
Adjuvant Effect of Quillaja saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus maximus) upon Immersion Vaccination

Abstract
:
1. Introduction
2. Results
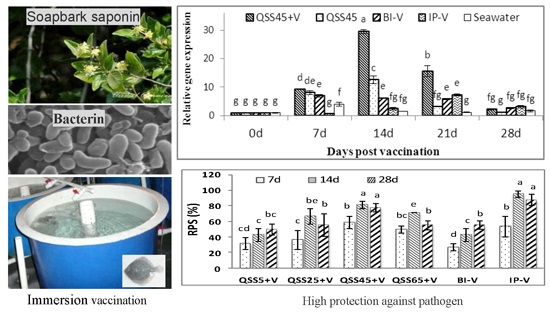
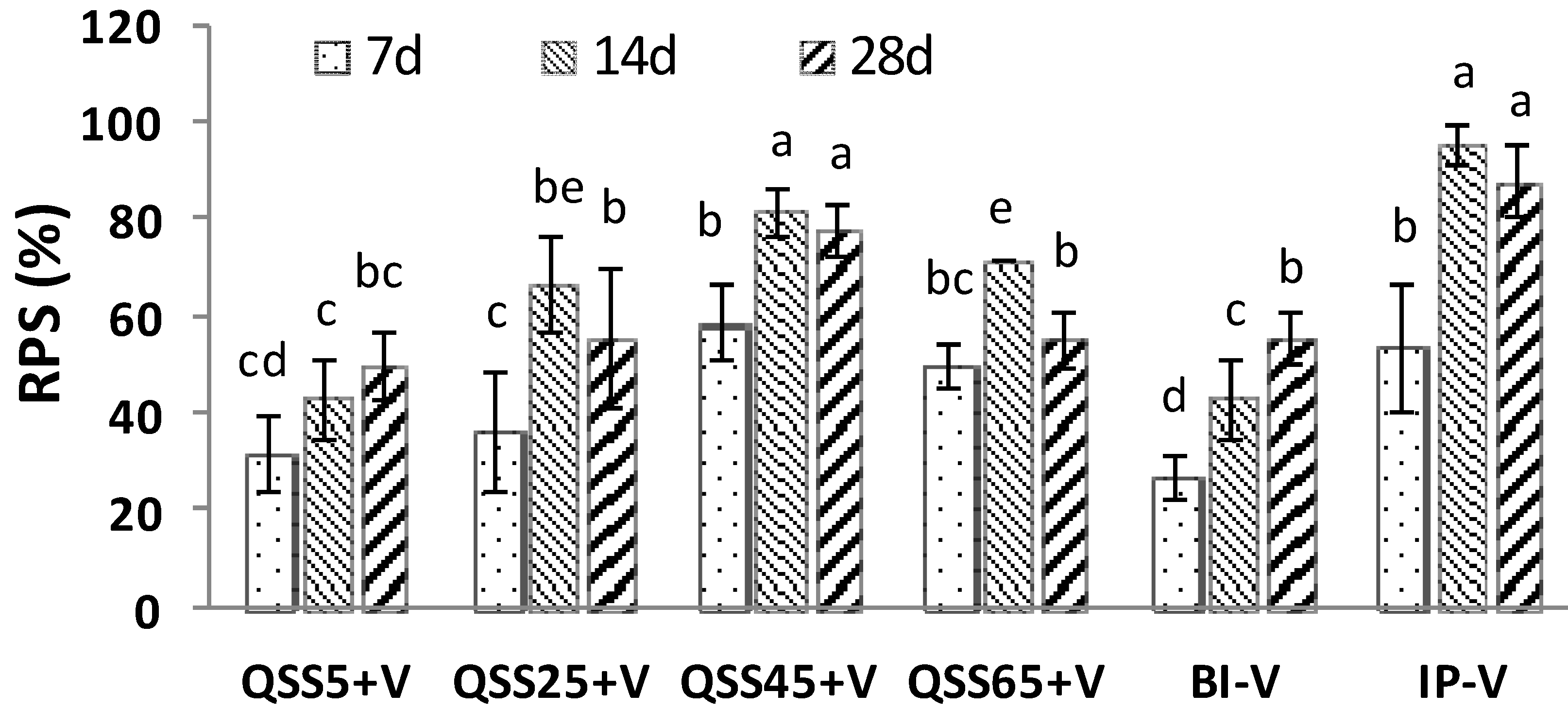
2.1. Challenge Experiments
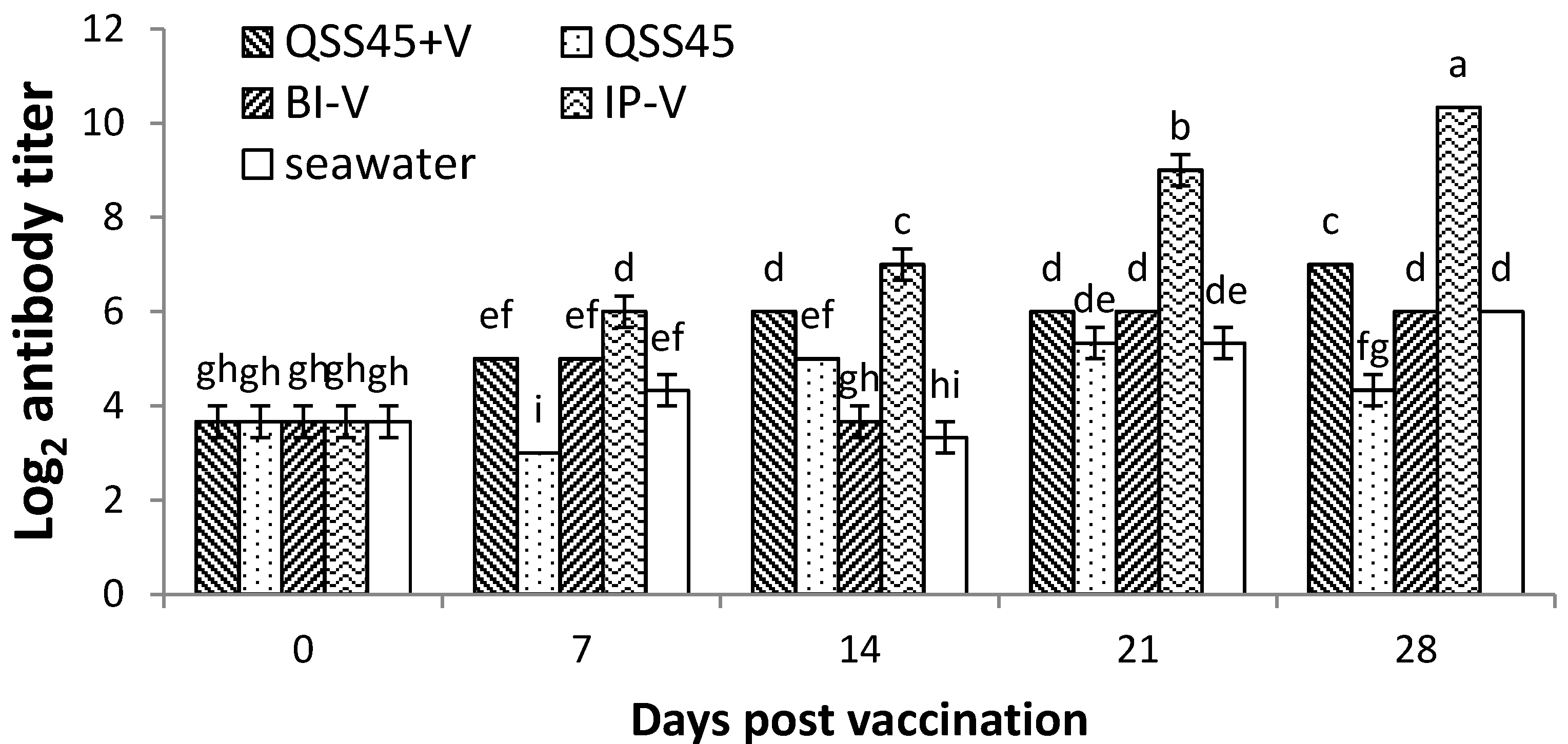
2.2. Antibody Titers
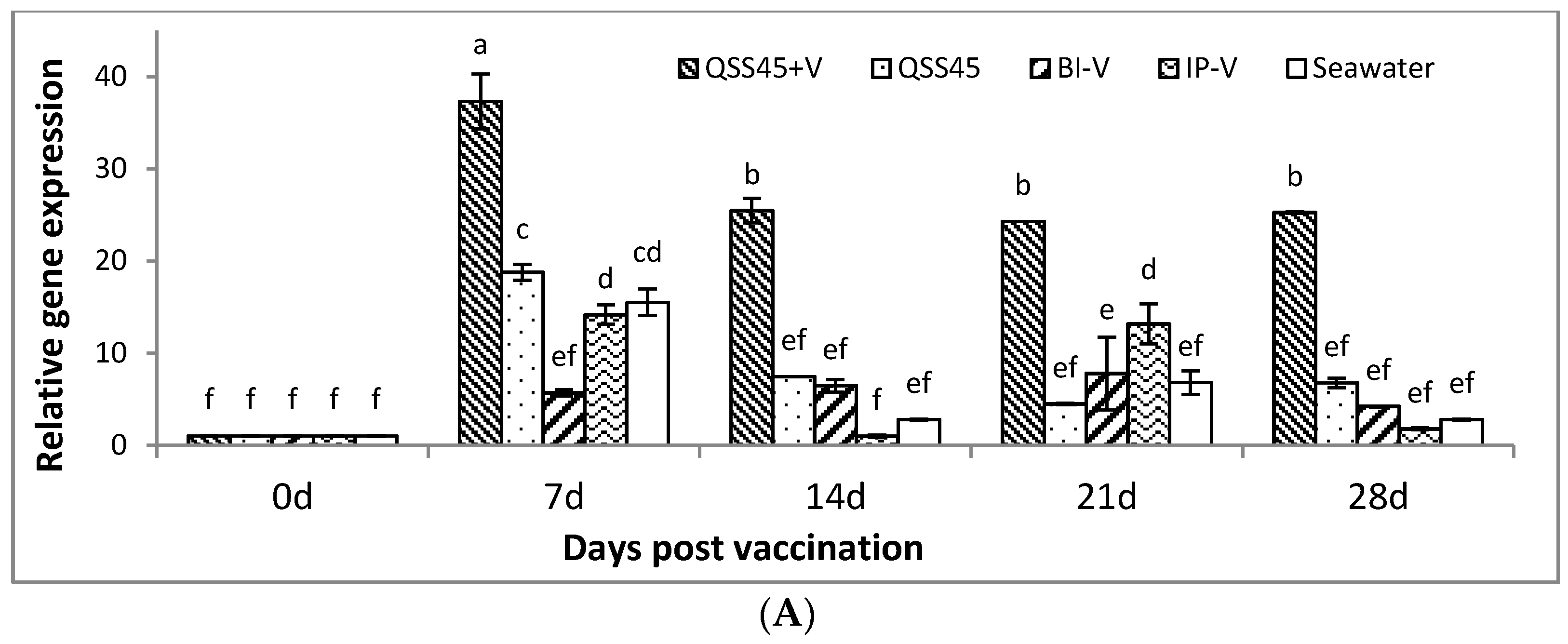
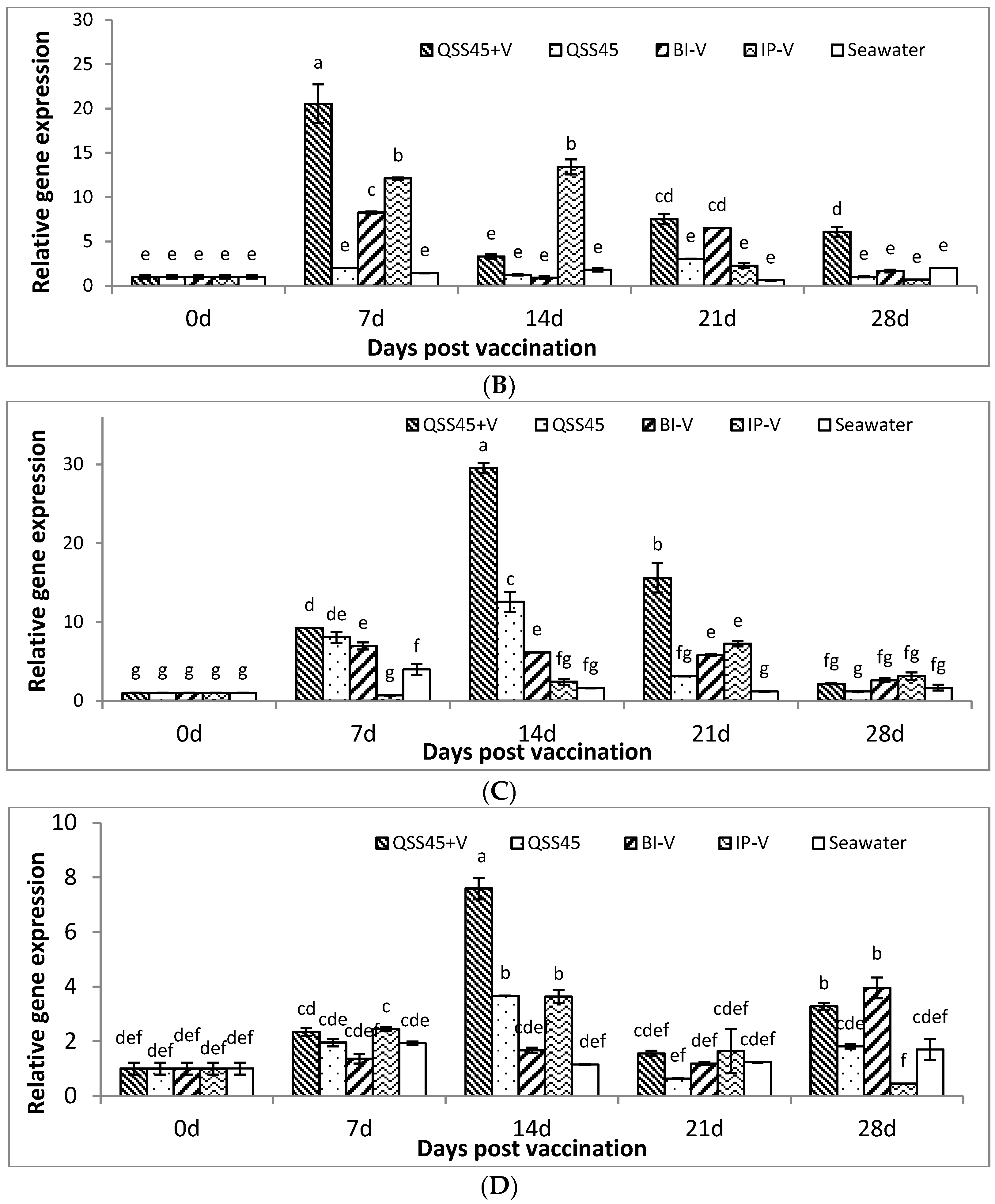
2.3. Expression of IgM mRNA in Tissues
3. Discussion
4. Materials and Methods
4.1. Fish
4.2. Saponins
4.3. Bacterial Strains
4.4. Vaccine Preparation
4.5. Vaccination Procedure
4.6. Challenges
4.7. Sample Collection
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. RNA Extraction
4.10. Reverse Transcription
4.11. Quantitative Real-Time PCR (qPCR)
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Kibenge, F.S.B.; Godoy, M.G.; Fast, M.; Workenhe, S.; Kibenge, M.J.T. Countermeasures against viral diseases of farmed fish. Antivir. Res. 2012, 95, 257–281. [Google Scholar] [CrossRef] [PubMed]
- Vinitantharat, S.; Gravningen, K.; Greger, E. Fish vaccines. Adv. Vet. Med. 1999, 41, 539–550. [Google Scholar]
- Sommerset, I.; Krossøy, B.; Biering, E.; Frost, P. Vaccines for fish in aquaculture. Expert Rev. Vaccines 2005, 4, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Toranzo, A.E.; Devesa, S.; Romalde, J.L.; Lamas, J.; Riaza, A.; Leiro, J.; Barja, J.L. Efficacy of intraperitoneal and immersion vaccination against Enterococcus sp. infection in turbot. Aquaculture 1995, 134, 17–27. [Google Scholar] [CrossRef]
- Esteve-Gassent, M.D.; Fouz, B.; Amaro, C. Efficacy of a bivalent vaccine against eel diseases caused by Vibrio vulnificus after its administration by four different routes. Fish Shellfish Immunol. 2004, 16, 93–105. [Google Scholar] [CrossRef]
- Pasquale, A.D.; Preiss, S.; de Silva, F.T.; Garcon, N. Vaccine adjuvants: From 1920 to 2015 and beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef] [PubMed]
- Stills, H.F. Adjuvants and antibody production: Dispelling the myths associated with Freund’s complete and othe adjuvants. ILAR J. 2015, 46, 280–293. [Google Scholar] [CrossRef]
- Song, X.; Hu, S. Adjuvant activities of saponins from traditional Chinese medicinal herbs. Vaccine 2009, 27, 4883–4890. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Milgate, J.; Roberts, D.C.K. The nutritional & biological significance of saponins. Nutr. Res. 1995, 15, 1223–1249. [Google Scholar]
- Campbell, J.B. Saponins. In The Theory and Practical Application of Adjuvants; Stewart-Tull, D.E.S., Ed.; John Wiley & Sons Ltd.: New York, NY, USA, 1995; pp. 95–127. [Google Scholar]
- Yoshiki, Y.; Kudou, S.; Okubo, K. Relationship between chemical structures and biological activities of triterpenoid saponins from soybean. Biosci. Biotechnol. Biochem. 1998, 62, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Riguera, R. Isolating bioactive compounds from marine organisms. J. Mar. Biotechnol. 1997, 5, 187–193. [Google Scholar]
- Charles, G.; Jean, L.; Karl, G.L.; Vakhtang, M.; André, P. Haemolytic activity, cytotoxicity and membrane cell permeabilization of semi-synthetic and natural lupane-and oleanane-type saponins. Bioorg. Med. Chem. 2009, 17, 2002–2008. [Google Scholar]
- Yang, Z.G.; Sun, H.X.; Fang, W.H. Haemolytic activities and adjuvant effect of Astragalus membranaceus saponins (AMS) on the immune responses to ovalbumin in mice. Vaccine 2005, 23, 5196–5203. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, M.; Liu, J. Haemolytic activities and adjuvant effect of Anemone raddeana saponins (ARS) on the immune responses to ovalbumin in mice. Int. Immunopharmacol. 2008, 8, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Cui, Y.; Huang, J.J.; Zhang, Y.Z.; Nie, Z.; Wang, L.F.; Yan, B.Z.; Tang, Y.L.; Liu, Y. Immuno-stimulating properties of diosgenyl saponins isolated from Paris polyphylla. Bioorg. Med. Chem. Lett. 2007, 17, 2408–2413. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.I.; Roner, M.R. Characterization of in vivo anti-rotavirus activities of saponin extracts from Quillaja saponaria Molina. Antivir. Res. 2011, 90, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Deng, W.; Sun, H.; Li, D. Platycodin D2 is a potential less hemolytic saponin adjuvant eliciting Th1 and Th2 immune responses. Int. Immunopharmacol. 2008, 8, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.J.; Wu, J.Y.; Gardner, B.H.; Anderson, C.A.; Kensil, C.R.; Recchia, J.; Coughlin, R.T.; Powell, M.F. Induction of cross-reactive cytotoxic T-lymphocyte responses specific for HIV-1 gp120 using saponin adjuvant (QS-21) supplemented subunit vaccine formulations. Vaccine 1997, 15, 1001–1007. [Google Scholar] [CrossRef]
- Marciani, D.J.; Pathak, A.K.; Reynolds, R.C.; Seitz, L.; May, R.D. Altered immunomodulating and toxicological properties of degraded Quillaja saponaria Molina saponins. Int. Immunopharmacol. 2001, 1, 813–818. [Google Scholar] [CrossRef]
- Santos, F.N.; Borja-Cabrera, G.P.; Miyashiro, L.M.; Grechi, J.; Reis, A.B.; Moreira, M.A.B.; Martins Filho, O.A.; Luvizotto, M.C.R.; Menz, I.; Pessôa, L.M.; et al. Immunotherapy against experimental canine visceral leishmaniasis with the saponin enriched-Leishmune® vaccine. Vaccine 2007, 25, 6176–6190. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C( T )) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Freitas, E.; Casas, C.P.; Borja-Cabrera, G.P.; Santos, F.N.; Nico, D.; Souza, L.O.P.; Tinoco, L.W.; da Silva, B.P.; Palatnik, M.; Parente, J.P.; et al. Acylated and deacylated saponins of Quillaja saponaria mixture as adjuvants for the FML-vaccine against visceral leishmaniasis. Vaccine 2006, 24, 3909–3920. [Google Scholar] [CrossRef] [PubMed]
- Su, B.K.; Chen, J.C. Effect of saponin immersion on enhancement of the immune response of white shrimp Litopenaeus vannamei and its resistance against Vibrio alginolyticus. Fish Shellfish Immunol. 2008, 24, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Makkar, H.P.S.; Becker, K. Quillaja saponins-a natural growth promoter for fish. Anim. Feed Sci. Technol. 2005, 121, 147–157. [Google Scholar] [CrossRef]
- Han, W.; Wang, X.H.; Wang, Y.J.; Wang, R.; Li, C.; Huang, J. Effect of saponin on non-specific immunity of turbot (Scophthamus maximus). J. Fish. China 2014, 38, 999–1006. [Google Scholar]
- Fuentes-Appelgren, P.; Opazo, R.; Barros, L.; Feijoó, C.G.; Urzua, V.; Romero, J. Effect of the dietary inclusion of soybeen components on the innate immune system in zebrafish. Zebrafish 2014, 11, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Marciani, D.J. Vaccine adjuvants: Role and mechanisms of action in vaccine immunogenicity. Drug Discov. Today 2003, 8, 934–943. [Google Scholar] [CrossRef]
- Amend, D.F.; Fender, D.C. Uptake of bovine serum albumin by rainbow trout from hyperosmotic solutions: A model for vaccinating fish. Science 1976, 192, 793–794. [Google Scholar] [CrossRef] [PubMed]
- Bagherwal, P. Phytosaponin adjuvants: A better option for vaccines. Int. J. PharmTech Res. 2011, 3, 1837–1842. [Google Scholar]
- Sasaki, S.; Sumino, K.; Hamajima, K.; Fukushima, J.; Ishii, N.; Kawamoto, S.; Mohri, H.; Kensil, C.R.; Okuda, K. Induction of systemic and mucosal immune responses to human immunodeficiency virus type 1 by a DNA vaccine formulated with QS-21 saponin adjuvant via intramuscular and intranasal routes. J. Virol. 1998, 72, 4931–4939. [Google Scholar] [PubMed]
- Oda, K.; Matsuda, H.; Murakami, T.; Katayama, S.; Ohgitani, T.; Yoshikawa, M. Relationship between adjuvant activity and amphipathic structure of soyasaponins. Vaccine 2003, 21, 2145–2151. [Google Scholar] [CrossRef]
- Sun, H.; Yang, Z.; Ye, Y. Structure and biological activity of protopanaxatriol-type saponins from the roots of Panax notoginseng. Int. Immunopharmacol. 2006, 6, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Crosbie, P.B.B.; Nowak, B.F. Immune responses of barramundi, Lates calcarifer (Bloch), after administration of an experimental Vibrio harveyi bacterin by intraperitoneal injection, anal intubation and immersion. J. Fish Dis. 2004, 27, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Joosten, P.H.M.; Tiemersma, E.; Threels, A.; Caumartin-Dhieux, C.; Rombout, J.H.W.M. Oral vaccination of fish against Vibrio anguillarum using alginate microparticles. Fish Shellfish Immunol. 1997, 7, 471–485. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Bath vaccination of rainbow trout (Oncorhynchus mykiss Walbaum) against Yersinia ruckeri: Effects of temperature on protection and gene expression. Vaccine 2008, 26, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Kaattari, I.M.; Ma, C.; Kaattari, S. The teleost humoral immune response. Fish Shellfish Immunol. 2013, 35, 1719–1728. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.M.; Zhan, W.B.; Sheng, X.Z.; Yang, K.; Han, J.G.; Wei, J.L.; Li, J.; Qiao, X.T. Response of mucosal and systemic sIgM-positive cells in turbot (Scophthalmus maximus L.) immunization with Edwardsiella tarda. Vet. Immunol. Immunopathol. 2009, 129, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Gassent, M.D.; Nielsen, M.E.; Amaro, C. The kinetics of antibody production in mucus and serum of European eel (Anguilla anguilla L.) after vaccination against Vibrio vulnificus: Development of a new method for antibody quantification in skin mucus. Fish Shellfish Immunol. 2003, 15, 51–61. [Google Scholar] [CrossRef]
- Tian, J.; Sun, B.; Luo, Y.; Zhang, Y.; Nie, P. Distribution of IgM, IgD and IgZ in mandarin fish, Siniperca chuatsi lymphoid tissues and their transcriptional changes after Flavobacterium columnare stimulation. Aquaculture 2009, 288, 14–21. [Google Scholar] [CrossRef]
- Vervarcke, S.; Ollevier, F.; Kinget, R.; Michoel, A. Mucosal response in African catfish after administration of Vibrio anguillarum O2 antigen via different routes. Fish Shellfish Immunol. 2005, 18, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Navot, N.; Kimmel, E.; Avtalion, R.R. Enhancement of antigen uptake and antibody production in goldfish (Carassius auratus) following bath immunization and ultrasound treatment. Vaccine 2004, 22, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Huang, J.; Wang, X.H. Comparative proteomics of two Vibrio anguillarum serotype O1 strains with different virulence phenotypes. Curr. Microbiol. 2012, 65, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Collado, R.; Fouz, B.; Sanjuán, E.; Amaro, C. Effectiveness of different vaccine formulations against vibriosis caused by Vibrio vulnificus serovar E (biotype 2) in European eels Anguilla anguilla. Dis. Aquat. Org. 2000, 43, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Wang, X.H.; Han, W.; Wang, R.; Huang, J. Preparation and application of polyclonal antibodies against five pathogenic bacteria isolated from flounder. Prog. Fish. Sci. 2013, 34, 68–74. [Google Scholar]
- Wang, X.; Zhou, L.; Wang, Y.; Liu, Q. Evaluation of immune effects of a multivalent vector vaccine in turbot, Scophthalmus maximus. J. Fish. Sci. China 2011, 18, 918–923. [Google Scholar] [CrossRef]
- Taylor, I.S.; Low, C.A.; Birkbeck, T.H.; Secombes, C.J.; Tatner, M.F.; Melvin, W.T. Turbot (Scophthalmus maximus) β-actin gene and mRNA. Unpublished work. 2000. [Google Scholar]
- Low, C.A.; Taylor, I.S.; Birkbeck, T.H.; Tatner, M.F.; Secombes, C.J. Scophthalmus maximus partial mRNA sequence for immunoglobulin M variable region. Unpublished work. 2000. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Cumulative Mortality (%) | ||
|---|---|---|---|
| 7 Days | 14 Days | 28 Days | |
| QSS5 + V | 50.0 ± 10.0 b,c,d | 40.0 ± 10 c,d | 30 ± 26.5 c |
| QSS5 | 60.0 ± 10.0 a,b,c | 63.3 ± 11.5 a,b | 60.0 ± 0 a,b |
| QSS25 + V | 46.7 ± 15.3 b,c,d | 23.3 ± 11.6 d,e | 26.7 ± 15.3 c |
| QSS25 | 63.3 ± 5.7 a,b,c | 56.7 ± 11.5 b,c | 60.0 ± 0 a,b |
| QSS45 + V | 30.0 ± 10.0 e,f | 13.3 ± 5.7 e,f | 13.3 ± 5.8 c |
| QSS45 | 66.7 ± 5.7 a,b,c | 63.3 ± 11.5 a,b | 53.3 ± 5.7 b |
| QSS65 + V | 30.7 ± 5.8 d,e,f | 20.0 ± 0 e,f | 26.7 ± 5.8 c |
| QSS65 | 60.0 ± 10.0 a,b,c | 53.3 ± 20.8 b,c | 50.0 ± 10.0 b |
| BI-V | 53.3 ± 5.8 b,c,d | 40.0 ± 10.0 c,d | 26.7 ± 5.7 c |
| Seawater | 73.3 ± 15.3 a | 70.0 ± 10.0 a,b | 60.0 ± 10.0 a,b |
| IP-V | 20.0 ± 10.0 f | 3.3 ± 5.7 f | 10.0 ± 10.0 c |
| PBS | 43.3 ± 15.3 c,d,e | 80.0 ± 10.0 a | 76.7 ± 5.8 a |
| Groups | Way of Immunization | No. of Fish (No. of Replicates) | Dosage of Vaccine (cfu/mL) |
|---|---|---|---|
| QSS5 + V | BI | 90(2) | 1 × 108 |
| QSS5 | BI | 90(2) | 0 |
| QSS25 + V | BI | 90(2) | 1 × 108 |
| QSS25 | BI | 90(2) | 0 |
| QSS45 + V | BI | 90(2) | 1 × 108 |
| QSS45 | BI | 90(2) | 0 |
| QSS65 + V | BI | 90(2) | 1 × 108 |
| QSS65 | BI | 90(2) | 0 |
| BI-V | BI | 90(2) | 1 × 108 |
| Seawater | BI | 90(2) | 0 |
| IP-V | IP | 90(2) | 1 × 108 |
| PBS | IP | 90(2) | 0 |
| Gene | Primers Used | Sequence(5′→3′) | Location on Partial Sequence |
|---|---|---|---|
| β-actin | β-actin F | AAGCTGTGCTGTCCCTGTATG | 311–331 |
| β-actin | β-actin R | GCAGTGGTGGTGAAGGAGTAG | 492–512 |
| IgM | IgM F | TCAGTATCGACTTAGACACTTGCAG | 70–94 |
| IgM | IgM R | TCCCCAGTAGTCAAAGATCCAC | 169–191 |
| Target Gene | Composition of Reaction Mixture (μL) | Cycling Protocol | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| cDNA (μL) | Forward Primer (10 μM) | Reverse Primer (10 μM) | TransStrat™ Green qPCRSuper Mix (2×) | Passive Reference Dye (50×) | Sterile Water | Denature | Anneal | Elongate | No. of Cycles | Product Size (bp) | |
| β-Actin | 2 | 0.5 | 0.5 | 12.5 | 0.5 | 9 | 95 °C/30 s | 1 | 202 | ||
| 95 °C/5 s | 55 °C/15 s | 72 °C/20 s | 40 | ||||||||
| IgM | 2 | 0.5 | 0.5 | 12.5 | 0.5 | 9 | 95 °C/30 s | 1 | 122 | ||
| 95 °C/5 s | 55 °C/15 s | 72 °C/20 s | 40 | ||||||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, X.; Huang, J.; Li, J. Adjuvant Effect of Quillaja saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus maximus) upon Immersion Vaccination. Int. J. Mol. Sci. 2016, 17, 325. https://doi.org/10.3390/ijms17030325
Wang Y, Wang X, Huang J, Li J. Adjuvant Effect of Quillaja saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus maximus) upon Immersion Vaccination. International Journal of Molecular Sciences. 2016; 17(3):325. https://doi.org/10.3390/ijms17030325
Chicago/Turabian StyleWang, Yujuan, Xiuhua Wang, Jie Huang, and Jun Li. 2016. "Adjuvant Effect of Quillaja saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus maximus) upon Immersion Vaccination" International Journal of Molecular Sciences 17, no. 3: 325. https://doi.org/10.3390/ijms17030325
APA StyleWang, Y., Wang, X., Huang, J., & Li, J. (2016). Adjuvant Effect of Quillaja saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus maximus) upon Immersion Vaccination. International Journal of Molecular Sciences, 17(3), 325. https://doi.org/10.3390/ijms17030325