wang, Z.; Liu, G.; Neuberg, D.; Su, L.; Asomaning, K.; Hollis, B.W.; Lynch, T.J.; Wain, J.C.; et al. Circulating 25-hydroxyvitamin D, VDR polymorphisms, and survival in advanced non-small-cell lung cancer. J. Clin. Oncol. 2008, 26, 5596–5602. [Google Scholar] [CrossRef] [PubMed]
- Ala-Houhala, M.J.; Vähävihu, K.; Snellman, E.; Hasan, T.; Kautiainen, H.; Karisola, P.; Dombrowski, Y.; Schauber, J.; Saha, H.; Reunala, T. A narrow-band ultraviolet B course improves vitamin D balance and alters cutaneous CYP27A1 and CYP27B1 mRNA expression levels in haemodialysis patients supplemented with oral vitamin D. Nephron. Clin. Pract. 2013, 124, 17–22. [Google Scholar]
- Jóźwicki, W.; Brożyna, A.A.; Siekiera, J.; Slominski, A.T. Expression of vitamin D receptor (VDR) positively correlates with survival of urothelial bladder cancer patients. Int. J. Mol. Sci. 2015, 16, 24369–24386. [Google Scholar] [CrossRef] [PubMed]
- Young, M.R.; Ihm, J.; Lozano, Y.; Wright, M.A.; Prechel, M.M. Treating tumor-bearing mice with vitamin D3 diminishes tumor-induced myelopoiesis and associated immunosuppression, and reduces tumor metastasis and recurrence. Cancer Immunol. Immunother. 1995, 41, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Kawar, N.; Maclaughlan, S.; Horan, T.C.; Uzun, A.; Lange, T.S.; Kim, K.K.; Hopson, R.; Singh, A.P.; Sidhu, P.S.; Glass, K.A.; et al. PT19c, another nonhypercalcemic vitamin D2 derivative, demonstrates antitumor efficacy in epithelial ovarian and endometrial cancer models. Genes Cancer 2013, 4, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Thun, M.J.; Hannan, L.M.; Adams-Campbell, L.L.; Boffetta, P.; Buring, J.E.; Feskanich, D.; Flanders, W.D.; Jee, S.H.; Katanoda, K.; Kolonel, L.N.; et al. Lung cancer occurrence in never-smokers: An analysis of 13 cohorts and 22 cancer registry studies. PLoS Med. 2008, 5, e185. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, S.; Hiraku, Y.; Oikawa, S. Mechanism of guanine-specific DNA damage by oxidative stress and its role in carcinogenesis and aging. Mutat. Res. 2001, 488, 65–76. [Google Scholar] [CrossRef]
- Pastoriza Gallego, M.; Sarasin, A. Transcription-coupled repair of 8-oxoguanine in human cells and its deficiency in some DNA repair diseases. Biochimie 2003, 85, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.H.; Jaakkola, J.J.; Chuang, C.Y.; Liou, S.H.; Lung, S.C.; Loh, C.H.; Yu, D.S.; Strickland, P.T. Exposure to cooking oil fumes and oxidative damages: A longitudinal study in Chinese military cooks. J. Expo. Sci. Environ. Epidemiol. 2013, 23, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Fry, J.S.; Lee, P.N.; Forey, B.A.; Coombs, K.J. How rapidly does the excess risk of lung cancer decline following quitting smoking? A quantitative review using the negative exponential model. Regul. Toxicol. Pharmacol. 2013, 67, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Maneechay, W.; Boonpipattanapong, T.; Kanngurn, S.; Puttawibul, P.; Geater, S.L.; Sangkhathat, S. Single nucleotide polymorphisms in the GC gene for vitamin D binding protein in common cancers in Thailand. Asian Pac. J. Cancer Prev. 2015, 16, 3339–3344. [Google Scholar] [CrossRef] [PubMed]
- Anic, G.M.; Weinstein, S.J.; Mondul, A.M.; Männistö, S.; Albanes, D. Serum vitamin D, vitamin D binding protein, and lung cancer survival. Lung Cancer 2014, 86, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Eliassen, A.H.; Spiegelman, D.; Willett, W.C.; Hankinson, S.E. Plasma free 25-hydroxyvitamin D, vitamin D binding protein, and risk of breast cancer in the Nurses’ Health Study II. Cancer Causes Control 2014, 25, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Pande, M.; Thompson, P.A.; Do, K.A.; Sahin, A.A.; Amos, C.I.; Frazier, M.L.; Bondy, M.L.; Brewster, A.M. Genetic variants in the vitamin D pathway and breast cancer disease-free survival. Carcinogenesis 2013, 34, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Piper, M.R.; Freedman, D.M.; Robien, K.; Kopp, W.; Rager, H.; Horst, R.L.; Stolzenberg-Solomon, R.Z. Vitamin D-binding protein and pancreatic cancer: A nested case-control study. Am. J. Clin. Nutr. 2015, 101, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Poynter, J.N.; Jacobs, E.T.; Figueiredo, J.C.; Lee, W.H.; Conti, D.V.; Campbell, P.T.; Levine, A.J.; Limburg, P.; Le Marchand, L.; Cotterchio, M.; et al. Genetic variation in the vitamin D receptor (VDR) and the vitamin D-binding protein (GC) and risk for colorectal cancer: Results from the colon cancer family registry. Cancer Epidemiol. Biomark. Prev. 2010, 19, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Flohil, S.C.; de Vries, E.; van Meurs, J.B.; Fang, Y.; Stricker, B.H.; Uitterlinden, A.G.; Nijsten, T. Vitamin D-binding protein polymorphisms are not associated with development of (multiple) basal cell carcinomas. Exp. Dermatol. 2010, 19, 1103–1105. [Google Scholar] [CrossRef] [PubMed]
- Leslie, H.S.; Mary, K.G.; Christian, W. TNM Classification of Malignant Tumours, 7th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 136–138. [Google Scholar]
- Wang, X.; Cui, J.; Gu, J.; He, H.; Li, B.; Li, W. Plasma 25-hydroxyvitamin D deficiency is associated with the risk of non-small cell lung cancer in a Chinese population. Cancer Biomark. 2015, 15, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Schaap, M.M.; Kunst, A.E.; Leinsalu, M.; Regidor, E.; Ekholm, O.; Dzurova, D.; Helmert, U.; Klumbiene, J.; Santana, P.; Mackenbach, J.P. Effect of nationwide tobacco control policies on smoking cessation in high and low educated groups in 18 European countries. Tob. Control 2008, 17, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Hanwell, H.E.; Vieth, R. An evaluation of automated methods for measurement of serum 25-hydroxyvitamin D. Clin. Biochem. 2009, 42, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.S.; Eberle, M.A.; Rieder, M.J.; Yi, Q.; Kruglyak, L.; Nickerson, D.A. Selecting a maximally informative set of single-nucleotide polymorphisms for association analyses using linkage disequilibrium. Am. J. Hum. Genet. 2004, 74, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Hu, Z.; Wu, C.; Guo, H.; Zhou, B.; Lv, J.; Lu, D.; Chen, K.; Shi, Y.; Chu, M.; et al. Association analyses identify multiple new lung cancer susceptibility loci and their interactions with smoking in the Chinese population. Nat. Genet. 2012, 44, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, H.; Tao, H.; Suzuki, M.; Mori, H.; Tsuboi, M.; Matsuura, S.; Goto, M.; Shinmura, K.; Ozawa, T.; Tanioka, F.; et al. Genetic susceptibility to lung cancer. Front. Biosci. 2011, 3, 1463–1477. [Google Scholar] [CrossRef]
- Reimers, L.L.; Crew, K.D.; Bradshaw, P.T.; Santella, R.M.; Steck, S.E.; Sirosh, I.; Terry, M.B.; Hershman, D.L.; Shane, E.; Cremers, S.; et al. Vitamin D-related gene polymorphisms, plasma 25-hydroxyvitamin D, and breast cancer risk. Cancer Causes Control 2015, 26, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Romanos, J.; van Diemen, C.C.; Nolte, I.M.; Trynka, G.; Zhernakova, A.; Fu, J.; Bardella, M.T.; Barisani, D.; McManus, R.; van Heel, D.A.; et al. Analysis of HLA and non-HLA alleles can identify individuals at high risk for celiac disease. Gastroenterology 2009, 137, 834–840. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Characteristics | Cases (n = 426) | Controls (n = 445) | p |
|---|---|---|---|
| Age, years | 0.283 | ||
| Mean ± SD | 57.4 ± 5.8 | 59.6 ± 4.7 | |
| Gender (%) | 0.447 | ||
| Male | 242 (56.8) | 269 (60.4) | |
| Female | 184 (43.2) | 176 (39.6) | |
| Races (Han, %) | 426 (100) | 445 (100) | 1 |
| Married (%) | 0.057 | ||
| Yes | 374 (87.8) | 401 (90.1) | |
| No | 52 (12.2) | 44 (9.9) | |
| BMI, kg/m2 (% a) | 0.2 | ||
| <25.0 | 94 (22.4) | 65 (14.8) | |
| ≥25.0 | 325 (77.6) | 375 (85.2) | |
| Missing b | 7 | 5 | |
| Family history of lung cancer (% a) | <0.001 | ||
| Yes | 74 (17.8) | 41 (9.5) | |
| No | 341 (82.2) | 390 (90.5) | |
| Missing b | 11 | 14 | |
| Education (% a) | |||
| Less than high school | 82 (19.5) | 65 (14.8) | |
| High school graduate | 339 (80.5) | 373 (85.2) | |
| Missing b | 5 | 7 | |
| Leisure physical activity (% a) | 0.252 | ||
| <3 times/week | 89 (21.2) | 111 (25.6) | |
| ≥3 times/week | 330 (78.8) | 322 (74.4) | |
| Missing b | 7 | 12 | |
| Dietary vitamin D intake (μg/day) | 5.7 ± 2.4 | 5.4 ± 3.1 | 0.759 |
| Smoking history (% a) | |||
| Yes | 297 (70.0) | 214 (48.5) | 0.001 |
| No | 127 (30.0) | 227 (51.5) | |
| Missing b | 2 | 4 | |
| Alcohol use (% a) | 0.132 | ||
| Yes | 261 (61.6) | 275 (63.2) | |
| No | 163 (38.4) | 160 (36.8) | |
| Missing b | 2 | 10 | |
| Plasma 25(OH)D (ng/mL) | 0.251 | ||
| Mean ± SD | 21.0 ± 6.3 | 22.5 ± 7.3 |
| Concentration of Plasma 25(OH)D a,b | ||||
|---|---|---|---|---|
| <20 ng/mL | ≥20 ng/mL | |||
| Smoking Status | Cases/Controls | OR c (95% CI) | Cases/Controls | OR b,c (95% CI) |
| Non-smoker | 85/149 | 1.00 (ref) | 42/78 | 1.00 (ref) |
| Smoker | 174/117 | 2.74 (1.97–3.01) | 123/97 | 2.48 (2.05–2.97) |
| p | <0.001 | <0.001 | ||
| Gene (Reference SNP number, rs) | Genotype | Cases (n = 426) | Controls (n = 445) | OR a (95% CI) | p |
|---|---|---|---|---|---|
| VDR (rs10735810) | CC | 166 (39.0%) | 160 (36.0%) | 1.00 (ref) | 0.47 |
| CT | 192 (45.1%) | 204 (45.8%) | 0.97 (0.73–1.41) | ||
| TT | 68 (15.9%) | 81 (18.2%) | 1.15 (0.84–1.37) | ||
| TT + CT | 260 (61.0%) | 285 (64.0%) | 1.09 (0.81–1.44) | ||
| VDR (rs11568820) | TT | 63 (14.8%) | 52 (11.7%) | 1.00 (ref) | 0.25 |
| TC | 324 (76.1%) | 360 (80.9%) | 1.22 (0.85–1.47) | ||
| CC | 39 (9.1%) | 33 (7.4%) | 0.96 (0.79–1.29) | ||
| CC + TC | 363 (85.2%) | 393 (88.3%) | 1.12 (0.87–1.54) | ||
| VDR (rs1544410) | GG | 403 (94.6%) | 373 (83.8%) | 1.00 (ref) | 0.032 |
| GA | 17 (4.0%) | 49 (11.0%) | 0.79 (0.64–1.13) | ||
| AA | 6 (1.4%) | 23 (5.2%) | 0.78 (0.68–0.96) | ||
| AA + GA | 23 (5.4%) | 72 (16.2%) | 0.78 (0.65–1.07) | ||
| VDR (rs7975232) | CC | 140 (32.9%) | 142 (31.9%) | 1.00 (ref) | 0.76 |
| CA | 191 (44.8%) | 214 (48.1%) | 1.11 (0.84–1.47) | ||
| AA | 95 (22.3%) | 89 (20.0%) | 0.99 (0.63–1.35) | ||
| AA + CA | 286 (67.1%) | 303 (68.1) | 1.02 (0.75–1.39) | ||
| VDR (rs731236) | TT | 409 (96.0%) | 416 (93.5%) | 1.00 (ref) | 0.037 |
| TC | 14 (3.3%) | 27 (6.1%) | 0.77 (0.59–0.99) | ||
| CC | 3 (0.7%) | 2 (0.4%) | 0.84 (0.57–0.95) | ||
| CC + TC | 17 (4.0%) | 29 (6.5%) | 0.79 (0.60–0.91) | ||
| CYP24A1 (rs6068816) | CC | 170 (39.9%) | 76 (17.1%) | 1.00 (ref) | <0.001 |
| CT | 222 (52.1%) | 307 (69.0%) | 0.85 (0.67–1.32) | ||
| TT | 34 (8.0%) | 62 (13.9%) | 0.31 (0.21–0.47) | ||
| TT + CT | 256 (60.1%) | 369 (82.9%) | 0.72 (0.58–1.02) | ||
| CYP24A1 (rs2244719) | TT | 312 (73.2%) | 320 (71.9%) | 1.00 (ref) | 0.65 |
| TC | 89 (20.9%) | 87 (19.6%) | 1.10 (0.92–1.35) | ||
| CC | 25 (5.9%) | 38 (8.5%) | 0.95 (0.77–1.16) | ||
| CC + TC | 114 (26.8%) | 125 (28.1%) | 1.04 (0.88–1.23) | ||
| CYP24A1 (rs4809960) | TT | 349 (81.9%) | 360 (80.9%) | 1.00 (ref) | 0.14 |
| TC | 62 (14.6%) | 75 (16.9%) | 0.85 (0.74–0.99) | ||
| CC | 15 (3.5%) | 10 (2.2%) | 1.09 (0.78–1.52) | ||
| CC + TC | 77 (18.1%) | 85 (19.1%) | 0.97 (0.75–1.21) | ||
| CYP24A1 (rs2762939) | GG | 160 (37.5%) | 156 (35.1%) | 1.00 (ref) | 0.50 |
| GC | 192 (45.1%) | 220 (49.4%) | 1.09 (0.85–1.32) | ||
| CC | 74 (17.4%) | 69 (15.5%) | 1.06 (0.79–1.33) | ||
| CC + GC | 266 (62.5%) | 289 (64.9%) | 1.08 (0.73–1.25) | ||
| CYP24A1 (rs2181874) | GG | 303 (71.1%) | 340 (76.4%) | 1.00 (ref) | 0.07 |
| GA | 84 (19.7%) | 76 (17.1%) | 1.17 (0.92–1.37) | ||
| AA | 39 (9.2%) | 29 (6.5%) | 1.40 (0.85–1.92) | ||
| AA + GA | 123 (28.9%) | 105 (23.6%) | 1.33 (0.89–1.65) | ||
| CYP24A1 (rs2296241) | GG | 119 (27.9%) | 114 (25.6%) | 1.00 (ref) | 0.09 |
| GA | 230 (54.0%) | 227 (51.0%) | 1.30 (0.88–1.63) | ||
| AA | 77 (18.1%) | 104 (23.4%) | 1.27 (0.76–1.55) | ||
| AA + GA | 307 (72.1%) | 331 (74.4%) | 1.29 (0.92–1.72) | ||
| CYP27B1 (rs10877012) | GG | 165 (38.7%) | 160 (35.9%) | 1.00 (ref) | 0.37 |
| GT | 209 (49.1%) | 209 (47.0%) | 1.10 (0.88–1.35) | ||
| TT | 52 (12.2%) | 76 (17.1%) | 0.95 (0.76–1.21) | ||
| TT + GT | 261 (61.3%) | 285 (64.1%) | 1.01 (0.75–1.32) | ||
| CYP27B1 (rs3782130) | CC | 194 (45.5%) | 187 (42.1%) | 1.00 (ref) | 0.15 |
| CG | 149 (35.0%) | 163 (36.6%) | 0.82 (0.76–1.45) | ||
| GG | 83 (19.5%) | 95 (21.3%) | 1.03 (0.89–1.34) | ||
| GG + CG | 232 (54.5%) | 258 (57.9%) | 0.94 (0.78–1.38) | ||
| GC (rs7041) | TT | 175 (41.1%) | 173 (38.8%) | 1.00 (ref) | <0.001 |
| TG | 230 (54.0%) | 225 (50.6%) | 0.79 (0.44–0.97) | ||
| GG | 21 (4.9%) | 47 (10.6%) | 0.61 (0.41–0.93) | ||
| GG + TG | 251 (58.9%) | 272 (61.2%) | 0.69 (0.38–1.15) | ||
| GC (rs4588) | CC | 230 (54.0%) | 235 (52.8%) | 1.00 (ref) | 0.58 |
| CA | 170 (39.9%) | 173 (38.9%) | 1.14 (0.91–1.45) | ||
| AA | 26 (6.1%) | 37 (8.3%) | 0.93 (0.71–1.43) | ||
| AA + CA | 196 (46.0%) | 210 (47.2%) | 1.11 (0.93–1.28) |
| Gene (rs) Case/Control | Homozygous Common Allele (Mean ± SD) | Heterozygous (Mean ± SD) | Homozygous Minor Allele (Mean ± SD) | Heterozygous + Homozygous Minor Allele (Mean ± SD) | p a | p b |
|---|---|---|---|---|---|---|
| VDR (rs10735810) | CC | CT | TT | TT + CT | ||
| Cases | 21.5 ± 6.7 | 22.6 ± 7.7 | 20.6 ± 8.5 | 22.0 ± 7.1 | 0.56 | 0.67 |
| Controls | 26.1 ± 5.4 | 25.3 ± 6.7 | 24.5 ± 8.2 | 24.9 ± 7.8 | 0.55 | 0.47 |
| VDR (rs11568820) | TT | TC | CC | CC + TC | ||
| Cases | 27.0 ± 6.4 | 24.9 ± 8.4 | 27.4 ± 7.4 | 25.9 ± 8.9 | 0.70 | 0.45 |
| Controls | 25.7 ± 9.2 | 27.4 ± 8.0 | 27.1 ± 10.3 | 27.3 ± 7.4 | 0.56 | 0.87 |
| VDR (rs1544410) | GG | GA | AA | AA + GA | ||
| Cases | 16.5 ± 8.6 | 20.7 ± 6.8 | 25.2 ± 7.3 | 24.6 ± 7.4 | 0.009 | 0.008 |
| Controls | 21.7 ± 4.6 | 23.0 ± 7.1 | 23.4 ± 7.0 | 23.1 ± 6.9 | 0.23 | 0.39 |
| VDR (rs7975232) | CC | CA | AA | AA + CA | ||
| Cases | 26.7 ± 6.7 | 25.9 ± 5.1 | 28.1 ± 7.7 | 26.9 ± 8.0 | 0.89 | 0.87 |
| Controls | 28.9 ± 5.7 | 25.8 ± 7.9 | 27.9 ± 7.3 | 27.1 ± 8.8 | 0.65 | 0.31 |
| VDR (rs731236) | TT | TC | CC | CC + TC | ||
| Cases | 24.5 ± 6.8 | 24.5 ± 6.9 | 23.4 ± 6.1 | 24.5 ± 5.4 | 0.70 | 0.87 |
| Controls | 27.7 ± 9.8 | 26.8 ± 5.1 | 28.6 ± 6.4 | 27.6 ± 7.5 | 0.80 | 0.65 |
| CYP24A1 (rs6068816) | CC | CT | TT | TT + CT | ||
| Cases | 16.4 ± 4.5 | 20.7 ± 7.3 | 25.7 ± 10.3 | 23.8 ± 7.7 | 0.007 | 0.01 |
| Controls | 17.9 ± 5.2 | 18.9 ± 7.8 | 18.4 ± 7.1 | 18.5 ± 7.2 | 0.20 | 0.27 |
| CYP24A1 (rs2244719) | TT | TC | CC | CC + TC | ||
| Cases | 28.6 ± 8.2 | 27.6 ± 7.8 | 29.0 ± 6.9 | 28.3 ± 6.2 | 0.80 | 0.85 |
| Controls | 26.8 ± 7.2 | 27.4 ± 6.5 | 25.5 ± 6.3 | 27.2 ± 6.3 | 0.51 | 0.44 |
| CYP24A1 (rs4809960) | TT | TC | CC | CC + TC | ||
| Cases | 24.6 ± 6.1 | 26.8 ± 8.2 | 25.7 ± 7.7 | 27.6 ± 7.4 | 0.76 | 0.60 |
| Controls | 25.9 ± 6.5 | 26.5 ± 7.6 | 27.7 ± 8.6 | 27.0 ± 8.0 | 0.62 | 0.56 |
| CYP24A1 (rs2762939) | GG | GC | CC | CC + GC | ||
| Cases | 25.9 ± 4.8 | 27.9 ± 7.1 | 27.1 ± 8.9 | 27.8 ± 7.0 | 0.61 | 0.70 |
| Controls | 31.4 ± 10.1 | 29.6 ± 8.9 | 29.7 ± 11.6 | 29.0 ± 10.5 | 0.90 | 0.93 |
| CYP24A1 (rs2181874) | GG | GA | AA | AA + GA | ||
| Cases | 26.2 ± 6.1 | 24.4 ± 5.7 | 24.4 ± 7.8 | 24.5 ± 7.6 | 0.29 | 0.15 |
| Controls | 28.1 ± 5.7 | 26.7 ± 8.9 | 24.9 ± 5.7 | 25.4 ± 8.1 | 0.13 | 0.09 |
| CYP24A1 (rs2296241) | GG | GA | AA | AA + GA | ||
| Cases | 24.6 ± 9.6 | 21.9 ± 9.5 | 20.5 ± 8.7 | 21.5 ± 8.5 | 0.13 | 0.21 |
| Controls | 25.7 ± 11.2 | 23.4 ± 10.1 | 24.0 ± 10.4 | 24.9 ± 9.5 | 0.36 | 0.61 |
| CYP27B1 (rs10877012) | GG | GT | TT | TT + GT | ||
| Cases | 28.1 ± 7.7 | 28.0 ± 8.3 | 29.4 ± 10.1 | 28.6 ± 9.8 | 0.80 | 0.71 |
| Controls | 27.1 ± 8.3 | 27.9 ± 4.9 | 29.9 ± 8.7 | 29.1 ± 5.6 | 0.33 | 0.34 |
| CYP27B1 (rs3782130) | CC | CG | GG | GG + CG | ||
| Cases | 27.0 ± 6.7 | 27.8 ± 8.9 | 25.7 ± 6.1 | 26.4 ± 7.4 | 0.81 | 0.77 |
| Controls | 26.5 ± 8.1 | 28.5 ± 14.3 | 28.2 ± 8.6 | 28.6 ± 10.7 | 0.25 | 0.10 |
| GC (rs7041) | TT | TG | GG | GG + TG | ||
| Cases | 24.7 ± 8.0 | 25.6 ± 6.9 | 26.1 ± 7.3 | 26.0 ± 6.3 | 0.37 | 0.43 |
| Controls | 28.0 ± 10.2 | 27.8 ± 7.1 | 29.5 ± 7.3 | 29.1 ± 8.4 | 0.87 | 0.61 |
| GC (rs4588) | CC | CA | AA | AA + CA | ||
| Cases | 24.7 ± 7.1 | 26.7 ± 7.9 | 27.1 ± 6.9 | 26.9 ± 7.1 | 0.73 | 0.74 |
| Controls | 27.2 ± 7.1 | 27.0 ± 7.5 | 29.1 ± 10.8 | 28.0 ± 6.5 | 0.19 | 0.41 |
| Homozygous Common Allele | Heterozygous and Homozygous Minor Allele | |||||
|---|---|---|---|---|---|---|
| Polymorphism | Plasma 25(OH)D (ng/mL) a | Cases/Controls | OR (95% CI) | Cases/Controls | OR (95% CI) | p-Interaction b |
| VDR (rs10735810) | <20 ng/mL | 47/38 | 1.00 (ref) | 78/56 | 1.00 (ref) | 0.015 |
| ≥20 ng/mL | 119/122 | 0.47 (0.24–1.43) | 182/229 | 0.46 (0.31–0.63) | ||
| VDR (rs11568820) | <20 ng/mL | 16/11 | 1.00 (ref) | 109/81 | 1.00(ref) | 0.263 |
| ≥20 ng/mL | 47/41 | 0.66 (0.34–1.52) | 254/312 | 0.68 (0.47–0.95) | ||
| VDR (rs1544410) | <20 ng/mL | 123/81 | 1.00 (ref) | 6/15 | 1.00 (ref) | 0.021 |
| ≥20 ng/mL | 280/292 | 0.73 (0.44–1.07) | 17/57 | 0.52 (0.29–0.97) | ||
| VDR (rs7975232) | <20 ng/mL | 42/29 | 1.00 (ref) | 84/64 | 1.00 (ref) | 0.631 |
| ≥20 ng/mL | 98/113 | 0.69 (0.41–1.21) | 202/239 | 0.74 (0.59–1.32) | ||
| VDR (rs731236) | <20 ng/mL | 124/83 | 1.00 (ref) | 5/6 | 1.00 (ref) | 0.841 |
| ≥20 ng/mL | 285/333 | 0.55 (0.27–0.66) | 12/23 | 0.60 (0.39–0.88) | ||
| CYP24A1 (rs6068816) | <20 ng/mL | 49/16 | 1.00 (ref) | 82/79 | 1.00 (ref) | 0.002 |
| ≥20 ng/mL | 121/60 | 0.41(0.28–0.61) | 174/290 | 0.48(0.25–0.65) | ||
| CYP24A1 (rs2244719) | <20 ng/mL | 87/66 | 1.00 (ref) | 35/26 | 1.00 (ref) | 0.731 |
| ≥20 ng/mL | 225/254 | 0.65 (0.42–0.91) | 79/99 | 0.78 (0.49–0.99) | ||
| CYP24A1 (rs4809960) | <20ng/mL | 100/71 | 1.00 (ref) | 23/18 | 1.00 (ref) | 0.724 |
| ≥20 ng/mL | 249/289 | 0.58 (0.31–1.05) | 54/67 | 0.67 (0.45–0.88) | ||
| CYP24A1 (rs2762939) | <20 ng/mL | 47/33 | 1.00 (ref) | 78/61 | 1.00 (ref) | 0.691 |
| ≥20 ng/mL | 113/123 | 0.67 (0.49–0.98) | 188/228 | 0.53 (0.31–0.87) | ||
| CYP24A1 (rs2181874) | <20 ng/mL | 98/77 | 1.00 (ref) | 46/21 | 1.00 (ref) | 0.113 |
| ≥20 ng/mL | 205/263 | 0.47 (0.28–0.90) | 77/84 | 0.72 (0.58–0.93) | ||
| CYP24A1 (rs2296241) | <20 ng/mL | 33/23 | 1.00 (ref) | 95/73 | 1.00 (ref) | 0.832 |
| ≥20 ng/mL | 86/91 | 0.79 (0.58–1.09) | 212/258 | 0.61 (0.40–0.97) | ||
| CYP27B1 (rs10877012) | <20 ng/mL | 48/32 | 1.00 (ref) | 78/64 | 1.00 (ref) | 0.875 |
| ≥20 ng/mL | 117/128 | 0.71 (0.53–0.98) | 183/221 | 0.74 (0.49–1.14) | ||
| CYP27B1 (rs3782130) | <20 ng/mL | 53/28 | 1.00 (ref) | 70/62 | 1.00 (ref) | 0.962 |
| ≥20 ng/mL | 141/159 | 0.80 (0.69–1.24) | 162/196 | 0.73 (0.53–1.00) | ||
| GC (rs7041) | <20 ng/mL | 43/36 | 1.00 (ref) | 73/59 | 1.00 (ref) | 0.182 |
| ≥20 ng/mL | 132/137 | 0.95 (0.81–1.53) | 178/213 | 0.57 (0.41–0.82) | ||
| GC (rs4588) | <20 ng/mL | 61/43 | 1.00 (ref) | 64/51 | 1.00 (ref) | 0.804 |
| ≥20 ng/mL | 169/192 | 0.52 (0.31–0.90) | 132/159 | 0.71 (0.39–0.86) | ||
| Gene (rs) Case/Control Smoking Status | Homozygous Common Allele (Mean ± SD) | Heterozygous (Mean ± SD) | Homozygous Minor Allele (Mean ± SD) | Heterozygous + Homozygous Minor Allele (Mean ± SD) | p a | p b |
|---|---|---|---|---|---|---|
| VDR (rs10735810) | CC | CT | TT | TT + CT | ||
| Cases | ||||||
| Smoker | 19.4 ± 7.8 | 16.9 ± 6.8 | 17.8 ± 5.9 | 17.5 ± 6.0 | 0.67 | 0.32 |
| Non-smoker | 24.8 ± 4.5 | 26.4 ± 10.6 | 21.8 ± 7.2 | 23.6 ± 8.4 | 0.45 | 0.87 |
| Controls | ||||||
| Smoker | 26.8 ± 3.5 | 24.7 ± 5.7 | 23.6 ± 5.4 | 24.1 ± 5.4 | 0.78 | 0.65 |
| Non-smoker | 25.8 ± 6.4 | 28.6 ± 9.3 | 24.6 ± 7.1 | 26.8 ± 8.2 | 0.41 | 0.44 |
| VDR (rs11568820) | TT | TC | CC | CC + TC | ||
| Cases | ||||||
| Smoker | 26.7 ± 7.8 | 25.8 ± 10.5 | 27.8 ± 10.2 | 26.6 ± 9.5 | 0.88 | 0.62 |
| Non-smoker | 27.8 ± 6.1 | 24.7 ± 7.9 | 26.3 ± 8.6 | 25.4 ± 7.2 | 0.75 | 0.37 |
| Controls | ||||||
| Smoker | 24.7 ± 6.7 | 25.8 ± 7.8 | 27.4 ± 8.8 | 26.4 ± 5.5 | 0.53 | 0.78 |
| Non-smoker | 26.7 ± 7.8 | 28.5 ± 9.2 | 26.6 ± 9.1 | 27.6 ± 7.9 | 0.84 | 0.93 |
| VDR (rs1544410) | GG | GA | AA | AA + GA | ||
| Cases | ||||||
| Smoker | 14.5 ± 7.8 | 18.5 ± 9.6 | 25.6 ± 7.0 | 24.1 ± 8.8 | 0.005 | 0.003 |
| Non-smoker | 18.9 ± 10.7 | 21.6 ± 4.7 | 25.1 ± 7.9 | 24.7 ± 5.6 | 0.01 | 0.01 |
| Controls | ||||||
| Smoker | 19.6 ± 12.5 | 22.4 ± 6.7 | 24.9 ± 7.8 | 23.8 ± 8.9 | 0.67 | 0.76 |
| Non-smoker | 22.3 ± 6.2 | 24.9 ± 7.5 | 25.4 ± 5.6 | 25.0 ± 7.5 | 0.08 | 0.12 |
| VDR (rs7975232) | CC | CA | AA | AA + CA | ||
| Cases | ||||||
| Smoker | 26.5 ± 12.2 | 25.7 ± 10.7 | 27.0 ± 5.6 | 26.3 ± 6.2 | 0.86 | 0.90 |
| Non-smoker | 28.9 ± 14.5 | 26.6 ± 6.2 | 28.4 ± 7.2 | 27.4 ± 7.3 | 0.91 | 0.76 |
| Controls | ||||||
| Smoker | 26.5 ± 8.4 | 24.6 ± 6.8 | 27.4 ± 7.9 | 26.4 ± 8.5 | 0.78 | 0.86 |
| Non-smoker | 30.4 ± 12.5 | 26.7 ± 10.6 | 28.4 ± 6.8 | 27.7 ± 9.3 | 0.56 | 0.12 |
| VDR (rs731236) | TT | TC | CC | CC + TC | ||
| Cases | ||||||
| Smoker | 21.5 ± 7.5 | 23.5 ± 3.2 | 22.5 ± 6.9 | 23.1 ± 4.7 | 0.88 | 0.91 |
| Non-smoker | 26.6 ± 4.4 | 25.1 ± 7.2 | 25.5 ± 5.6 | 25.4 ± 6.8 | 0.69 | 0.70 |
| Controls | ||||||
| Smoker | 29.0 ± 11.4 | 22.5 ± 5.6 | 29.3 ± 6.7 | 27.4 ± 10.2 | 0.87 | 0.75 |
| Non-smoker | 26.8 ± 9.7 | 28.3 ± 6.0 | 27.4 ± 7.1 | 27.9 ± 6.7 | 0.75 | 0.59 |
| CYP24A1 (rs6068816) | CC | CT | TT | TT + CT | ||
| Cases | ||||||
| Smoker | 14.7 ± 5.2 | 16.3 ± 7.8 | 18.7 ± 5.8 | 17.4 ± 6.8 | 0.002 | 0.02 |
| Non-smoker | 18.4 ± 4.7 | 22.4 ± 6.9 | 27.2 ± 13.4 | 25.6 ± 9.2 | 0.009 | 0.003 |
| Controls | ||||||
| Smoker | 15.6 ± 7.8 | 17.8 ± 10.3 | 16.5 ± 6.7 | 17.0 ± 7.8 | 0.18 | 0.12 |
| Non-smoker | 19.5 ± 6.7 | 18.6 ± 8.6 | 19.6 ± 6.9 | 19.2 ± 7.5 | 0.24 | 0.35 |
| CYP24A1 (rs2244719) | TT | TC | CC | CC + TC | ||
| Cases | ||||||
| Smoker | 26.4 ± 7.9 | 27.6 ± 10.9 | 27.7 ± 6.8 | 27.6 ± 5.7 | 0.95 | 0.85 |
| Non-smoker | 29.2 ± 11.3 | 28.5 ± 9.8 | 29.6 ± 8.5 | 29.0 ± 8.7 | 0.75 | 0.84 |
| Controls | ||||||
| Smoker | 27.0 ± 7.9 | 28.5 ± 5.7 | 24.6 ± 8.4 | 27.7 ± 6.8 | 0.54 | 0.74 |
| Non-smoker | 26.7 ± 9.1 | 25.7 ± 8.9 | 26.1 ± 6.7 | 26.0 ± 7.4 | 0.50 | 0.31 |
| CYP24A1 (rs4809960) | TT | TC | CC | CC + TC | ||
| Cases | ||||||
| Smoker | 23.7 ± 5.8 | 25.6 ± 9.2 | 24.7 ± 7.1 | 25.5 ± 6.7 | 0.78 | 0.67 |
| Non-smoker | 25.5 ± 5.7 | 28.3 ± 7.6 | 27.6 ± 8.7 | 28.0 ± 8.4 | 0.62 | 0.56 |
| Controls | ||||||
| Smoker | 26.8 ± 6.7 | 25.2 ± 6.5 | 29.7 ± 9.8 | 27.8 ± 6.5 | 0.87 | 0.67 |
| Non-smoker | 24.7 ± 7.8 | 27.8 ± 7.1 | 28.3 ± 10.3 | 28.1 ± 8.7 | 0.54 | 0.26 |
| CYP24A1 (rs2762939) | GG | GC | CC | CC + GC | ||
| Cases | ||||||
| Smoker | 25.6 ± 4.6 | 26.4 ± 6.8 | 26.6 ± 9.1 | 26.7 ± 7.6 | 0.71 | 0.65 |
| Non-smoker | 26.8 ± 5.3 | 28.5 ± 7.9 | 27.8 ± 7.6 | 28.1 ± 7.7 | 0.56 | 0.73 |
| Controls | ||||||
| Smoker | 30.3 ± 11.4 | 28.6 ± 9.5 | 29.8 ± 10.5 | 28.8 ± 9.6 | 0.91 | 0.97 |
| Non-smoker | 32.3 ± 14.5 | 30.4 ± 8.7 | 29.5 ± 12.1 | 29.9 ± 11.4 | 0.86 | 0.90 |
| CYP24A1 (rs2181874) | GG | GA | AA | AA + GA | ||
| Cases | ||||||
| Smoker | 25.7 ± 6.7 | 24.2 ± 9.2 | 23.5 ± 10.4 | 23.1 ± 8.6 | 0.26 | 0.17 |
| Non-smoker | 27.6 ± 4.6 | 26.7 ± 7.9 | 25.2 ± 6.4 | 25.8 ± 6.8 | 0.31 | 0.12 |
| Controls | ||||||
| Smoker | 28.0 ± 6.5 | 25.6 ± 7.8 | 24.6 ± 7.2 | 24.4 ± 6.5 | 0.06 | 0.07 |
| Non-smoker | 28.5 ± 7.9 | 27.8 ± 6.8 | 25.4 ± 7.0 | 26.4 ± 10.4 | 0.17 | 0.12 |
| CYP24A1 (rs2296241) | GG | GA | AA | AA + GA | ||
| Cases | ||||||
| Smoker | 24.3 ± 11.3 | 21.6 ± 10.5 | 20.6 ± 9.6 | 21.4 ± 7.5 | 0.18 | 0.25 |
| Non-smoker | 24.8 ± 7.4 | 22.7 ± 9.8 | 20.5 ± 8.5 | 21.8 ± 9.4 | 0.06 | 0.17 |
| Controls | ||||||
| Smoker | 24.4 ± 6.5 | 22.7 ± 8.3 | 23.5 ± 11.4 | 24.5 ± 10.7 | 0.44 | 0.77 |
| Non-smoker | 26.4 ± 14.7 | 25.7 ± 12.1 | 24.7 ± 10.9 | 25.1 ± 11.1 | 0.22 | 0.25 |
| CYP27B1 (rs10877012) | GG | GT | TT | TT + GT | ||
| Cases | ||||||
| Smoker | 26.1 ± 9.5 | 28.6 ± 7.8 | 30.1 ± 10.3 | 28.4 ± 11.4 | 0.87 | 0.65 |
| Non-smoker | 28.8 ± 6.4 | 27.5 ± 8.9 | 29.7 ± 9.4 | 28.6 ± 9.1 | 0.76 | 0.86 |
| Controls | ||||||
| Smoker | 27.0 ± 7.2 | 25.7 ± 8.9 | 29.7 ± 6.7 | 28.8 ± 8.1 | 0.21 | 0.45 |
| Non-smoker | 27.8 ± 9.8 | 29.9 ± 7.4 | 30.4 ± 7.9 | 30.0 ± 7.7 | 0.43 | 0.28 |
| CYP27B1 (rs3782130) | CC | CG | GG | GG + CG | ||
| Cases | ||||||
| Smoker | 24.5 ± 5.6 | 26.5 ± 7.5 | 26.0 ± 5.8 | 26.3 ± 6.1 | 0.87 | 0.56 |
| Non-smoker | 27.5 ± 8.5 | 29.8 ± 10.6 | 28.6 ± 7.0 | 28.1 ± 9.2 | 0.73 | 0.96 |
| Controls | ||||||
| Smoker | 25.4 ± 5.1 | 30.2 ± 11.5 | 27.5 ± 9.8 | 28.9 ± 10.8 | 0.13 | 0.09 |
| Non-smoker | 27.6 ± 9.6 | 27.7 ± 10.6 | 28.8 ± 7.9 | 28.1 ± 10.7 | 0.42 | 0.12 |
| GC (rs7041) | TT | TG | GG | GG + TG | ||
| Cases | ||||||
| Smoker | 23.9 ± 7.8 | 26.5 ± 6.7 | 25.3 ± 6.5 | 26.1 ± 5.9 | 0.45 | 0.34 |
| Non-smoker | 25.1 ± 8.3 | 24.7 ± 7.8 | 26.2 ± 5.8 | 25.7 ± 7.1 | 0.32 | 0.56 |
| Controls | ||||||
| Smoker | 28.0 ± 11.8 | 28.9 ± 6.8 | 29.8 ± 7.9 | 29.3 ± 7.4 | 0.91 | 0.56 |
| Non-smoker | 28.1 ± 9.5 | 28.6 ± 9.4 | 29.6 ± 8.1 | 29.0 ± 9.2 | 0.83 | 0.67 |
| GC (rs4588) | CC | CA | AA | AA + CA | ||
| Cases | ||||||
| Smoker | 24.3 ± 6.1 | 26.3 ± 5.9 | 27.0 ± 8.3 | 26.7 ± 7.4 | 0.43 | 0.61 |
| Non-smoker | 25.1 ± 6.0 | 27.1 ± 9.4 | 27.4 ± 5.7 | 27.3 ± 8.7 | 0.85 | 0.93 |
| Controls | ||||||
| Smoker | 25.7 ± 3.5 | 26.4 ± 5.6 | 27.4 ± 7.6 | 27.0 ± 6.3 | 0.21 | 0.34 |
| Non-smoker | 29.6 ± 8.2 | 27.3 ± 8.9 | 29.6 ± 11.5 | 28.5 ± 10.2 | 0.16 | 0.56 |
| Genotype | Non-Smoker | Smoker | ||||
|---|---|---|---|---|---|---|
| Cases (%) | Controls (%) | OR a (95% CI) | Cases (%) | Controls (%) | OR a (95% CI) | |
| CYP24A1 (rs2181874) | ||||||
| GG + GA | 102 (80.3%) | 215 (94.7%) | 1.00 (ref) | 256 (86.2%) | 197 (92.1%) | 1.00 (ref) |
| AA | 25 (19.7%) | 12 (5.3%) | 2.14 (1.47–3.43) | 41 (13.8%) | 17 (7.9%) | 3.57 (2.66–4.74) |
| p | 0.031 | 0.019 | ||||
| p-Interaction | 0.016 | |||||
| CYP24A1 (rs6068816) | ||||||
| CC + CT | 91 (71.7%) | 158 (74.0%) | 1.00 (ref) | 282 (94.9%) | 182 (85.1%) | 1.00 (ref) |
| TT | 36 (28.3%) | 69 (30.4%) | 0.84 (0.65–1.41) | 15 (5.1%) | 32 (14.9%) | 0.43 (0.27–1.02) |
| p | 0.079 | 0.006 | ||||
| p-Interaction | 0.038 | |||||
| VDR (rs10735810) | ||||||
| CC + CT | 112 (88.2%) | 198 (87.2%) | 1.00 (ref) | 237 (79.8%) | 186 (86.9%) | 1.00 (ref) |
| TT | 15 (11.8%) | 29 (12.8%) | 1.16 (0.82–1.34) | 60 (20.2%) | 28 (13.1%) | 1.93 (1.41–2.76) |
| p | 0.256 | 0.015 | ||||
| p-Interaction | 0.004 | |||||
| VDR (rs1544410) | ||||||
| GG + GA | 116 (91.3%) | 172 (75.8%) | 1.00 (ref) | 294 (98.9%) | 175 (81.8%) | 1.00 (ref) |
| AA | 11 (8.7%) | 55 (24.2%) | 0.51 (0.34–1.17) | 3 (1.0%) | 39 (18.2%) | 0.26 (0.20–0.69) |
| p | 0.002 | 0.001 | ||||
| p-Interaction | 0.002 | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Cheng, J.; Yang, K. Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk. Int. J. Mol. Sci. 2016, 17, 1597. https://doi.org/10.3390/ijms17101597
Wu X, Cheng J, Yang K. Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk. International Journal of Molecular Sciences. 2016; 17(10):1597. https://doi.org/10.3390/ijms17101597
Chicago/Turabian StyleWu, Xiayu, Jiaoni Cheng, and Kaiyun Yang. 2016. "Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk" International Journal of Molecular Sciences 17, no. 10: 1597. https://doi.org/10.3390/ijms17101597
APA StyleWu, X., Cheng, J., & Yang, K. (2016). Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk. International Journal of Molecular Sciences, 17(10), 1597. https://doi.org/10.3390/ijms17101597