
HMGB1 Promotes Intraoral Palatal Wound Healing through RAGE-Dependent Mechanisms


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
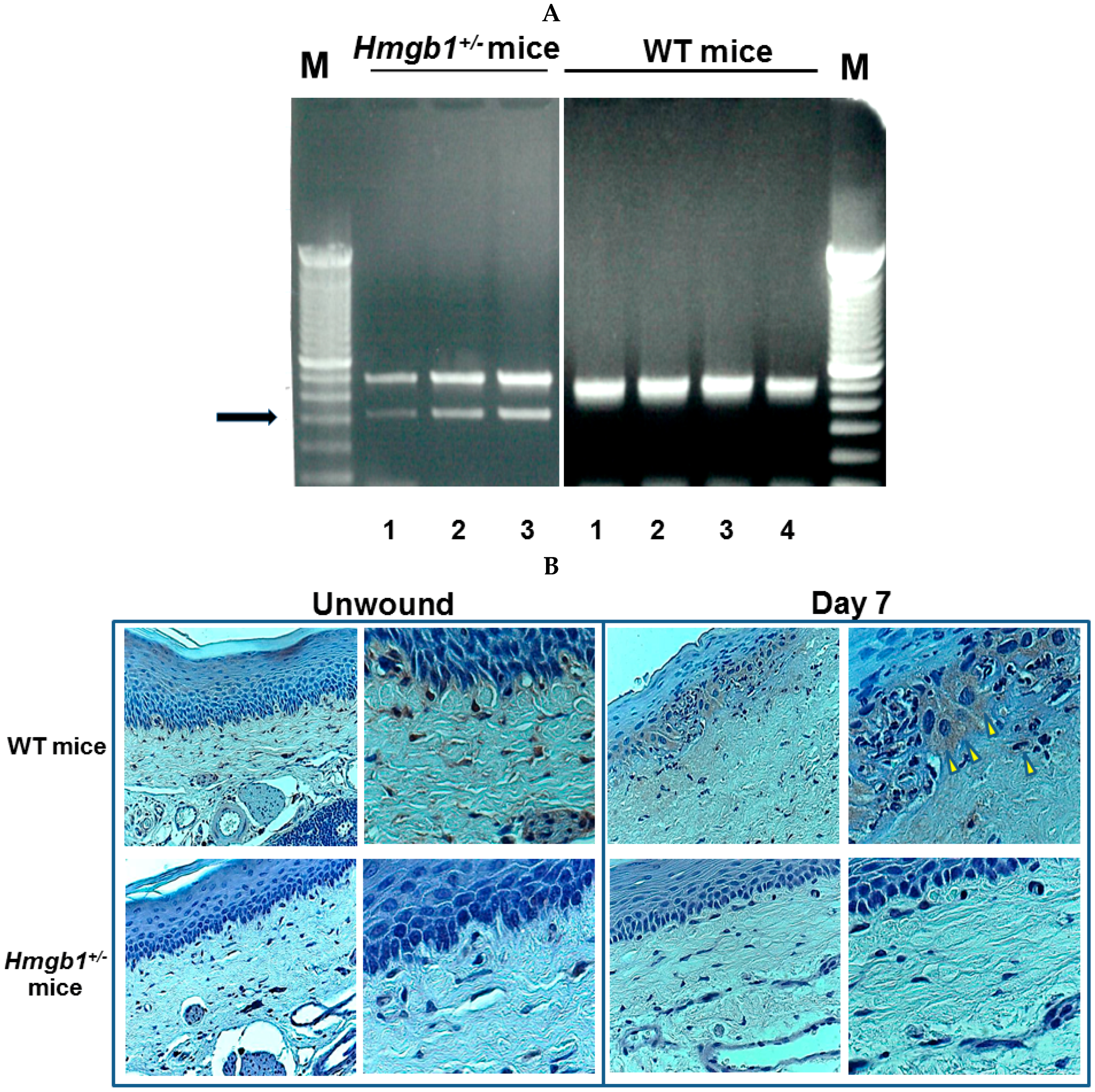
2.1. Identification and Targeted Disruption of the Mouse Gene
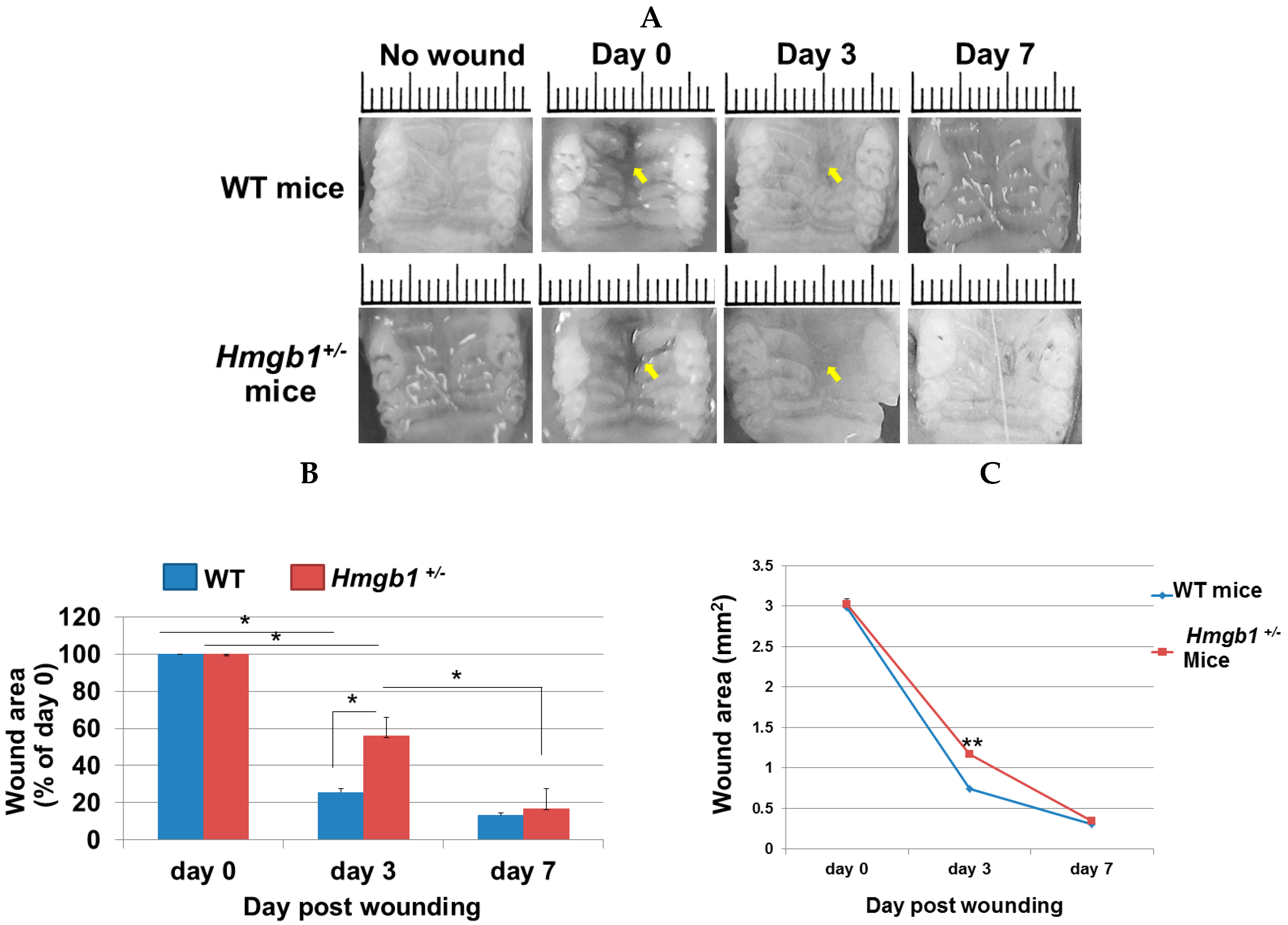
2.2. Wound Closure Is Attenuated in Hmgb1+/− Mice
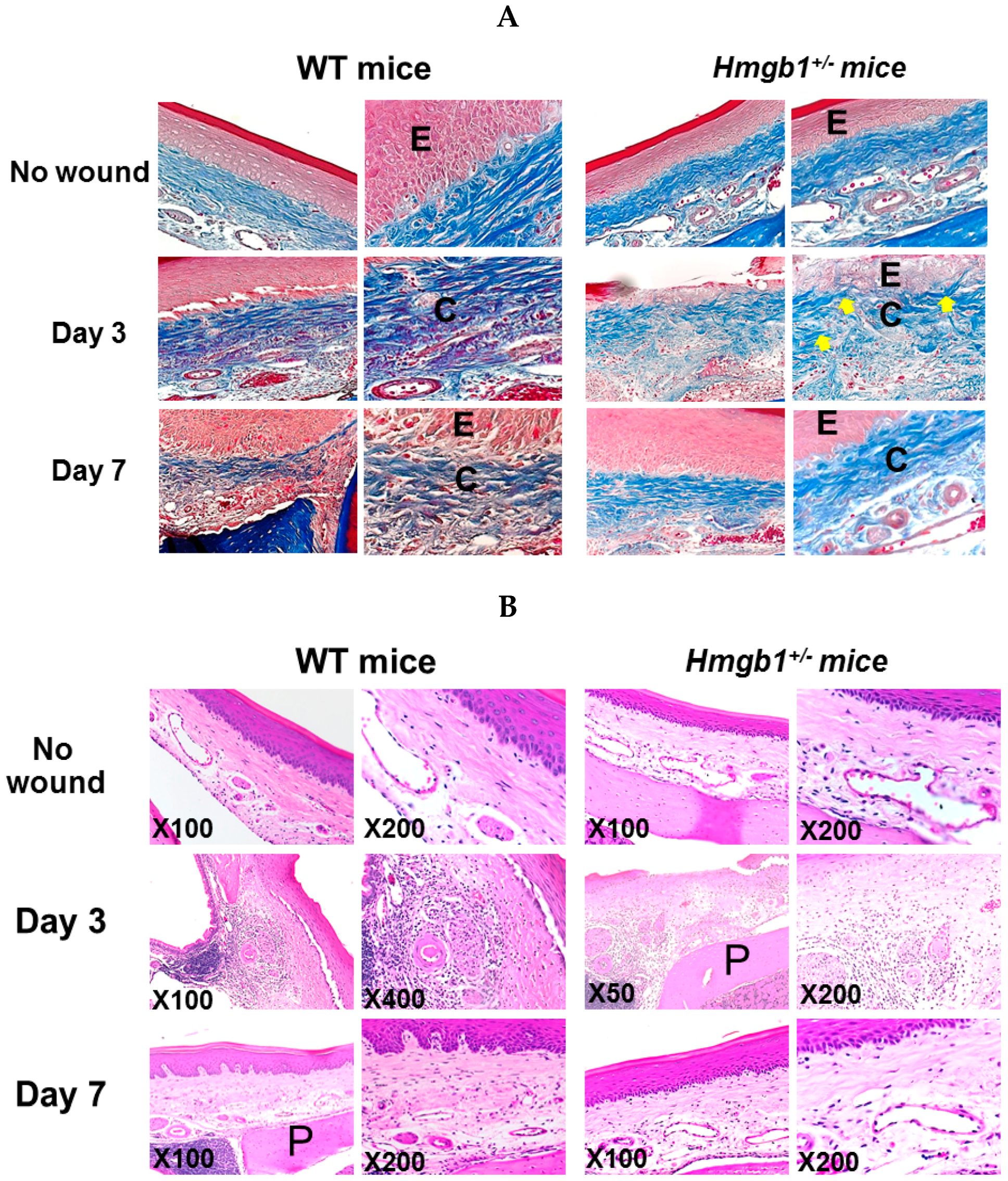
2.3. Reduction of Collagen Fibers and Delayed Re-Epithelialization in HMGB1+/− Wound
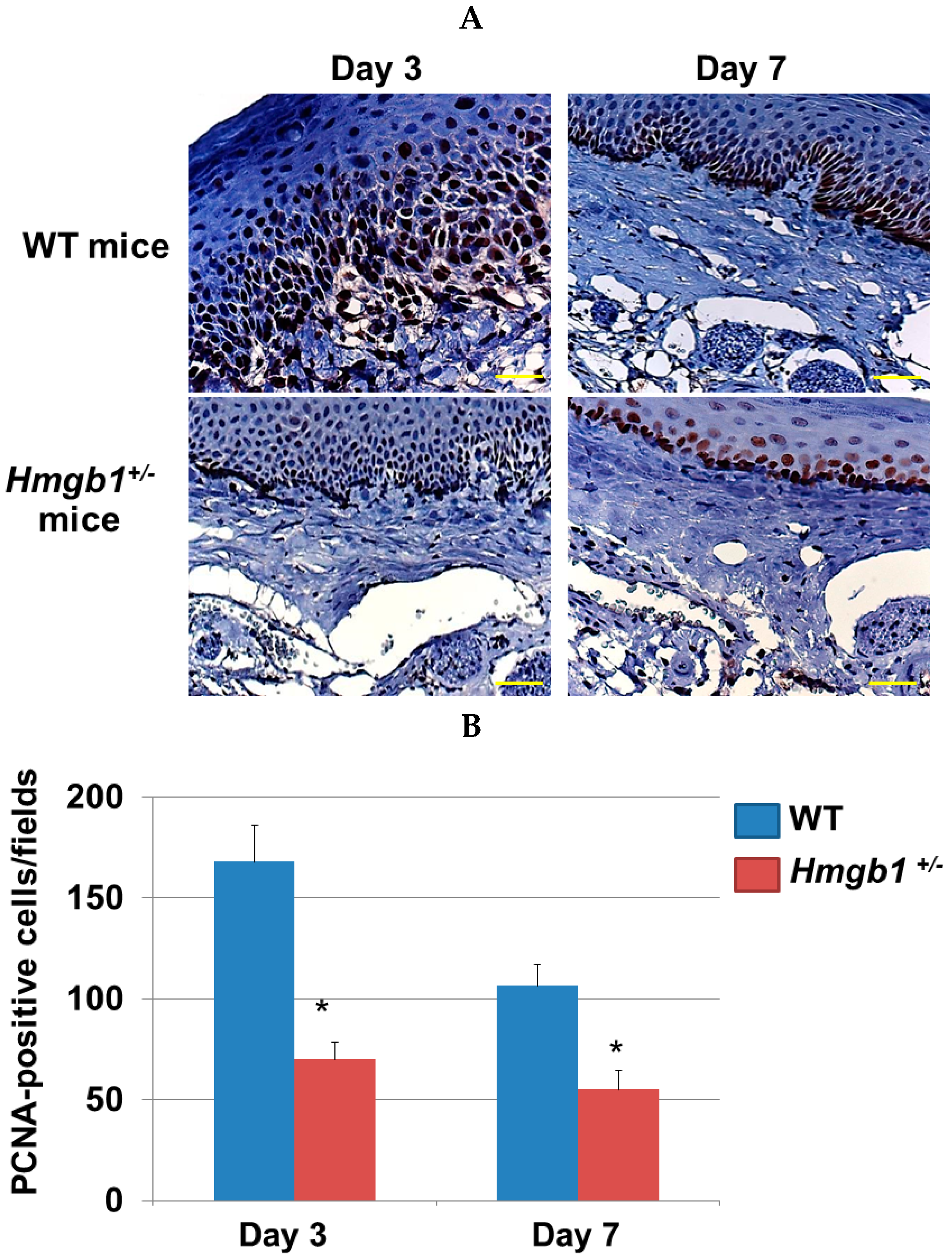
2.4. Immunohistochemistry Determination of Proliferating Cells at Palatal Wounds in WT and Hmgb1+/− Mice
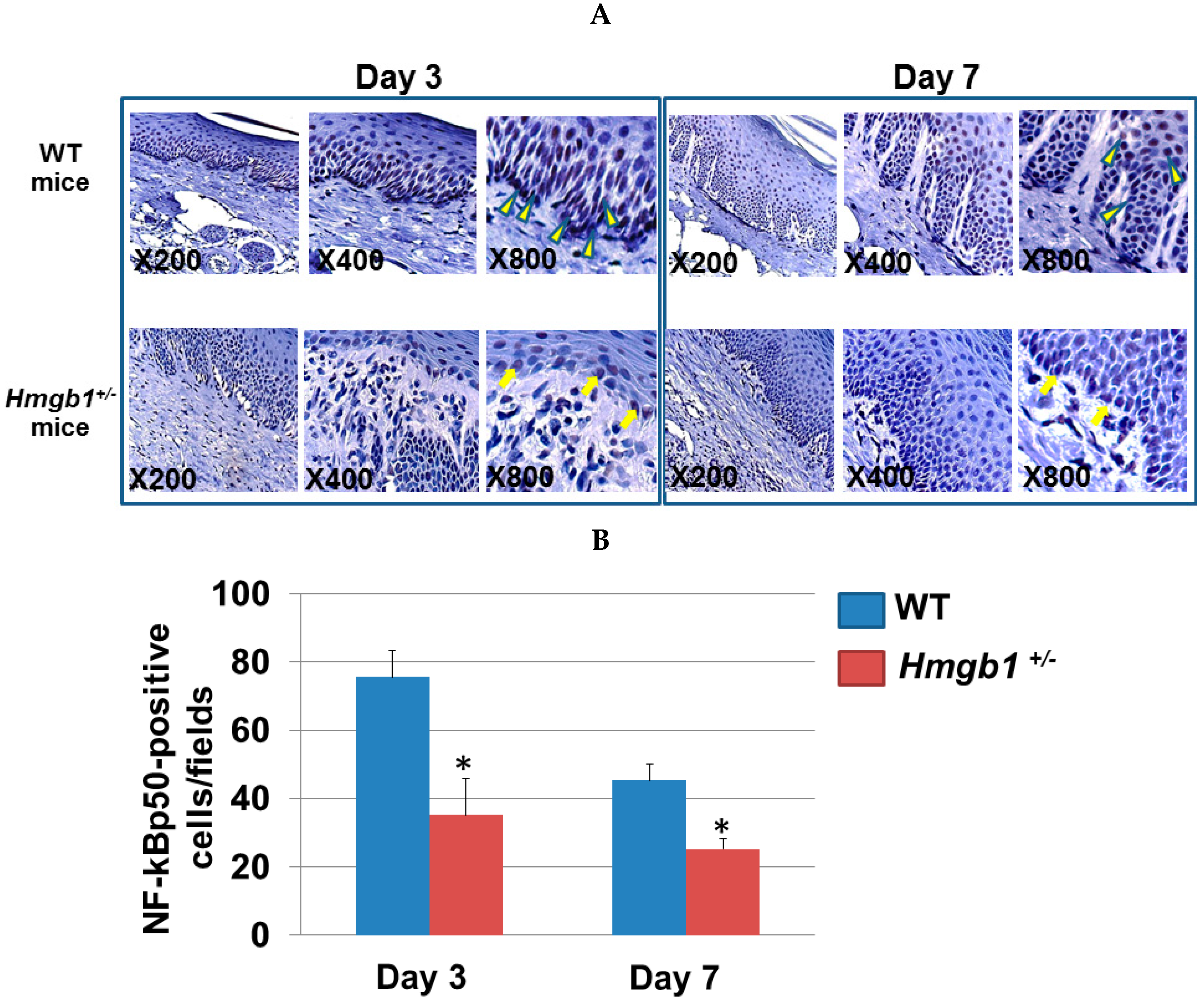
2.5. Localization of NF-κB p50 Isoform at Palatal Wounds in WT and Hmgb1+/− Mice
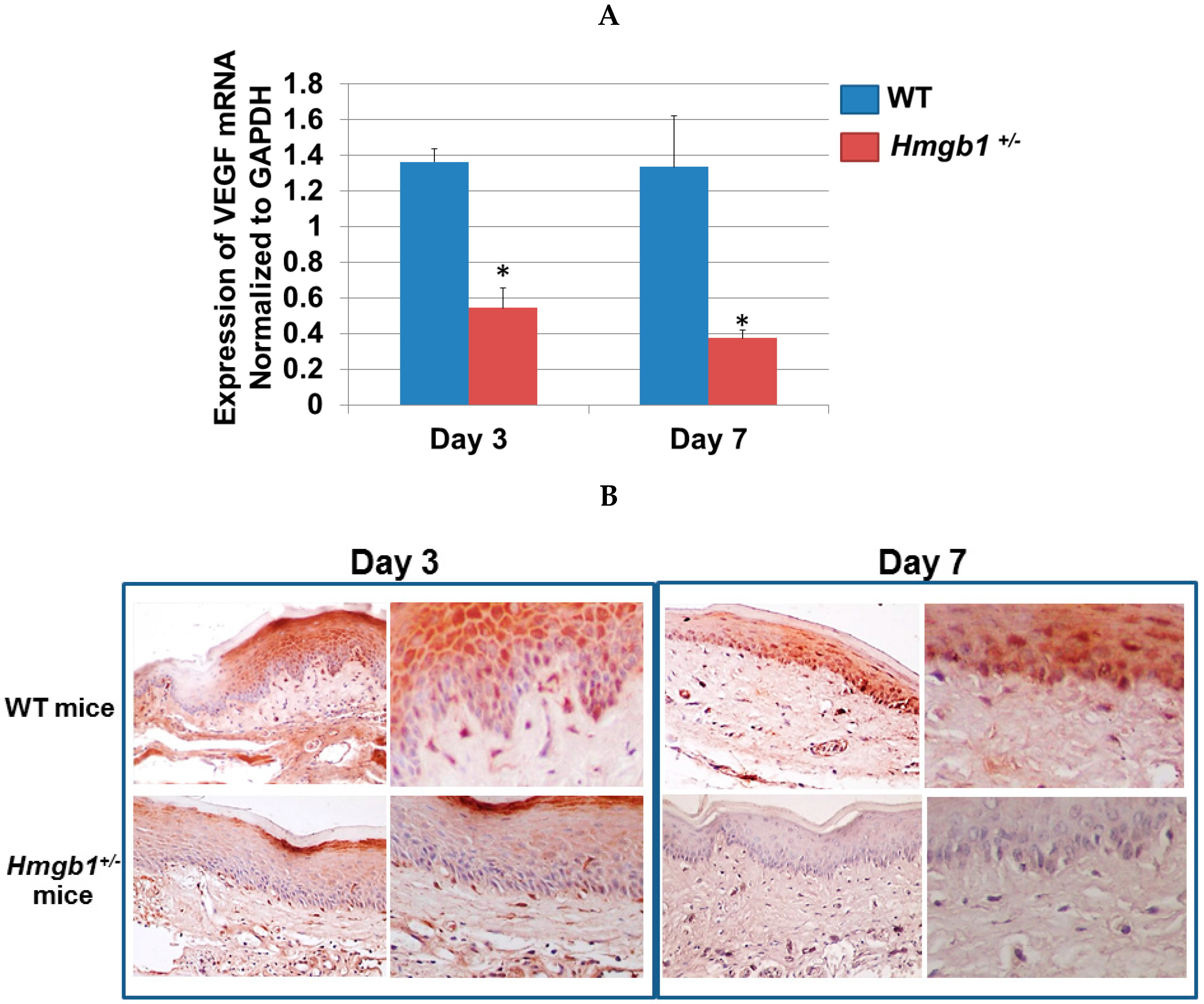
2.6. Determination of VEGF Expression and Localization in Palatal Wounds of WT and Hmgb1+/− Mice
2.7. Efficiency of RAGE Gene Knockdown
2.8. RAGE Silencing Reduces Closure of an In Vitro Scratch Wound and PCNA Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Mice
4.3. Polymerase Chain Reaction (PCR) Genotyping Assay
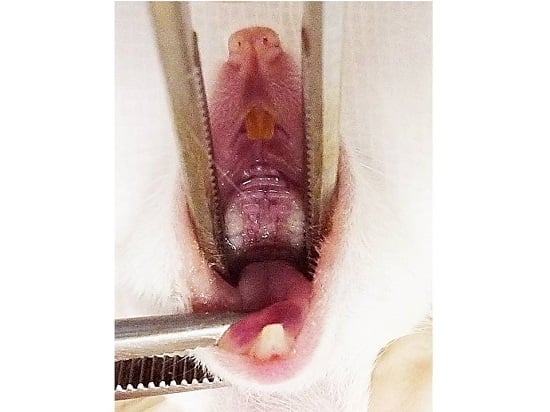
4.4. Palatal Wound Healing Model and Histological Analysis
4.5. Immunohistochemistry
4.6. Quantitative Real-Time PCR Analysis of VEGF Expression in the Wound Tissues
4.7. Primary Cell Cultures
4.8. Silencing of RAGE Gene Expression and Western Blot Detection for RAGE Protein Expression
4.9. Cell Viability Test
4.10. In Vitro Scratch Assay
4.11. Quantitative Real-Time PCR Analysis of PCNA Expression in the Gingival Epithelial Cells
4.12. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guo, S.; di Pietro, L.A. Factors Affecting Wound Healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Leoni, G.; Neumann, P.A.; Sumagin, R.; Denning, T.L.; Nusrat, A. Wound repair: Role of immune-epithelial interactions. Mucosal Immunol. 2015, 8, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Martin, P. Wound healing—Aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [PubMed]
- Mori, R.; Ohshima, T.; Ishida, Y.; Mukaida, N. Accelerated wound healing in tumor necrosis factor receptor p55-deficient mice with reduced leukocyte infiltration. FASEB. J. 2002, 16, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Kondo, T.; Takayasu, T.; Iwakura, Y.; Mukaida, N. The essential involvement of cross-talk between IFN-gamma and TGF-β in the skin wound-healing process. J. Immunol. 2004, 172, 1848–1855. [Google Scholar] [CrossRef] [PubMed]
- Gallucci, R.M.; Simeonova, P.P.; Matheson, J.M. Impaired cutaneous wound healing in interleukin-6-deficient and immunosuppressed mice. FASEB. J. 2000, 14, 2525–2531. [Google Scholar] [CrossRef] [PubMed]
- Jinno, K.; Takahashi, T.; Tsuchida, K.; Tanaka, E.; Moriyama, K. Acceleration of palatal wound healing in Smad3-deficient mice. J. Dent. Res. 2009, 88, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Werner, S.; Bugnon, P. Accelerated wound closure in mice deficient for interleukin-10. Am. J. Pathol. 2007, 170, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. HMG proteins: Dynamic players in gene regulation and differentiation. Curr. Opin. Genet. Dev. 2005, 15, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Oyama, Y.; Hashiguchi, T.; Taniguchi, N. High-mobility group box-1 protein promotes granulomatous nephritis in adenine-induced nephropathy. Lab. Investig. 2010, 90, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, N.; Kawahara, K.; Yone, K. High mobility group box chromosomal protein 1 plays a role in the pathogenesis of rheumatoid arthritis as a novel cytokine. Arthritis Rheum. 2003, 48, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Uchikado, H.; Miura, N. HMGB1 as a therapeutic target in spinal cord injury: A hypothesis for novel therapy development. Exp. Ther. Med. 2011, 2, 767–770. [Google Scholar] [PubMed]
- Morimoto, Y.; Kawahara, K.I.; Tancharoen, S. Tumor necrosis factor-α stimulates gingival epithelial cells to release high mobility-group box 1. J. Periodontal Res. 2008, 43, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, H.J.; Kuja-Panula, J.; Rauvala, H. Receptor for advanced glycation end products (RAGE) signaling induces CREB-dependent chromogranin expression during neuronal differentiation. J. Biol. Chem. 2002, 277, 38635–38646. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, R.; Sampaolesi, M.; de Marchis, F. Extracellular HMGB1, a signal of tissue damage, induces mesoangioblast migration and proliferation. J. Cell Biol. 2004, 164, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limana, F.; Germani, A.; Zacheo, A. Exogenous high-mobility group box 1 protein induces myocardial regeneration after infarction via enhanced cardiac C-kit+ cell proliferation and differentiation. Circ. Res. 2005, 97, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Patrone, M.; Pedrazzi, M.; Burlando, B. HMGb1 promotes scratch wound closure of HaCaT keratinocytes via ERK1/2 activation. Mol. Cell. Biochem. 2009, 332, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Bhawal, U.K.; Sasahira, T. Involvement of HMGB1 and RAGE in IL-1β-induced gingival inflammation. Arch. Oral Biol. 2012, 57, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M.; Hofmann, M.; Taguchi, A.; Du, Y.S.; Stern, D.M. RAGE: A Multiligand Receptor Contributing to the Cellular Response in Diabetic Vasculopathy and Inflammation. Semin. Thromb. Hemost. 2000, 26, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.D. Receptor for age (RAGE) is a gene within the major histocompatibility class III region: implications for host response mechanisms in homeostasis and chronic disease. Front. Biosci. 2001, 6, D1151–D1160. [Google Scholar] [CrossRef] [PubMed]
- Hori, B.J.; Slattery, T.; Cao, R.; Zhang, J.; Chen, J.X.; Nagashima, M.; Lundh, E.R.; Vijay, S.; Nitecki, D. The receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin. Mediation of neurite outgrowth and co-expression of rage and amphoterin in the developing nervous system. J. Biol. Chem. 1995, 270, 25752–25761. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, S.D.; Fu, C. RAGE mediates a novel proinflammatory axis: A central cell surface receptor for S100/calgranulin polypeptides. Cell 1999, 97, 889–901. [Google Scholar] [CrossRef]
- Bierhaus, H.P.; Morcos, M.; Wendt, T.; Chavakis, T.; Arnold, B.; Stern, D.M.; Nawroth, P.P. Understanding RAGE, the receptor for advanced glycation end products. J. Mol. Med. (Berl.) 2005, 83, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, P.M.; Pedrazzi, M.; Burlando, B. HMGB1 promotes wound healing of 3T3 mouse fibroblasts via RAGE-dependent ERK1/2 activation. Cell. Biochem. Biophys. 2010, 57, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Straino, S.; di, C.; Mangoni, A. High-mobility group box 1 protein in human and murine skin: Involvement in wound healing. J. Investig. Dermatol. 2008, 128, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Chitanuwat, A.; Laosrisin, N.; Dhanesuan, N. Role of HMGB1 in proliferation and migration of human gingival and periodontal ligament fibroblasts. J. Oral Sci. 2013, 55, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Szczepanski, M.J.; Luczak, M.; Olszewska, E. Molecular signaling of the HMGB1/RAGE axis contributes to cholesteatoma pathogenesis. J. Mol. Med. 2015, 93, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Tancharoen, S.; Tengrungsun, T.; Suddhasthira, T.; Kikuchi, K.; Vechvongvan, N.; Tokuda, M.; Maruyama, I. Overexpression of Receptor for Advanced Glycation End Products and High-Mobility Group Box 1 in Human Dental Pulp Inflammation. Mediat. Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Chaichalotornkul, S.; Nararatwanchai, T.; Narkpinit, S. Secondhand smoke exposure-induced nucleocytoplasmic shuttling of HMGB1 in a rat premature skin aging model. Biochem. Biophys. Res. Commun. 2015, 456, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Ebe, N.; Iwasaki, K.; Iseki, S.; Okuhara, S.; Podyma-Inoue, K.A.; Terasawa, K.; Watanabe, A.; Akizuki, T.; Watanabe, H.; Yanagishita, M.; et al. Pocket epithelium in the pathological setting for HMGB1 release. J. Dent. Res. 2011, 90, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; O’Hearn, S.; Kavalukas, S.L.; Barbul, A. Role of high mobility group box 1 (HMGB1) in wound healing. J. Surg. Res. 2012, 176, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.O.; Jones, R.O. The role of vascular endothelial growth factor in wound healing. Int. J. Lower Extremity Wounds 2003, 2, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Bueno, F.G.; Moreira, E.A.; Morais, G.R.; Pacheco, I.A.; Baesso, M.L.; Leite-Mello, E.V.; Mello, J.C. Enhanced cutaneous wound healing in vivo by standardized crude extract of poincianella pluviosa. PLoS ONE 2016, 11, e0149223. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, J.; Tam, B.Y.; Sundram, U.; von Degenfeld, G.; Blau, H.M.; Kuo, C.J.; Cooke, J.P. Discordant effects of a soluble VEGF receptor on wound healing and angiogenesis. Gene Ther. 2004, 11, 302–409. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Triveni, M.G.; Kumar, A.B.; Mehta, D.S. Role of platelet-rich-fibrin in enhancing palatal wound healing after free graft. Contemp. Clin. Dent. 2012, 3, S240–S243. [Google Scholar] [PubMed]
- Biscetti, F.; Straface, G.; de, C.R. High-Mobility Group Box-1 protein promotes angiogenesis after peripheral ischemia in diabetic mice through a VEGF-dependent mechanism. Diabetes 2010, 59, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Matthias, S. Cancer as an overhealing wound: An old hypothesis revisited. Nat. Rev. Mol. Cell Biol. 2008, 9, 628–638. [Google Scholar]
- Tang, D.; Shi, Y.; Kang, R. Hydrogen peroxide stimulates macrophages and monocytes to actively release HMGB1. J. Leukoc. Biol. 2007, 81, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Mohammedsaeed, W.; Cruickshank, S.; McBain, A.J.; O’Neill, C.A. Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration. Sci. Rep. 2015, 5, 16147. [Google Scholar] [CrossRef] [PubMed]
- Rossini, A.; Zacheo, A.; Mocini, D. HMGB1-stimulated human primary cardiac fibroblasts exert a paracrine action on human and murine cardiac stem cells. J. Mol. Cell. Cardiol. 2008, 44, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.G.; Zhang, H.; Yang, P.T. HMGB1 activates nuclear factor-kappaB signaling by RAGE and increases the production of TNF-α in human umbilical vein endothelial cells. Immunobiology 2010, 215, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Yi, P.P.; Zhou, R.R. The role of HMGB1-RAGE axis in migration and invasion of hepatocellular carcinoma cell lines. Mol. Cell. Biochem. 2014, 390, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Ruochan, C.; Qiuhong, Z.; Rui, K.; Xue-Gong, F.; Daolin, T. Emerging roles for HMGB1 protein in immunity, inflammation, and cancer. Mol. Med. 2013, 19, 357–366. [Google Scholar]
- Diabetic, R.; Dorria, M.; Elbardisey, H.M.; Eltokhy, M.; Doaa, T. Effect of transforming growth factor β 1 on wound healing in induced diabetic rats. Int. J. Health Sci. 2013, 7, 160–172. [Google Scholar]
- Calogero, S.; Grassi, F.; Aguzzi, A.; Voigtländer, T.; Ferrier, P.; Ferrari, S.; Bianchi, M.E. The lack of chromosomal protein HMG1 does not disrupt cell growth but causes lethal hypoglycaemia in newborn mice. Nat. Genet. 1999, 22, 276–280. [Google Scholar] [PubMed]
- Silva, C.O.; Ribeiro Edel, P.; Sallum, A.W.; Tatakis, D.N. Free gingival grafts: Graft shrinkage and donor-site healing in smokers and non-smokers. J. Periodontol. 2010, 81, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Boucek, R.J. Factors affecting wound healing. Otolaryngol. Clin. N. Am. 1984, 17, 243–264. [Google Scholar]
- Jettanacheawchankit, S.; Sasithanasate, S.; Sangvanich, P.; Banlunara, W.; Thunyakitpisal, P. Acemannan stimulates gingival fibroblast proliferation; expressions of keratinocyte growth factor-1, vascular endothelial growth factor, and type I collagen; and wound healing. J. Pharm. Sci. 2009, 109, 525–531. [Google Scholar] [CrossRef]
- Tancharoen, S.; Matsuyama, T.; Kawahara, K. Cleavage of host cytokeratin-6 by lysine-specific gingipain induces gingival inflammation in periodontitis patients. PLoS ONE 2015, 10, e0117775. [Google Scholar] [CrossRef] [PubMed]
- Merlo, S.; Canonico, P.L.; Sortino, M.A. Differential involvement of estrogen receptorα and estrogen receptor β in the healing promoting effect of estrogen in human keratinocytes. J. Endocrinol. 2009, 200, 189–197. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tancharoen, S.; Gando, S.; Binita, S.; Nagasato, T.; Kikuchi, K.; Nawa, Y.; Dararat, P.; Yamamoto, M.; Narkpinit, S.; Maruyama, I. HMGB1 Promotes Intraoral Palatal Wound Healing through RAGE-Dependent Mechanisms. Int. J. Mol. Sci. 2016, 17, 1961. https://doi.org/10.3390/ijms17111961
Tancharoen S, Gando S, Binita S, Nagasato T, Kikuchi K, Nawa Y, Dararat P, Yamamoto M, Narkpinit S, Maruyama I. HMGB1 Promotes Intraoral Palatal Wound Healing through RAGE-Dependent Mechanisms. International Journal of Molecular Sciences. 2016; 17(11):1961. https://doi.org/10.3390/ijms17111961
Chicago/Turabian StyleTancharoen, Salunya, Satoshi Gando, Shrestha Binita, Tomoka Nagasato, Kiyoshi Kikuchi, Yuko Nawa, Pornpen Dararat, Mika Yamamoto, Somphong Narkpinit, and Ikuro Maruyama. 2016. "HMGB1 Promotes Intraoral Palatal Wound Healing through RAGE-Dependent Mechanisms" International Journal of Molecular Sciences 17, no. 11: 1961. https://doi.org/10.3390/ijms17111961
APA StyleTancharoen, S., Gando, S., Binita, S., Nagasato, T., Kikuchi, K., Nawa, Y., Dararat, P., Yamamoto, M., Narkpinit, S., & Maruyama, I. (2016). HMGB1 Promotes Intraoral Palatal Wound Healing through RAGE-Dependent Mechanisms. International Journal of Molecular Sciences, 17(11), 1961. https://doi.org/10.3390/ijms17111961