
Morphological, Genome and Gene Expression Changes in Newly Induced Autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino





Abstract
:1. Introduction
2. Results
2.1. Chromosome Number and Morphology Analysis
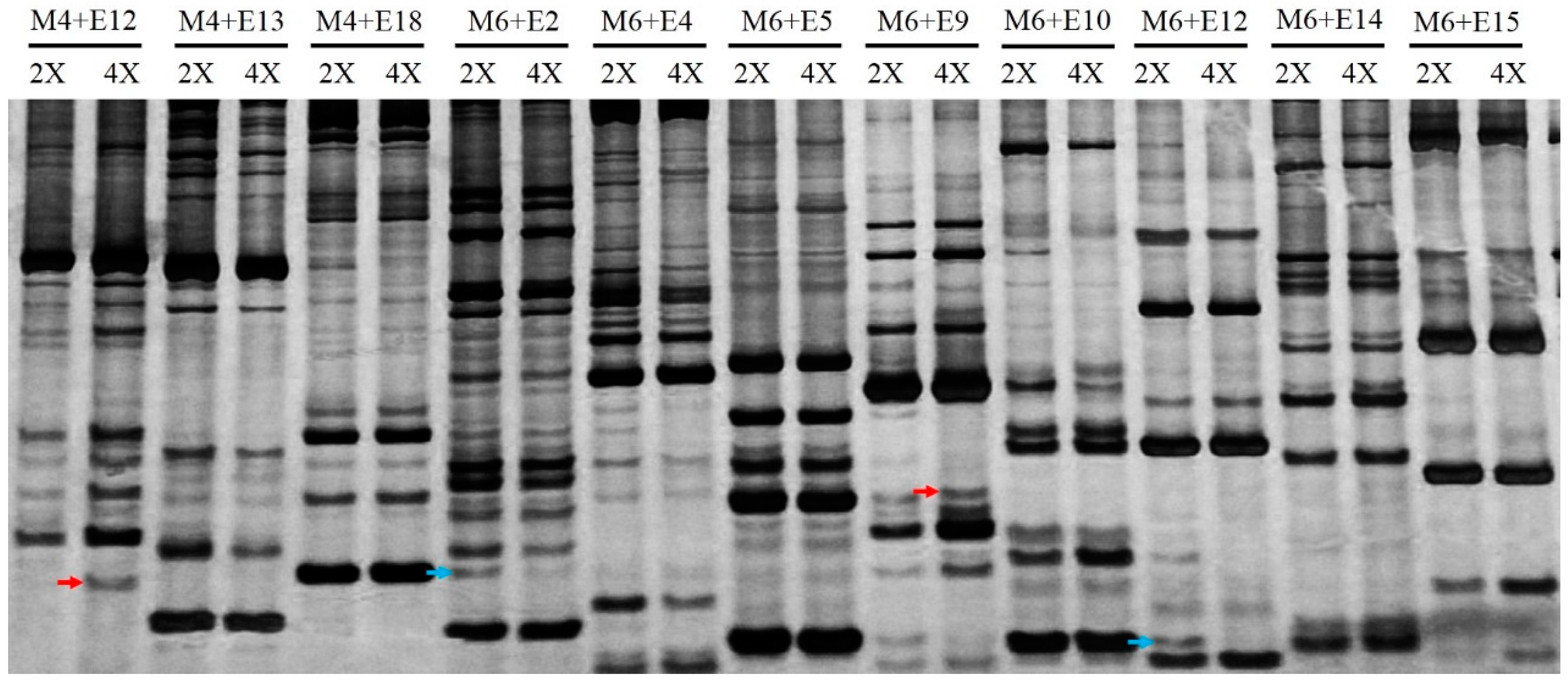
2.2. Genomic Variation Induced by Autopolyploidy
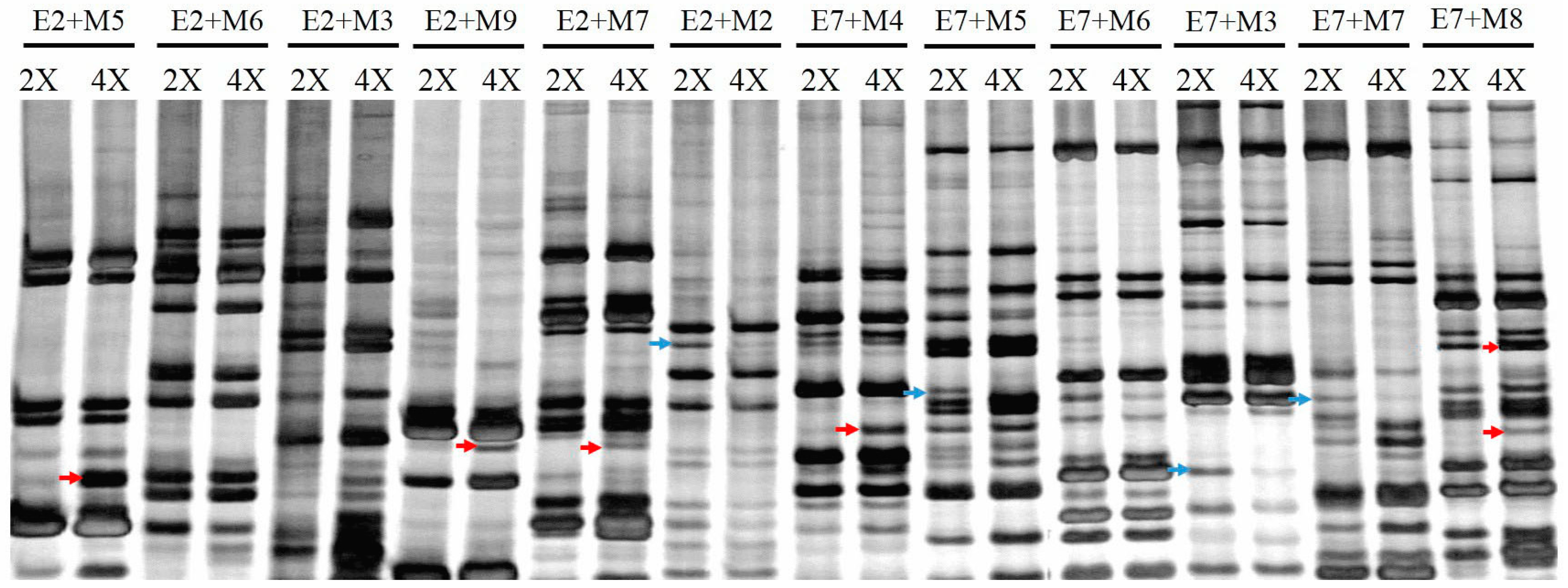
2.3. Detection of Different Transcripts
2.4. Epigenetic Changes Induced by Autopolyploidy
3. Discussion
3.1. Chromosome Number and Morphology Analysis
3.2. Implications of Rapid Genome Change for Polyploidy Evolution
3.3. Induced Differences in the Transcriptome
3.4 Methylation Alterations in Synthesized Autopolyploids
4. Materials and Method
4.1. Plant Material and Growth Conditions
4.2. Morphological and Anatomy Analysis
4.3. Genomic Variation Induced by Autopolyploidy
4.4. Complementary DNA Amplified Fragment Length Polymorphism Analysis
4.5. Mthylation Sensitive Amplified Polymorphism Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Song, K.; Lu, P.; Tang, K.; Osborn, T. Rapid genome change in synthetic polyploids of Brassica and its implications for polyploid evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 7719–7723. [Google Scholar] [CrossRef] [PubMed]
- Parisod, C.; Holderegger, R.; Brochmann, C. Evolutionary consequences of autopolyploidy. New Phytol. 2010, 186, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.Y.; Dilkes, B.; Luo, H.; Douglas, A.; Yakubova, E.; Lahner, B.; Salt, D.E. Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis. Science 2013, 341, 658–659. [Google Scholar] [CrossRef] [PubMed]
- Gornickia, P.; Farisb, J.D. Rewiring the wheat reproductive system to harness heterosis for the next wave of yield improvement. Proc. Natl. Acad. Sci. USA 2014, 111, 9024–9025. [Google Scholar] [CrossRef] [PubMed]
- Lavania, U.C.; Srivastava, S.; Lavania, S.; Basu, S.; Misra, N.K.; Muka, Y. Autopolyploidy differentially influences body size in plants, but facilitates enhanced accumulation of secondary metabo-lites, causing increased cytosine methylation. Plant J. 2012, 71, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, D.; Chen, Z.J.; Pires, J.C. Epigenetic regulation of flowering time in polyploids. Curr. Opin. Plant Biol. 2011, 14, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Soltis, P.S. Polyploidy: Recurrent formation and genome evolution. Trends Ecol. Evol. 1999, 14, 348–352. [Google Scholar] [CrossRef]
- Wendel, J.F. Genome evolution in polyploids. Plant Mol. Biol. 2000, 42, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Osborn, T.C.; Pires, J.C.; Birchler, J.A.; Auger, D.L. Understanding mechanisms of novel gene expression in polyploids. Trends Genet. 2003, 19, 14–147. [Google Scholar] [CrossRef]
- Coate, J.E.; Bar, H.; Doylec, J.J. Extensive translational regulation of gene expression in an allopolyploid (Glycine dolichocarpa). Plant Cell 2014, 26, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Shaked, H.; Kashkush, K.; Ozkan, H.; Feldman, M.; Levy, A. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat. Plant Cell 2001, 13, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, J.; Chen, S.; Fang, W.; Guan, Z.; Liao, Y.; Chen, F. Rapid genomic and transcriptomic alterations induced by wide hybridization: Chrysanthemum nankingense × Tanacetum vulgare and C. crassum × Crossostephium chinense (Asteraceae). BMC Genom. 2013, 14, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, J.; Chen, S.; Qi, X.; Fang, W.; Guan, Z.; Teng, N.; Liao, Y.; Chen, F. Rapid genetic and epigenetic alterations under intergeneric genomic shock in newly synthesized Chrysanthemum morifolium × Leucanthemum paludosum hybrids (Asteraceae). Genome Biol. Evol. 2014, 6, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971. [Google Scholar]
- Ramsey, J.; Schemske, D. Neopolyploidy in flowering plants. Annu. Rev. Ecol. Syst. 2002, 33, 589–639. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J. Polyploidy and ecological adaptation in wild yarrow. Proc. Natl. Acad. Sci. USA 2011, 108, 7096–7101. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wendel, J. Epigenetic phenomena and the evolution of plant allopolyploids. Mol. Phylogenet. Evol. 2003, 29, 365–379. [Google Scholar] [CrossRef]
- Rapp, R.; Wendel, J. Epigenetics and plant evolution. New Phytol. 2005, 168, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Shin, C. MicroRNA-target interactions: New insights from genome wide approaches. Ann. N. Y. Acad. Sci. 2012, 1271, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, R.; Lindquist, S. Epigenetics in the extreme: Prions and the inheritance of environmentally acquired traits. Science 2010, 330, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Vanyushin, B.; Ashapkin, V. DNA methylation in higher plants: Past, present and future. BBA Gene Regul. Mech. 2011, 1809, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Dar, T.H.; Raina, S.N.; Goel, S. Molecular analysis of genomic changes in synthetic autotetraploid Phlox drummondii Hook. Biol. J. Linn. Soc. 2013, 110, 591–605. [Google Scholar] [CrossRef]
- Rodriguez, M.; Cervigni, G.; Quarin, C.; Ortiz, J. Frequencies and variation in cytosine methylation patterns in diploid and tetraploid cytotypes of Paspalum notatum. Biol. Plant. 2012, 56, 276–282. [Google Scholar] [CrossRef]
- Kashkush, K.; Feldman, M.; Levy, A.A. Gene loss, silencing and activation in a newly synthesized wheat allotetraploid. Genetics 2002, 160, 1651–1659. [Google Scholar] [PubMed]
- Church, S.A.; Spaulding, E.J. Gene expression in a wild autopolyploid sunflower series. J. Hered. 2009, 100, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Pan, X.; Zhang, L.; Huang, B.; Sun, L.; Li, B.; Yi, B.; Zheng, S.; Yu, X.; Chen, W. A genome-wide comparison of genes responsive to autopolyploidy in Isatis indigotica using Arabidopsis thaliana Affymetrix Genechips. Plant Mol. Biol. Rep. 2006, 24, 197–204. [Google Scholar] [CrossRef]
- Pignatta, D.; Dilkes, B.P.; Yoo, S.Y.; Henry, I.M.; Madlung, A.; Doerge, R.W.; Chen, Z.J.; Comai, L. Differential sensitivity of the Arabidopsis thaliana transcriptome and enhancers to the effects of genome doubling. New Phytol. 2010, 186, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Karis, P.O. Morphological phylogenetics of the Asteraceae-Asteroideae, with notes on character evolution. Plant Syst. Evol. 1993, 186, 69–93. [Google Scholar] [CrossRef]
- Allum, J.F.; Bringloe, D.H.; Roberts, A.V. Chromosome doubling in a Rosa rugosa Thunb. hybrid by exposure of in vitro nodes to oryzalin: The effects of node length, oryzalin concentration and exposure time. Plant Cell Rep. 2007, 26, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Petolino, J.; Widholm, J. Efficient production of doubled haploid plants through colchicine treatment of anther-derived maize callus. Theor. Appl. Genet. 1989, 77, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, Z.; Bao, M. Colchicine-induced chromosome doubling in Platanus acerifolia and its evect on plant morphology. Euphytica 2007, 157, 145–154. [Google Scholar] [CrossRef]
- El-Morsy, S.I.; Dorra, M.; El-Hady, E.A.A.; Hiaba, A.A.; Mohamed, A. Comparative studies on diploid and tetraploid levels of Nicotiana alata. Acad. J. Plant Sci. 2009, 2, 182–188. [Google Scholar]
- Rauf, S.; Munir, H.; Abdullojon, E.; Basra, S.M. Role of colchicine and plant growth regulators to overcome interspecific incompatibility. Gen. Appl. Plant Physiol. 2006, 32, 223–232. [Google Scholar]
- Tsukaya, H. Controlling size in multicellular organs: Focus on the leaf. PLoS Biol. 2008, 6, 1373–1376. [Google Scholar] [CrossRef] [PubMed]
- Leiva-Neto, J.; Grafi, G.; Sabelli, P.; Dante, R.; Woo, Y.-M. A dominant negative mutant of Cyclin-Dependent Kinase A reduces endoreduplication but not cell size or gene expression in maize endosperm. Plant Cell 2004, 16, 1854–1869. [Google Scholar] [CrossRef] [PubMed]
- Španiel, S.; Marhold, K.; Hodálová, I.; Lihová, J. Diploid and tetraploid cytotypes of Centaurea stoebe (Asteraceae) in central europe: Morphological differentiation and cytotype Distribution Patterns. Folia Geobot. 2008, 43, 131–158. [Google Scholar]
- Werner, J.E.; Peloqu, S.J. Occurrence and mechanisms of 2n egg formation in 2x potato. Genome 1991, 34, 975–982. [Google Scholar] [CrossRef]
- Bretagnolle, B.F.; Thompson, J.D. Gametes with the somatic chromosome number: Mechanisms of their formation and role in the evolution of autopolyploid plants. New Phytol. 1995, 129, 1–22. [Google Scholar] [CrossRef]
- Soltis, D.; Soltis, P.; Schemske, D.; Hancock, J.; Thompson, J.; Husband, B.; Judd, W.S. Autopolyploidy in angiosperms: Have we grossly underestimated the number of species? Taxon 2007, 56, 13–30. [Google Scholar]
- Ozkan, H.; Levy, A.A.; Feldman, M. Allopolyploidy-Induced rapid genome evolution in the wheat group. Plant Cell 2001, 13, 1735–1747. [Google Scholar] [CrossRef] [PubMed]
- Mecchia, M.; Ochogaví, A.; Selva, J.; Laspina, N.; Felitti, S. Genome polymorphisms and gene differential expression in a ‘back-and-forth’ ploidy-altered series of weeping lovegrass. J. Plant Physiol. 2007, 164, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Martelotto, L.G.; Ortiz, J.P.A.; Stein, J.; Espinoz, F.; Quarin, C.L.; Pessino, S.C. Genome rearrangements derived from autopolyploidization in Paspalum sp. Plant Sci. 2007, 172, 970–977. [Google Scholar] [CrossRef]
- Bardil, A.; Tayal, A.; Parisod, C. Evolutionary dynamics of retrotransposons following autopolyploidy in the Buckler Mustard species complex. Plant J. 2015, 82, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Flagel, L.E.; Wendel, J.F.; Udall, J.A. Duplicate gene evolution, homoeologous recombination, and transcriptome characterization in allopolyploid cotton. BMC Genom. 2012, 13, 302–115. [Google Scholar] [CrossRef] [PubMed]
- Akhunova, A.R.; Matniyazov, R.T.; Liang, H.; Akhunov, E.D. Homoeolog-specific transcriptional bias in allopolyploid wheat. BMC Genom. 2010, 11, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Madlung, A.; Tyagi, A.P.; Watson, B.; Jiang, H.; Kagochi, T.; Doerge, R.W.; Martienssen, R.; Comai, L. Genomic changes in synthetic Arabidopsis polyploids. Plant J. 2005, 41, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, L.; Madlung, A.; Lee, H.-S.; Chen, M.; Lee, J.J.; Watson, B.; Kagochi, T.; Comai, L.; Chen, Z.J. Stochastic and epigenetic changes of gene expression in Arabidopsis polyploids. Genetics 2004, 167, 1961–1973. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Chen, Z.J. Protein-coding genes are epigenetically regulated in Arabidopsis polyploids. Proc. Natl. Acad. Sci. USA 2001, 98, 6753–6758. [Google Scholar] [CrossRef] [PubMed]
- Richards, E.; Elgin, S. Epigenetic codes for heterochromatin formation and silencing: Rounding up the usual suspects. Cell 2002, 108, 489–500. [Google Scholar] [CrossRef]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Anssour, S.; Krügel, T.; Sharbel, T.F.; Saluz, H.P.; Bonaventure, G.; Baldwin, I.T. Phenotypic, genetic and genomic consequences of natural and synthetic polyploidization of Nicotiana attenuata and Nicotiana obtusifolia. Ann. Bot. 2009, 103, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Cervigni, G.; Paniego, N.; Pessino, S.; Selva, J.; Diaz, M.; Spangenberg, G.; Echenique, V. Gene expression in diplosporous and sexual Eragrostis curvula genotypes with differing ploidy levels. Plant Mol. Biol. 2008, 67, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Deng, M.; Fan, G. Differential transcriptome analysis between paulownia fortunei and its synthesized autopolyploid. Int. J. Mol. Sci. 2014, 15, 5079–5093. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Lua, J.; Tian, L.; Ramachandrand, V.; Kasschau, K.D.; Chapman, E.J.; Carrington, J.C.; Chen, X.; Wang, X.-J.; Chen, Z.J. Small RNAs serve as a genetic buffer against genomic shock in Arabidopsis interspecific hybrids and allopolyploids. Proc. Natl. Acad. Sci. USA 2009, 106, 17835–17840. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Ni, Z. Mechanisms of genomic rearrangements and gene expression changes in plant polyploids. Bioessay 2006, 28, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.F.; Cosson, P.; Guirao, A.; Alary, R.; Joudrier, P. Puroindoline genes are highly conserved in diploid ancestor wheats and related species but absent in tetraploid Triticum species. Plant Sci. 2000, 153, 81–91. [Google Scholar] [CrossRef]
- Feuillet, C.; Penger, A.; Gellner, K.; Mast, A.; Keller, B. Molecular evolution of receptor-like kinase genes in hexaploid wheat. Independent evolution of orthologs after polyploidization and mechanisms of local rearrangements at paralogous loci. Plant Physiol. 2001, 125, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gu, Y.; Chen, C.; Jiao, D.; Xue, Z.; Song, W. Analysis of the level and pattern of genomic DNA methylation in different ploidy watermelons by MSAP (Citrullus lanatus). J. Mol. Cell Biol. 2009, 42, 118–125. [Google Scholar]
- Aversano, R.; Caruso, I.; Aronne, G.; Micco, V.D.; Scognamiglio, N. Stochastic changes affect Solanum wild species following autopolyploidization. J. Exp. Bot. 2013, 62, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wu, R.; Zhang, Y.F.; Liu, X.M.; Wang, H.Y.; Gong, L.; Zhang, Z.H.; Liu, B. Tissue culture-induced locus-specific alteration in DNA methylation and its correlation with genetic variation in Codonopsis lanceolata Benth. et Hook. Plant Cell Rep. 2007, 26, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Matthes, M.; Singh, R.; Cheah, S.-C.; Karp, A. Variation in oil palm (Elaeis guineensis Jacq.) tissue culture-derived regenerants revealed by AFLPs with methylation-sensitive enzymes. Theor. Appl. Genet. 2001, 102, 971–979. [Google Scholar] [CrossRef]
- Stroud, H.; Ding, B.; Simon, S.A.; Feng, S.; Bellizzi, M.; Pellegrini, M.; Wang, G.L.; Meyers, B.C.; Jacobsen, S.E. Plants regenerated from tissue culture contain stable epigenome changes in rice. Elife 2013, 2, e00354. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Lee, T.V.D.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acid Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Diploid | Tetraploid |
|---|---|---|
| Plant height (cm) | 83.2 ± 2.2 | 85.13 ± 1.92 |
| Leaf length (cm) | 4.19 ± 0.20 | 4.73 ± 0.21 * |
| Leaf width (cm) | 2.74 ± 0.13 | 3.50 ± 0.21 * |
| Inflorescence diameter (mm) | 16.8 ± 0.24 | 16.65 ± 0.53 |
| Disc flower diameter (mm) | 7.73 ± 0.36 | 8.41 ± 0.25 |
| No. of ligulate florets | 13.11 ± 0.48 * | 10.61 ± 0.86 |
| Ligulate florets dry weight (mg/100) | 0.018 ± 0.0015 | 0.03 ± 0.0012 ** |
| No. of tubular florets | 78.75 ± 1.97 ** | 57.71 ± 1.69 |
| Tubular florets dry weight (mg/100) | 0.015 ± 0.0014 | 0.033 ± 0.0010 ** |
| Palisade thickness (µm) | 43.39 ± 0.33 | 47.83 ± 1.85 * |
| Spongy tissue thickness (µm) | 76.67 ± 1.01 | 85.16 ± 1.41 * |
| Upper epidermis thickness (µm) | 8.51 ± 0.18 | 8.99 ± 0.32 |
| Lower epidermis thickness (µm) | 7.40 ± 0.17 | 6.86 ± 0.17 |
| Leaf thickness (µm) | 136.45 ± 1.16 | 148.84 ± 0.92 * |
| No. Cell (104) | 138.99 ± 22.49 | 161.89 ± 2.91 * |
| Size of cell | 15.17 ± 0.36 | 23.86 ± 0.57 * |
| Plant Lines | Total Sites | Nonmethylated Type I | Methylated | ||
|---|---|---|---|---|---|
| Type II | Type III | Total (II + III) | |||
| Diploid | 260 | 61 (23.5%) | 114 (43.8%) | 85 (32.7%) | 199 (76.5%) |
| Tetraploid | 251 | 52 (20.7%) | 109 (43.4%) | 90 (35.9%) | 199 (79.3%) |
| Fragment Type | Fragment Display Pattern in MSAP Gel | Number of Sites | Sataus | ||||
|---|---|---|---|---|---|---|---|
| 2X | 4X | 2X H lane | 2X M lane | 4X H lane | 4X M lane | 2 | ↓ |
| Type IV | Type I | − | − | + | + | 0 | ↓ |
| Type I | Type IV | + | + | − | − | 2 | ↑ |
| Type IV | Type III | − | − | + | − | 7 | ↓ |
| Type III | Type IV | + | − | − | − | 12 | ↑ |
| Type IV | Type II | − | − | − | + | 4 | ↓ |
| Type II | Type IV | − | + | − | − | 5 | ↑ |
| Type II | Type I | − | + | + | + | 4 | ↓ |
| Type I | Type II | + | + | − | + | 5 | ↑ |
| Type III | Type I | + | − | + | + | 1 | ↓ |
| Type I | Type III | + | + | + | − | 13 | ↑ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, R.; Wang, H.; Dong, B.; Yang, X.; Chen, S.; Jiang, J.; Zhang, Z.; Liu, C.; Zhao, N.; Chen, F. Morphological, Genome and Gene Expression Changes in Newly Induced Autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino. Int. J. Mol. Sci. 2016, 17, 1690. https://doi.org/10.3390/ijms17101690
Gao R, Wang H, Dong B, Yang X, Chen S, Jiang J, Zhang Z, Liu C, Zhao N, Chen F. Morphological, Genome and Gene Expression Changes in Newly Induced Autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino. International Journal of Molecular Sciences. 2016; 17(10):1690. https://doi.org/10.3390/ijms17101690
Chicago/Turabian StyleGao, Ri, Haibin Wang, Bin Dong, Xiaodong Yang, Sumei Chen, Jiafu Jiang, Zhaohe Zhang, Chen Liu, Nan Zhao, and Fadi Chen. 2016. "Morphological, Genome and Gene Expression Changes in Newly Induced Autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino" International Journal of Molecular Sciences 17, no. 10: 1690. https://doi.org/10.3390/ijms17101690
APA StyleGao, R., Wang, H., Dong, B., Yang, X., Chen, S., Jiang, J., Zhang, Z., Liu, C., Zhao, N., & Chen, F. (2016). Morphological, Genome and Gene Expression Changes in Newly Induced Autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino. International Journal of Molecular Sciences, 17(10), 1690. https://doi.org/10.3390/ijms17101690