
Beyond the Niche: Myelodysplastic Syndrome Topobiology in the Laboratory and in the Clinic


Abstract
:1. Introduction
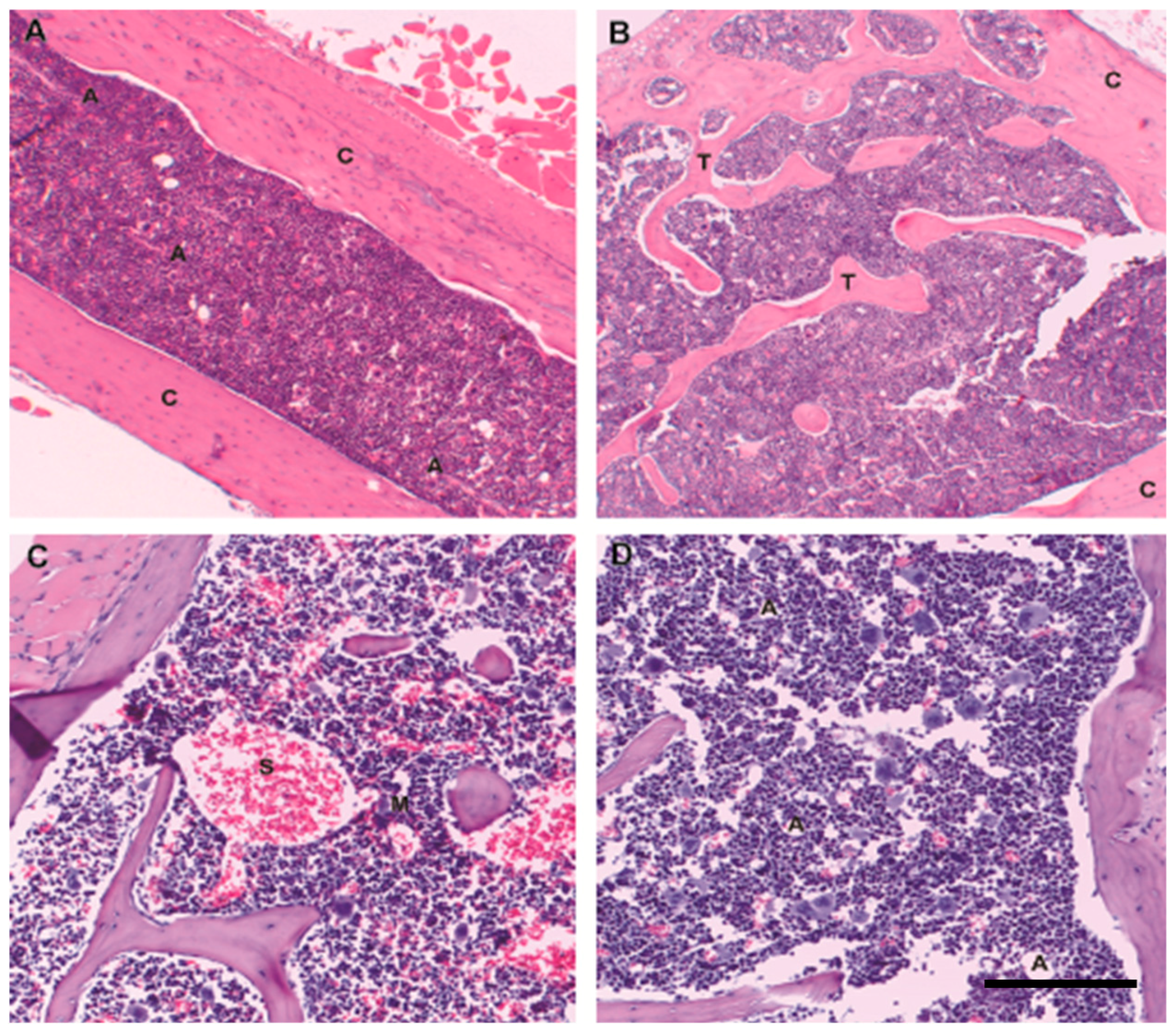
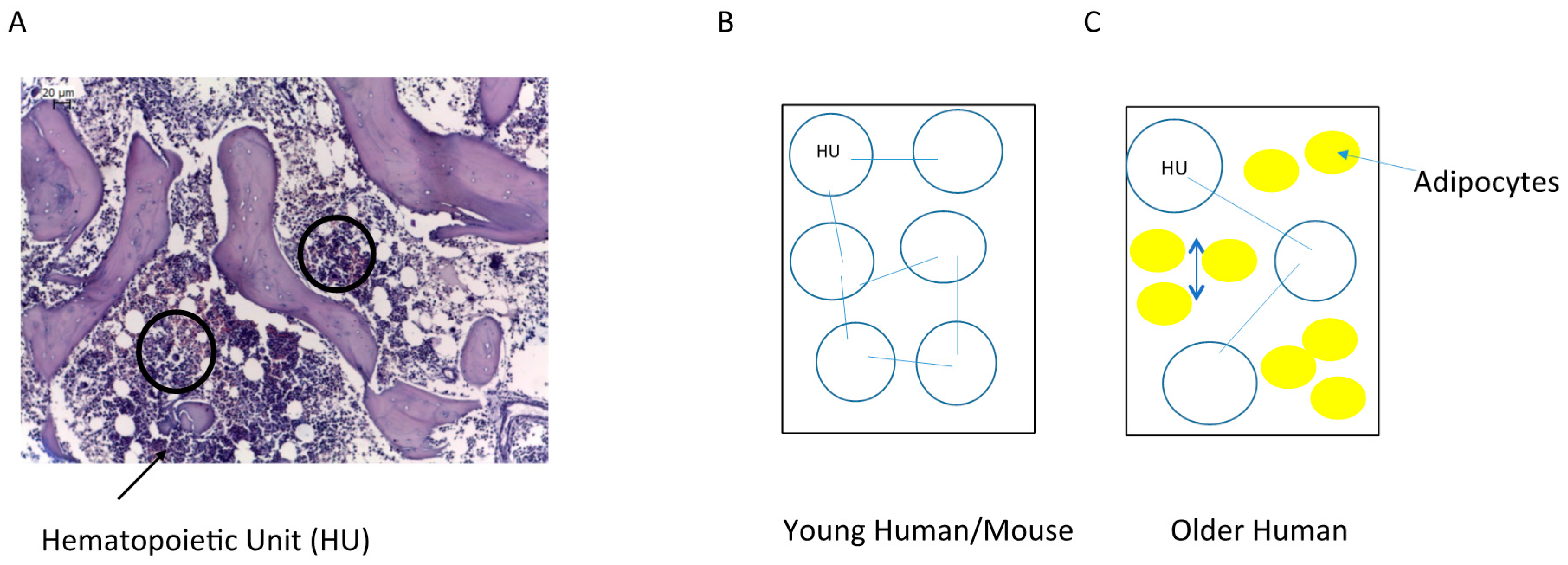
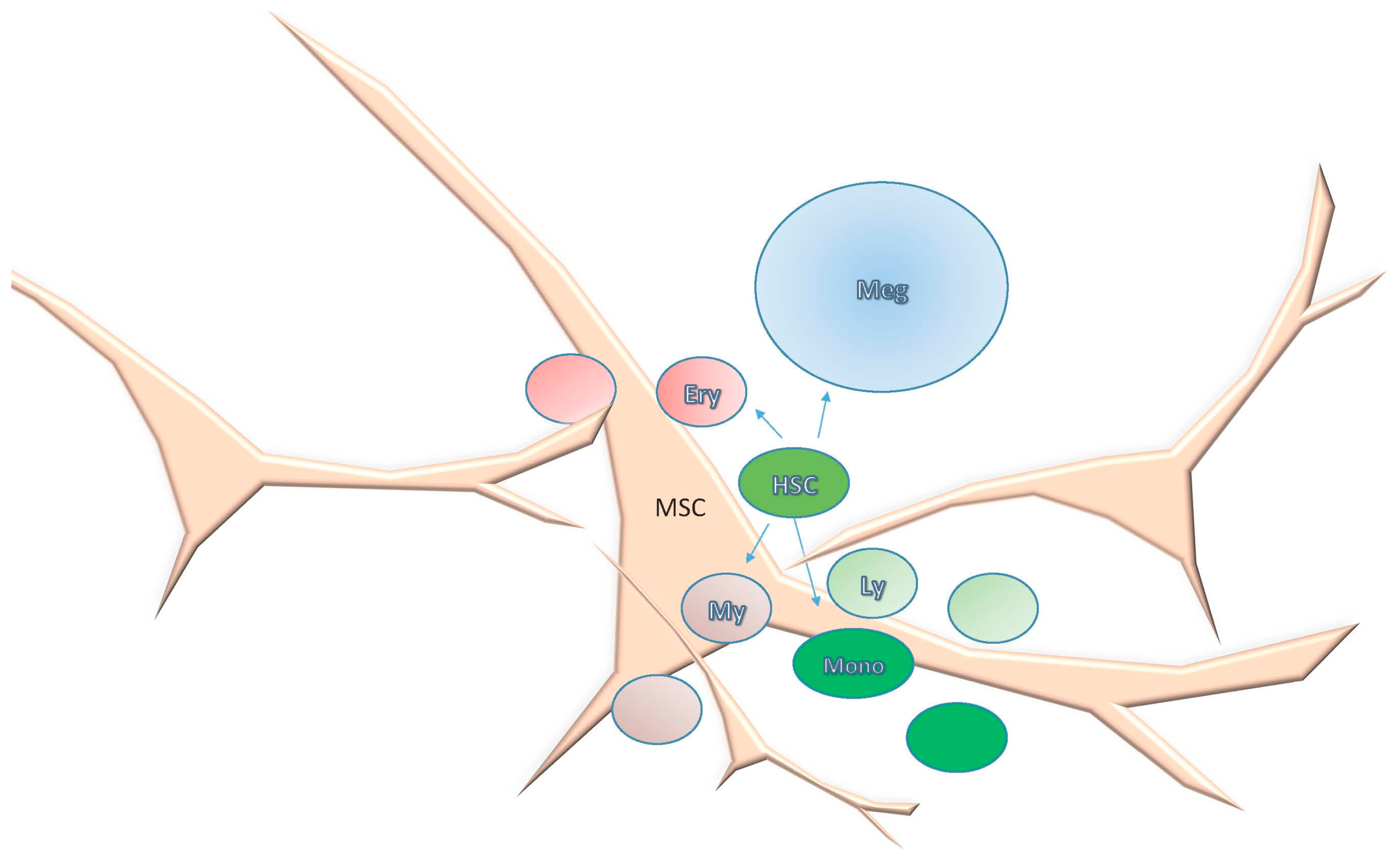
2. Bone Marrow Architecture in Man and Mouse
3. The Niche/Microenvironment in Drosophila and in the Mouse
3.1. The Stem Cell Niche Concept
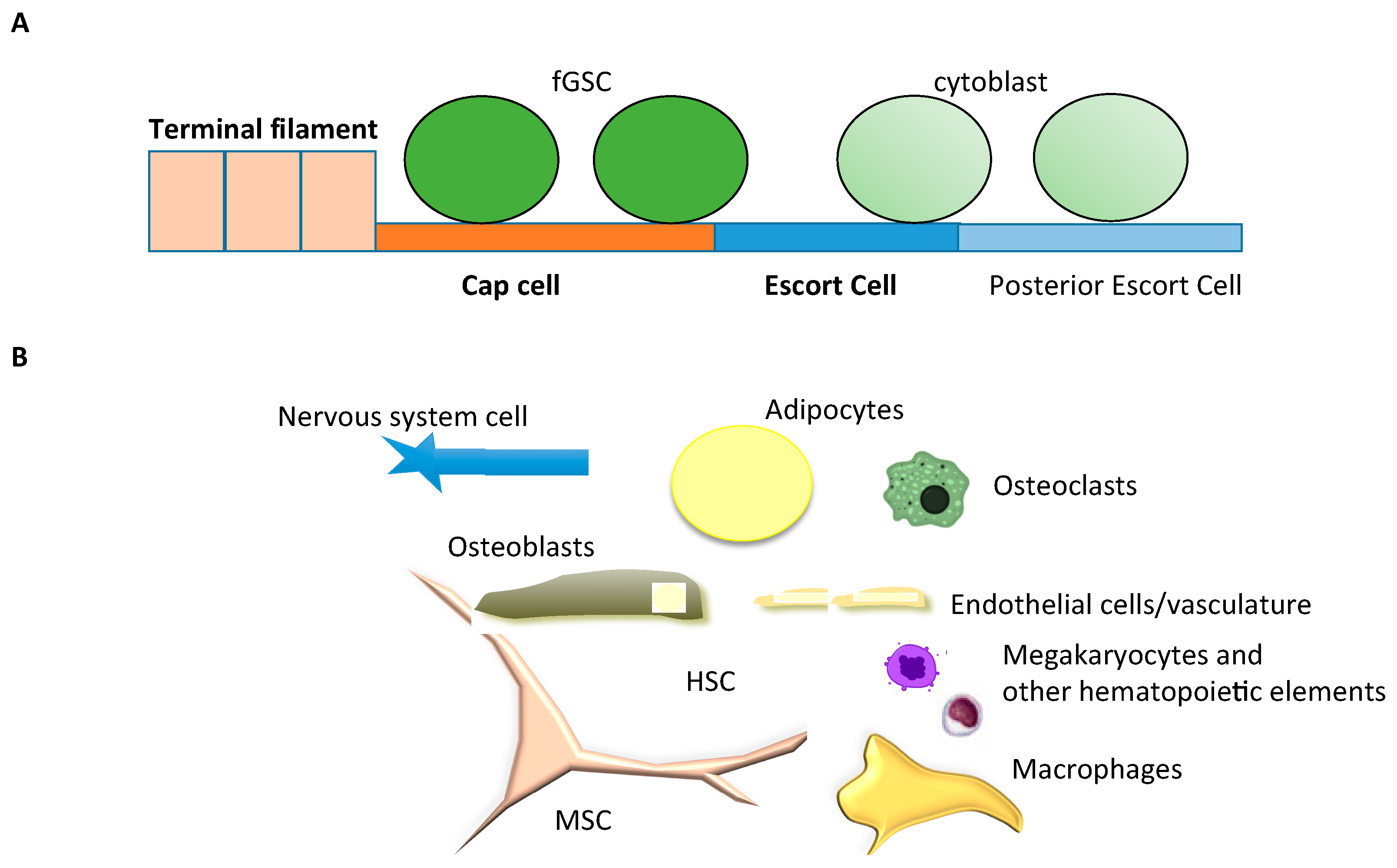
3.2. Drosophila Germinal Stem Cell Niche, a Fixed Niche
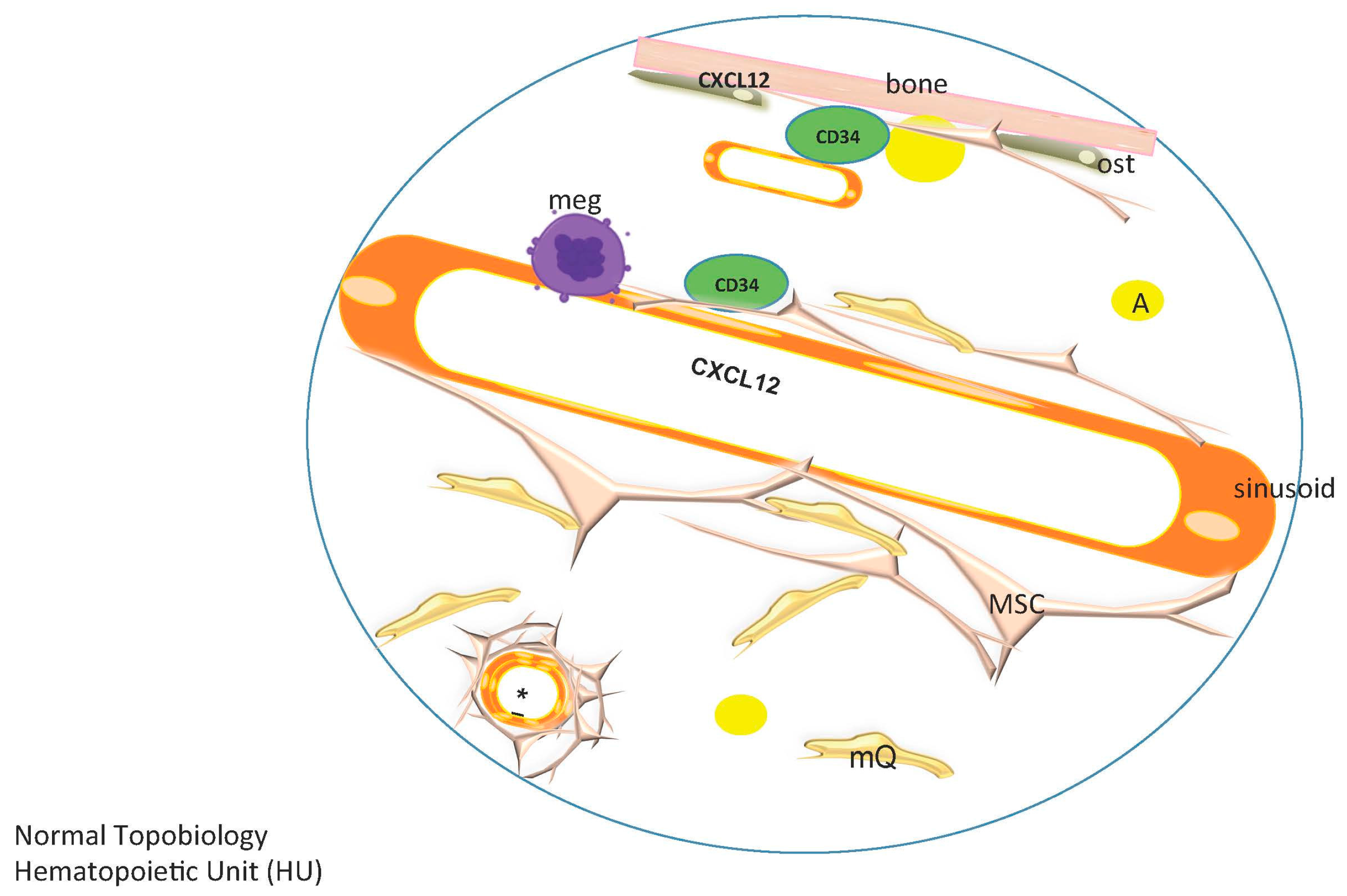
3.3. The Hematopoietic Stem Cell Niche in the Mouse, a Dynamic and Complex Niche
3.4. HSC Niche Is Dynamic
3.5. HSC Niche Is Complex
3.6. The HSC Niche Is Not Exclusive
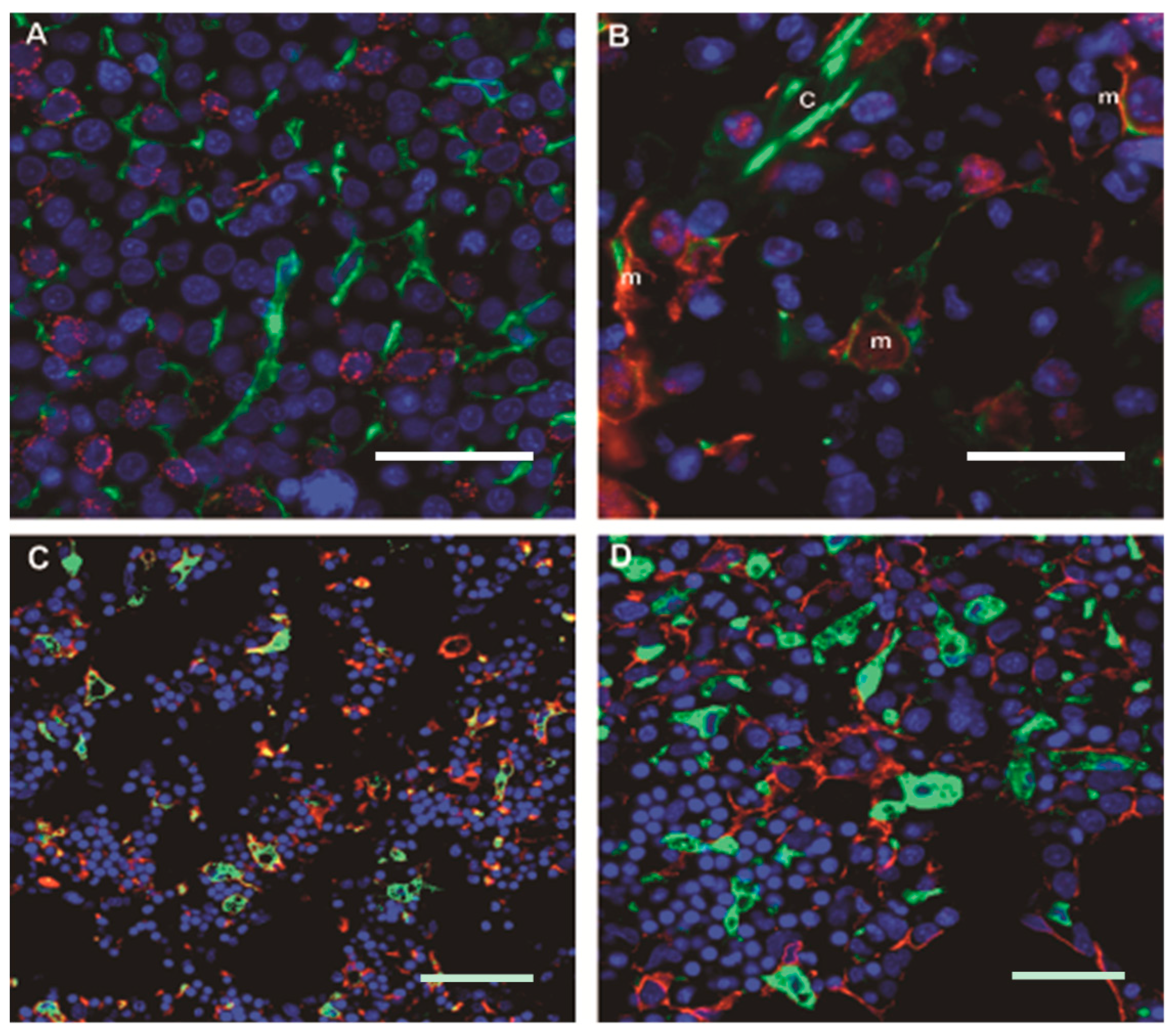
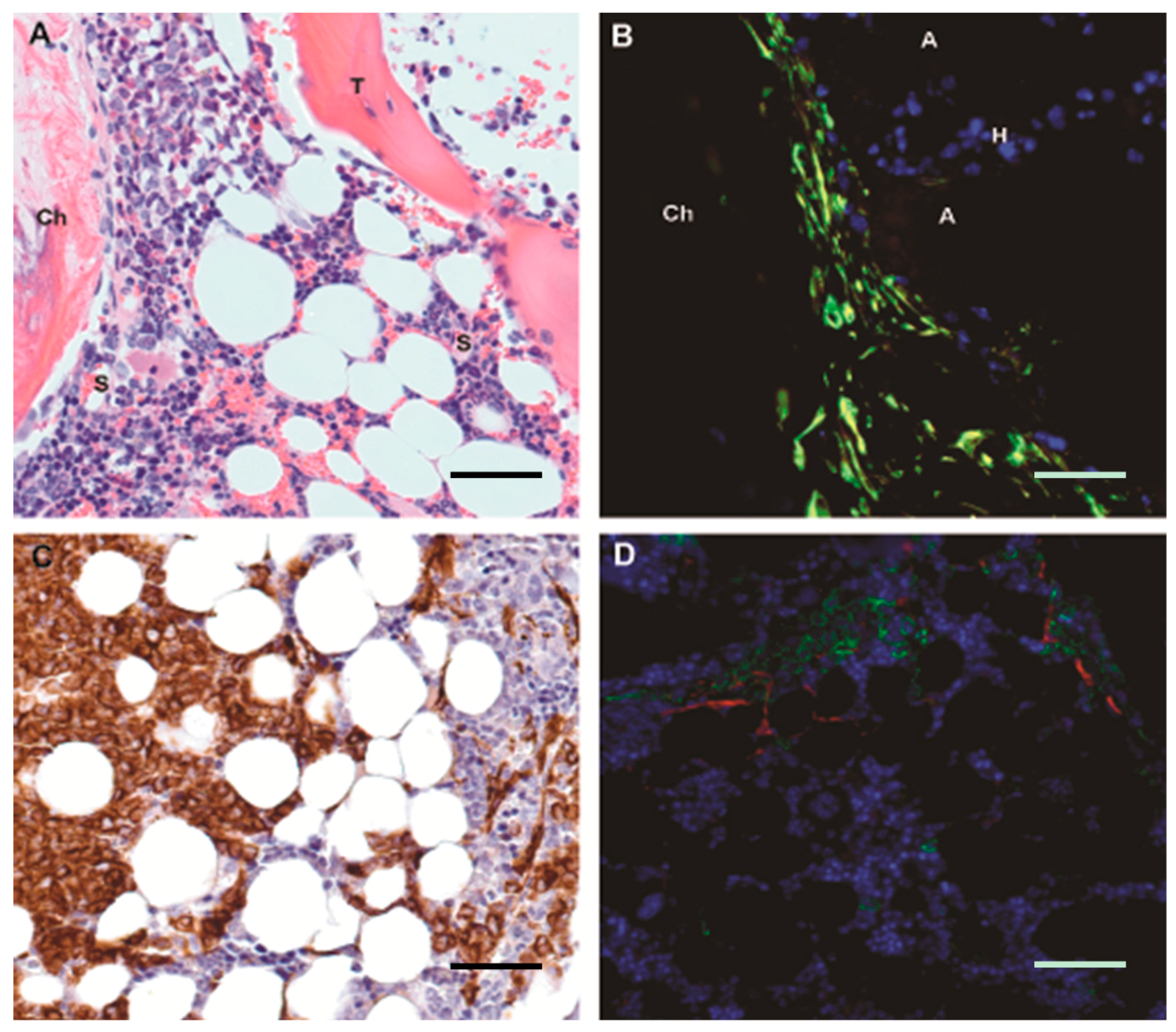
4. The Niche/Microenvironment in Humans
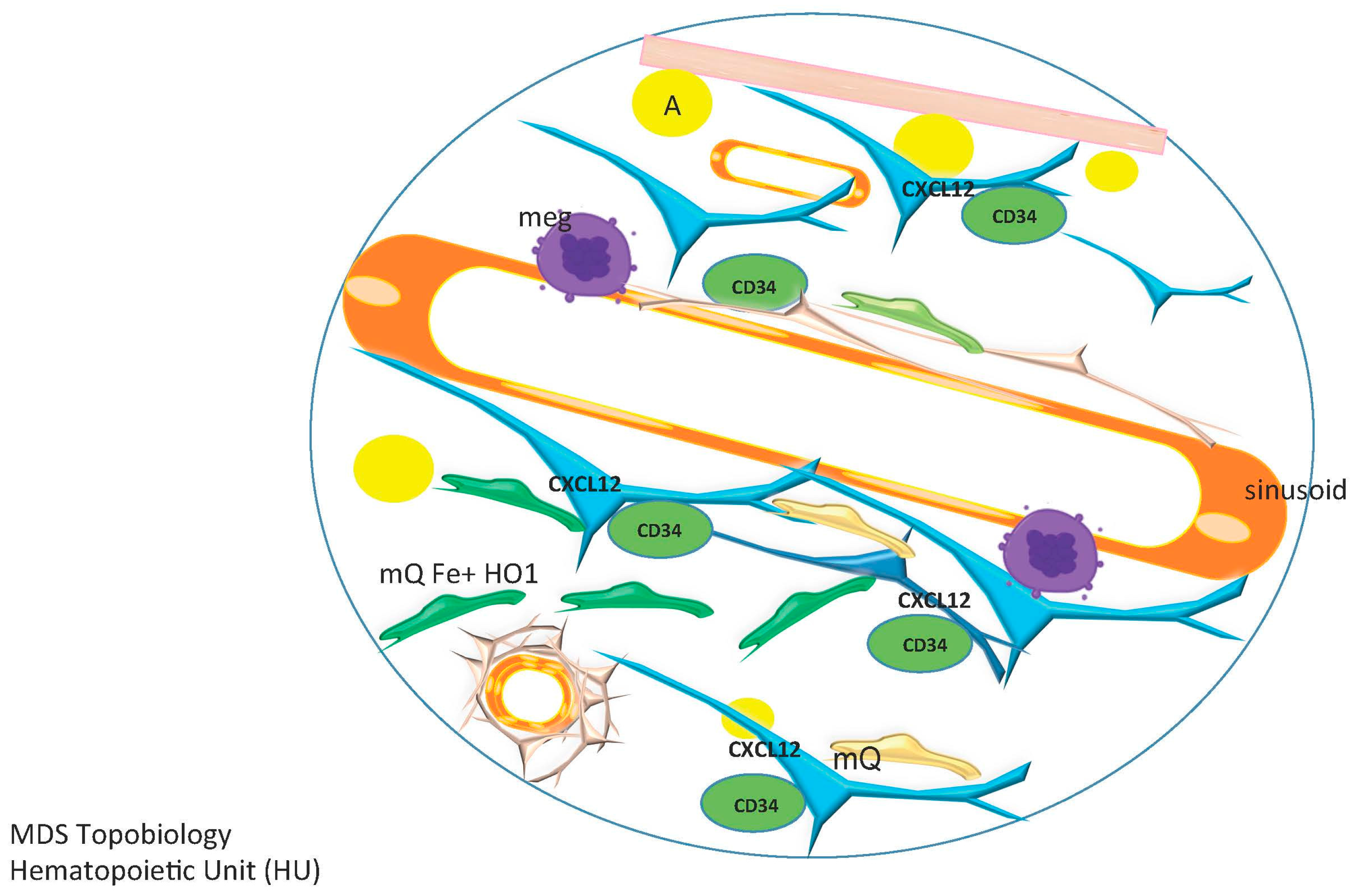
5. The Niche/Microenvironment in MDS
5.1. Mesenchymal Stromal Cells in MDS
5.2. Mouse Models to Study the Role of the Hematopoietic Niche/Bone Marrow Microenvironment in Hematopoiesis and MDS
6. Clinical Application of the Study of Niche/Microenvironment
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Edelman, G.M. Topobiology. Sci. Am. 1989, 260, 76–82, 84–86, 88. [Google Scholar] [CrossRef] [PubMed]
- Ma, X. Epidemiology of myelodysplastic syndromes. Am. J. Med. 2012, 125, S2–S5. [Google Scholar] [CrossRef] [PubMed]
- Brunning, R.D.; Orazi, A.; Germing, U.; LeBeau, M.M.; Porwit, A.; Baumann, I.; Vardiman, J.W.; Hellstrom-Lindberg, E. Myelodysplastic syndromes. In WHO Classification of Tumours of Hematopoietic and Lymphoid Tissues; World Health Organization Classification of Tumours, International Agency for Research on Cancer: Lyon, France, 2008; pp. 87–104. [Google Scholar]
- Gratzinger, D.; Greenberg, P.L. Update on Myelodysplastic Syndromes Classification and Prognosis. Surg. Pathol. Clin. 2013, 6, 693–728. [Google Scholar] [CrossRef] [PubMed]
- Cristy, M. Active bone marrow distribution as a function of age in humans. Phys. Med. Biol. 1981, 26, 389. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.A.; Callahan, J.; Bressel, M.; Simoens, N.; Everitt, S.; Hofman, M.S.; Hicks, R.J.; Burbury, K.; MacManus, M. Distribution Atlas of Proliferating Bone Marrow in Non-Small Cell Lung Cancer Patients Measured by FLT-PET/CT Imaging, With Potential Applicability in Radiation Therapy Planning. Int. J. Radiat. Oncol. Biol. Phys. 2015, 92, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Hartsock, R.J.; Smith, E.B.; Petty, C.S. Normal variations with aging of the amount of hematopoietic tissue in bone marrow from the anterior iliac crest. A study made from 177 cases of sudden death examined by necropsy. Am. J. Clin. Pathol. 1965, 43, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Rajaratnam, J.H.; Denton, J.; Hoyland, J.A.; Byers, R.J. Adipocytic proportion of bone marrow is inversely related to bone formation in osteoporosis. J. Clin. Pathol. 2002, 55, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, P.; Steiniche, T.; Vesterby, A.; Mosekilde, L.; Hessov, I.; Melsen, F. Primary hyperparathyroidism: Iliac crest trabecular bone volume, structure, remodeling, and balance evaluated by histomorphometric methods. Bone 1992, 13, 41–49. [Google Scholar] [CrossRef]
- Naito, K.; Tamahashi, N.; Chiba, T.; Kaneda, K.; Okuda, M.; Endo, K.; Yoshinaga, K.; Takahashi, T. The microvasculature of the human bone marrow correlated with the distribution of hematopoietic cells. A computer-assisted three-dimensional reconstruction study. Tohoku J. Exp. Med. 1992, 166, 439–450. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, P.P.; Breen, P.C.; Thomas, T.B. The microcirculation of the bone marrow. Anat. Rec. 1970, 168, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.P.; de Bruyn, P.P. The transmural passage of blood cells into myeloid sinusoids and the entry of platelets into the sinusoidal circulation; a scanning electron microscopic investigation. Am. J. Anat. 1976, 145, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Schaffler, M.B.; Wolde-Semait, H.T.; Hernandez, C.J.; Jepsen, K.J. Genetic background influences cortical bone response to ovariectomy. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2005, 20, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Büsche, G.; Ganser, A.; Li, Z. Morphology and quantitative composition of hematopoietic cells in murine bone marrow and spleen of healthy subjects. Ann. Hematol. 2013, 92, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, P.K.; Horowitz, M.C.; MacDougald, O.A.; Scheller, E.L.; Rodeheffer, M.S.; Rosen, C.J.; Klibanski, A. Marrow fat and bone—New perspectives. J. Clin. Endocrinol. Metab. 2013, 98, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Schofield, R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 1978, 4, 7–25. [Google Scholar] [PubMed]
- Grinnell, J. The Niche-Relationships of the California Thrasher. The Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. (Eds.) Ecological Niches. Linking Classical and Contemporary Approaches, 1st ed.; University of Chicago Press: Chicago, IL, USA, 2003.
- Xie, T.; Spradling, A.C. A niche maintaining germ line stem cells in the Drosophila ovary. Science 2000, 290, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Calvi, L.M.; Adams, G.B.; Weibrecht, K.W.; Weber, J.M.; Olson, D.P.; Knight, M.C.; Martin, R.P.; Schipani, E.; Divieti, P.; Bringhurst, F.R.; et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 2003, 425, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Niu, C.; Ye, L.; Huang, H.; He, X.; Tong, W.-G.; Ross, J.; Haug, J.; Johnson, T.; Feng, J.Q.; et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003, 425, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Visnjic, D.; Kalajzic, Z.; Rowe, D.W.; Katavic, V.; Lorenzo, J.; Aguila, H.L. Hematopoiesis is severely altered in mice with an induced osteoblast deficiency. Blood 2004, 103, 3258–3264. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Garrett, R.; Jung, Y.; Zhang, Y.; Kim, N.; Wang, J.; Joe, G.J.; Hexner, E.; Choi, Y.; Taichman, R.S.; et al. Osteoblasts support B-lymphocyte commitment and differentiation from hematopoietic stem cells. Blood 2007, 109, 3706–3712. [Google Scholar] [CrossRef] [PubMed]
- Acar, M.; Kocherlakota, K.S.; Murphy, M.M.; Peyer, J.G.; Oguro, H.; Inra, C.N.; Jaiyeola, C.; Zhao, Z.; Luby-Phelps, K.; Morrison, S.J. Deep imaging of bone marrow shows non-dividing stem cells are mainly perisinusoidal. Nature 2015, 526, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Coates, P. Bone turnover markers. Aust. Fam. Physician 2013, 42, 285–287. [Google Scholar] [PubMed]
- Teufel, S.; Grötsch, B.; Luther, J.; Derer, A.; Schinke, T.; Amling, M.; Schett, G.; Mielenz, D.; David, J.-P. Inhibition of bone remodeling in young mice by bisphosphonate displaces the plasma cell niche into the spleen. J. Immunol. 2014, 193, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Guezguez, B.; Campbell, C.J.V.; Boyd, A.L.; Karanu, F.; Casado, F.L.; di Cresce, C.; Collins, T.J.; Shapovalova, Z.; Xenocostas, A.; Bhatia, M. Regional localization within the bone marrow influences the functional capacity of human HSCs. Cell Stem Cell 2013, 13, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Morrison, S.J. Haematopoietic stem cells and early lymphoid progenitors occupy distinct bone marrow niches. Nature 2013, 495, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Gori, J.L.; Butler, J.M.; Chan, Y.-Y.; Chandrasekaran, D.; Poulos, M.G.; Ginsberg, M.; Nolan, D.J.; Elemento, O.; Wood, B.L.; Adair, J.E.; et al. Vascular niche promotes hematopoietic multipotent progenitor formation from pluripotent stem cells. J. Clin. Investig. 2015, 125, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, Y.; Sugiyama, T.; Kohara, H.; Kondoh, G.; Fujii, N.; Kohno, K.; Nagasawa, T. The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity 2010, 33, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; van Rooijen, N.; Alexander, K.A.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Huggins, M.; Ahmed, J.; Hashimoto, D.; Lucas, D.; Kunisaki, Y.; Pinho, S.; Leboeuf, M.; Noizat, C.; van Rooijen, N.; et al. CD169+ macrophages provide a niche promoting erythropoiesis under homeostasis and stress. Nat. Med. 2013, 19, 429–436. [Google Scholar] [CrossRef] [PubMed]
- McCabe, A.; MacNamara, K.C. Macrophages: Key regulators of steady-state and demand-adapted hematopoiesis. Exp. Hematol. 2016, 44, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Ferrer, S.; Lucas, D.; Battista, M.; Frenette, P.S. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 2008, 452, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Heazlewood, S.Y.; Neaves, R.J.; Williams, B.; Haylock, D.N.; Adams, T.E.; Nilsson, S.K. Megakaryocytes co-localise with hemopoietic stem cells and release cytokines that up-regulate stem cell proliferation. Stem Cell Res. 2013, 11, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Ishizu, A.; Takubo, K.; Fujioka, M.; Suda, T. Megakaryocytes are essential for HSC quiescence through the production of thrombopoietin. Biochem. Biophys. Res. Commun. 2014, 454, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Yoshida, S.; Kawasumi, M.; Hashimoto, K.; Kimura, T.; Sato, Y.; Kobayashi, T.; Miyauchi, Y.; Hoshi, H.; Iwasaki, R.; et al. Osteoclasts are dispensable for hematopoietic stem cell maintenance and mobilization. J. Exp. Med. 2011, 208, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.; Abou-Ezzi, G.; Sitnicka, E.; Jacobsen, S.E.W.; Wakkach, A.; Blin-Wakkach, C. Osteoclasts promote the formation of hematopoietic stem cell niches in the bone marrow. J. Exp. Med. 2012, 209, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Mayani, H.; Guilbert, L.J.; Janowska-Wieczorek, A. Biology of the hemopoietic microenvironment. Eur. J. Haematol. 1992, 49, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Noll, J.E.; Williams, S.A.; Purton, L.E.; Zannettino, A.C.W. Tug of war in the haematopoietic stem cell niche: Do myeloma plasma cells compete for the HSC niche? Blood Cancer J. 2012, 2, e91. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, A.; Hsu, Y.-M.S.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Nagasawa, T.; Link, D.C. CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 2013, 495, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.O.; Yue, R.; Murphy, M.M.; Peyer, J.G.; Morrison, S.J. Leptin-receptor-expressing mesenchymal stromal cells represent the main source of bone formed by adult bone marrow. Cell Stem Cell 2014, 15, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Orduña, G.R.; Mayani, H.; Montesinos, J.J. Hematopoietic support capacity of mesenchymal stem cells: biology and clinical potential. Arch. Med. Res. 2015, 46, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, S.; Saffrich, R.; Seckinger, A.; Krause, U.; Horsch, K.; Miesala, K.; Ho, A.D. Human mesenchymal stromal cells regulate initial self-renewing divisions of hematopoietic progenitor cells by a beta1-integrin-dependent mechanism. Stem Cells Dayt. Ohio 2007, 25, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Torlakovic, E.; Tenstad, E.; Funderud, S.; Rian, E. CD10+ stromal cells form B-lymphocyte maturation niches in the human bone marrow. J. Pathol. 2005, 205, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Cattoretti, G.; Schiro, R.; Orazi, A.; Soligo, D.; Colombo, M.P. Bone marrow stroma in humans: Anti-nerve growth factor receptor antibodies selectively stain reticular cells in vivo and in vitro. Blood 1993, 81, 1726–1738. [Google Scholar] [PubMed]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- Caneva, L.; Soligo, D.; Cattoretti, G.; de Harven, E.; Deliliers, G.L. Immuno-electron microscopy characterization of human bone marrow stromal cells with anti-NGFR antibodies. Blood Cells Mol. Dis. 1995, 21, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Tormin, A.; Li, O.; Brune, J.C.; Walsh, S.; Schütz, B.; Ehinger, M.; Ditzel, N.; Kassem, M.; Scheding, S. CD146 expression on primary nonhematopoietic bone marrow stem cells is correlated with in situ localization. Blood 2011, 117, 5067–5077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Figueroa, E.; Varma, S.; Montgomery, K.; Greenberg, P.L.; Gratzinger, D. Distinctive contact between CD34+ hematopoietic progenitors and CXCL12+ CD271+ mesenchymal stromal cells in benign and myelodysplastic bone marrow. Lab. Investig. J. Tech. Methods Pathol. 2012, 92, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Hooper, A.T.; Butler, J.M.; Nolan, D.J.; Kranz, A.; Iida, K.; Kobayashi, M.; Kopp, H.-G.; Shido, K.; Petit, I.; Yanger, K.; et al. Engraftment and reconstitution of hematopoiesis is dependent on VEGFR2-mediated regeneration of sinusoidal endothelial cells. Cell Stem Cell 2009, 4, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Ewalt, M.D.; Gratzinger, D. Selective Quantitation of Microvessel Density Reveals Sinusoidal Expansion in Myelodysplastic Syndromes. Leuk. Lymphoma 2016, in press. [Google Scholar]
- Sharma, M.; Afrin, F.; Satija, N.; Tripathi, R.P.; Gangenahalli, G.U. Stromal-derived factor-1/CXCR4 signaling: Indispensable role in homing and engraftment of hematopoietic stem cells in bone marrow. Stem Cells Dev. 2011, 20, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Lawen, A.; Lane, D.J.R. Mammalian iron homeostasis in health and disease: Uptake, storage, transport, and molecular mechanisms of action. Antioxid. Redox Signal. 2013, 18, 2473–2507. [Google Scholar] [CrossRef] [PubMed]
- Koorts, A.M.; Viljoen, M. Ferritin and ferritin isoforms I: Structure-function relationships, synthesis, degradation and secretion. Arch. Physiol. Biochem. 2007, 113, 30–54. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Lucas, D.; Hidalgo, A.; Mendez-Ferrer, S.; Hashimoto, D.; Scheiermann, C.; Battista, M.; Leboeuf, M.; Prophete, C.; van Rooijen, N.; et al. Bone marrow CD169+ macrophages promote the retention of hematopoietic stem and progenitor cells in the mesenchymal stem cell niche. J. Exp. Med. 2011, 208, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wegiel, B.; Hedblom, A.; Li, M.; Gallo, D.; Csizmadia, E.; Harris, C.; Nemeth, Z.; Zuckerbraun, B.S.; Soares, M.; Persson, J.L.; et al. Heme oxygenase-1 derived carbon monoxide permits maturation of myeloid cells. Cell Death Dis. 2014, 5, e1139. [Google Scholar] [CrossRef] [PubMed]
- Vanella, L.; Kim, D.H.; Asprinio, D.; Peterson, S.J.; Barbagallo, I.; Vanella, A.; Goldstein, D.; Ikehara, S.; Kappas, A.; Abraham, N.G. HO-1 expression increases mesenchymal stem cell-derived osteoblasts but decreases adipocyte lineage. Bone 2010, 46, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, G.; Gratzinger, D. Myelodysplastic syndrome macrophages have aberrant iron storage and heme oxygenase-1 expression. Leuk. Lymphoma 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; di Giuseppe, M.; Njah, J.; Sala, E.; Shiva, S.; St Croix, C.M.; Stolz, D.B.; Watkins, S.C.; Di, Y.P.; Leikauf, G.D.; et al. Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 2015, 6, 8472. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, B.; Funari, A.; Michienzi, S.; di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Maijenburg, M.W.; Kleijer, M.; Vermeul, K.; Mul, E.; van Alphen, F.; van der Schoot, C.E.; Voermans, C. The composition of the mesenchymal stromal cell compartment in human bone marrow changes during development and aging. Haematologica 2012, 97, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Hanoun, M.; Maryanovich, M.; Arnal-Estapé, A.; Frenette, P.S. Neural regulation of hematopoiesis, inflammation, and cancer. Neuron 2015, 86, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Ema, H.; Karlsson, G.; Yamaguchi, T.; Miyoshi, H.; Shioda, S.; Taketo, M.M.; Karlsson, S.; Iwama, A.; Nakauchi, H. Nonmyelinating schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 2011, 147, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Allen, T.D. Ultrastructural morphometric study of efferent nerve terminals on murine bone marrow stromal cells, and the recognition of a novel anatomical unit: The “neuro-reticular complex”. Am. J. Anat. 1990, 187, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Vasikova, A.; Belickova, M.; Budinska, E.; Cermak, J. A distinct expression of various gene subsets in CD34+ cells from patients with early and advanced myelodysplastic syndrome. Leuk. Res. 2010, 34, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Maftoun-Banankhah, S.; Maleki, A.; Karandikar, N.J.; Arbini, A.A.; Fuda, F.S.; Wang, H.-Y.; Chen, W. Multiparameter flow cytometric analysis reveals low percentage of bone marrow hematogones in myelodysplastic syndromes. Am. J. Clin. Pathol. 2008, 129, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Day, R.B.; Bhattacharya, D.; Nagasawa, T.; Link, D.C. Granulocyte colony-stimulating factor reprograms bone marrow stromal cells to actively suppress B lymphopoiesis in mice. Blood 2015, 125, 3114–3117. [Google Scholar] [CrossRef] [PubMed]
- Borbényi, Z.; Tetla, N.G.; Varga, G. Granulocytic progenitor cells in the adherent layer of human long-term bone marrow cultures. Haematologia (Budap.) 1988, 21, 115–120. [Google Scholar] [PubMed]
- Wetzler, M.; Kurzrock, R.; Estrov, Z.; Estey, E.; Talpaz, M. Cytokine expression in adherent layers from patients with myelodysplastic syndrome and acute myelogenous leukemia. Leuk. Res. 1995, 19, 23–34. [Google Scholar] [CrossRef]
- Flores-Figueroa, E.; Gutierrez-Espindola, G.; Guerrero-Rivera, S.; Pizzuto-Chavez, J.; Mayani, H. Hematopoietic progenitor cells from patients with myelodysplastic syndromes: In vitro colony growth and long-term proliferation. Leuk. Res. 1999, 23, 385–394. [Google Scholar] [CrossRef]
- Flores-Figueroa, E.; Gutierrez-Espindola, G.; Montesinos, J.J.; Arana-Trejo, R.M.; Mayani, H. In vitro characterization of hematopoietic microenvironment cells from patients with myelodysplastic syndrome. Leuk. Res. 2002, 26, 677–686. [Google Scholar] [CrossRef]
- Alvi, S.; Shaher, A.; Shetty, V.; Henderson, B.; Dangerfield, B.; Zorat, F.; Joshi, L.; Anthwal, S.; Lisak, L.; Little, L.; et al. Successful establishment of long-term bone marrow cultures in 103 patients with myelodysplastic syndromes. Leuk. Res. 2001, 25, 941–954. [Google Scholar] [CrossRef]
- Coutinho, L.H.; Geary, C.G.; Chang, J.; Harrison, C.; Testa, N.G. Functional studies of bone marrow haemopoietic and stromal cells in the myelodysplastic syndrome (MDS). Br. J. Haematol. 1990, 75, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, S.; Nakano, M.; Iwase, O.; Yaguchi, M.; Hiramoto, M.; Hoshi, H.; Nabeshima, R.; Shima, D.; Handa, H.; Toyama, K. Bone marrow stroma from refractory anemia of myelodysplastic syndrome is defective in its ability to support normal CD34-positive cell proliferation and differentiation in vitro. Leuk. Res. 1999, 23, 239–246. [Google Scholar] [CrossRef]
- Mundle, S.; Iftikhar, A.; Shetty, V.; Dameron, S.; Wright-Quinones, V.; Marcus, B.; Loew, J.; Gregory, S.; Raza, A. Novel in situ double labeling for simultaneous detection of proliferation and apoptosis. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1994, 42, 1533–1537. [Google Scholar] [CrossRef]
- Kitagawa, M.; Saito, I.; Kuwata, T.; Yoshida, S.; Yamaguchi, S.; Takahashi, M.; Tanizawa, T.; Kamiyama, R.; Hirokawa, K. Overexpression of tumor necrosis factor (TNF)-alpha and interferon (IFN)-gamma by bone marrow cells from patients with myelodysplastic syndromes. Leuk. Off. J. Leuk. Soc. Am. Leuk. Res. Fund UK 1997, 11, 2049–2054. [Google Scholar] [CrossRef]
- Boudard, D.; Viallet, A.; Piselli, S.; Guyotat, D.; Campos, L. In vitro study of stromal cell defects in myelodysplastic syndromes. Haematologica 2003, 88, 827–829. [Google Scholar] [PubMed]
- Shetty, V.; Mundle, S.; Alvi, S.; Showel, M.; Broady-Robinson, L.; Dar, S.; Borok, R.; Showel, J.; Gregory, S.; Rifkin, S.; et al. Measurement of apoptosis, proliferation and three cytokines in 46 patients with myelodysplastic syndromes. Leuk. Res. 1996, 20, 891–900. [Google Scholar] [CrossRef]
- Ohmori, S.; Ohmori, M.; Yamagishi, M.; Okuma, M. MDS-macrophage derived inhibitory activity on myelopoiesis of MDS abnormal clones. Br. J. Haematol. 1993, 83, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Allampallam, K.; Shetty, V.; Mundle, S.; Dutt, D.; Kravitz, H.; Reddy, P.L.; Alvi, S.; Galili, N.; Saberwal, G.S.; Anthwal, S.; et al. Biological significance of proliferation, apoptosis, cytokines, and monocyte/macrophage cells in bone marrow biopsies of 145 patients with myelodysplastic syndrome. Int. J. Hematol. 2002, 75, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, P.L.; Tuechler, H.; Schanz, J.; Sanz, G.; Garcia-Manero, G.; Solé, F.; Bennett, J.M.; Bowen, D.; Fenaux, P.; Dreyfus, F.; et al. Revised International Prognostic Scoring System (IPSS-R) for myelodysplastic syndromes. Blood 2012, 120, 2454–2465. [Google Scholar] [CrossRef] [PubMed]
- Kastrinaki, M.-C.; Pavlaki, K.; Batsali, A.K.; Kouvidi, E.; Mavroudi, I.; Pontikoglou, C.; Papadaki, H.A. Mesenchymal stem cells in immune-mediated bone marrow failure syndromes. Clin. Dev. Immunol. 2013, 2013, 265608. [Google Scholar] [CrossRef] [PubMed]
- Cogle, C.R.; Saki, N.; Khodadi, E.; Li, J.; Shahjahani, M.; Azizidoost, S. Bone marrow niche in the myelodysplastic syndromes. Leuk. Res. 2015, 39, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Bulycheva, E.; Rauner, M.; Medyouf, H.; Theurl, I.; Bornhäuser, M.; Hofbauer, L.C.; Platzbecker, U. Myelodysplasia is in the niche: Novel concepts and emerging therapies. Leukemia 2015, 29, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Mies, A.; Bulycheva, E.; Rogulj, I.M.; Hofbauer, L.C.; Platzbecker, U. Alterations within the Osteo-Hematopoietic Niche in MDS and their Therapeutic Implications. Curr. Pharm. Des. 2016, in press. [Google Scholar] [CrossRef]
- Flores-Figueroa, E.; Arana-Trejo, R.M.; Gutierrez-Espindola, G.; Perez-Cabrera, A.; Mayani, H. Mesenchymal stem cells in myelodysplastic syndromes: Phenotypic and cytogenetic characterization. Leuk. Res. 2005, 29, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Blau, O.; Hofmann, W.K.; Baldus, C.D.; Thiel, G.; Serbent, V.; Schumann, E.; Thiel, E.; Blau, I.W. Chromosomal aberrations in bone marrow mesenchymal stroma cells from patients with myelodysplastic syndrome and acute myeloblastic leukemia. Exp. Hematol. 2007, 35, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Villar, O.; Garcia, J.L.; Sanchez-Guijo, F.M.; Robledo, C.; Villaron, E.M.; Hernandez-Campo, P.; Lopez-Holgado, N.; Diez-Campelo, M.; Barbado, M.V.; Perez-Simon, J.A.; et al. Both expanded and uncultured mesenchymal stem cells from MDS patients are genomically abnormal, showing a specific genetic profile for the 5q- syndrome. Leuk. Off. J. Leuk. Soc. Am. Leuk. Res. Fund UK 2009, 23, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Soenen-Cornu, V.; Tourino, C.; Bonnet, M.-L.; Guillier, M.; Flamant, S.; Kotb, R.; Bernheim, A.; Bourhis, J.-H.; Preudhomme, C.; Fenaux, P.; et al. Mesenchymal cells generated from patients with myelodysplastic syndromes are devoid of chromosomal clonal markers and support short- and long-term hematopoiesis in vitro. Oncogene 2005, 24, 2441–2448. [Google Scholar] [CrossRef] [PubMed]
- Varga, G.; Kiss, J.; Várkonyi, J.; Vas, V.; Farkas, P.; Pálóczi, K.; Uher, F. Inappropriate Notch activity and limited mesenchymal stem cell plasticity in the bone marrow of patients with myelodysplastic syndromes. Pathol. Oncol. Res. POR 2007, 13, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Geyh, S.; Oz, S.; Cadeddu, R.-P.; Fröbel, J.; Brückner, B.; Kündgen, A.; Fenk, R.; Bruns, I.; Zilkens, C.; Hermsen, D.; et al. Insufficient stromal support in MDS results from molecular and functional deficits of mesenchymal stromal cells. Leukemia 2013, 27, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Kurzer, J.H.; Greenberg, P.L.; Gratzinger, D. Mesenchymal stromal cell density is increased in higher grade myelodysplastic syndromes and independently predicts survival. Am. J. Clin. Pathol. 2014, 142, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Abe-Suzuki, S.; Kurata, M.; Abe, S.; Onishi, I.; Kirimura, S.; Nashimoto, M.; Murayama, T.; Hidaka, M.; Kitagawa, M. CXCL12+ stromal cells as bone marrow niche for CD34+ hematopoietic cells and their association with disease progression in myelodysplastic syndromes. Lab. Investig. J. Tech. Methods Pathol. 2014, 94, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Fei, C.; Zhao, Y.; Gu, S.; Guo, J.; Zhang, X.; Li, X.; Chang, C. Impaired osteogenic differentiation of mesenchymal stem cells derived from bone marrow of patients with lower-risk myelodysplastic syndromes. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 4307–4316. [Google Scholar] [CrossRef] [PubMed]
- Pavlaki, K.; Pontikoglou, C.G.; Demetriadou, A.; Batsali, A.K.; Damianaki, A.; Simantirakis, E.; Kontakis, M.; Galanopoulos, A.; Kotsianidis, I.; Kastrinaki, M.-C.; et al. Impaired proliferative potential of bone marrow mesenchymal stromal cells in patients with myelodysplastic syndromes is associated with abnormal WNT signaling pathway. Stem Cells Dev. 2014, 23, 1568–1581. [Google Scholar] [CrossRef] [PubMed]
- Fei, C.; Zhao, Y.; Guo, J.; Gu, S.; Li, X.; Chang, C. Senescence of bone marrow mesenchymal stromal cells is accompanied by activation of p53/p21 pathway in myelodysplastic syndromes. Eur. J. Haematol. 2014, 93, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, D.; Fei, C.; Guo, J.; Gu, S.; Zhu, Y.; Xu, F.; Zhang, Z.; Wu, L.; Li, X.; et al. Down-regulation of Dicer1 promotes cellular senescence and decreases the differentiation and stem cell-supporting capacities of mesenchymal stromal cells in patients with myelodysplastic syndrome. Haematologica 2015, 100, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Mellibovsky, L.; Diez, A.; Serrano, S.; Aubia, J.; Pérez-Vila, E.; Mariñoso, M.L.; Nogués, X.; Recker, R.R. Bone remodeling alterations in myelodysplastic syndrome. Bone 1996, 19, 401–405. [Google Scholar] [CrossRef]
- Flores-Figueroa, E.; Montesinos, J.J.; Flores-Guzman, P.; Gutierrez-Espindola, G.; Arana-Trejo, R.M.; Castillo-Medina, S.; Perez-Cabrera, A.; Hernandez-Estevez, E.; Arriaga, L.; Mayani, H. Functional analysis of myelodysplastic syndromes-derived mesenchymal stem cells. Leuk. Res. 2008, 32, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, Z.; Li, Q.; Li, W.; You, Y.; Zou, P. The different immunoregulatory functions of mesenchymal stem cells in patients with low-risk or high-risk myelodysplastic syndromes. PLoS ONE 2012, 7, e45675. [Google Scholar] [CrossRef] [PubMed]
- Klaus, M.; Stavroulaki, E.; Kastrinaki, M.-C.; Fragioudaki, P.; Giannikou, K.; Psyllaki, M.; Pontikoglou, C.; Tsoukatou, D.; Mamalaki, C.; Papadaki, H.A. Reserves, functional, immunoregulatory, and cytogenetic properties of bone marrow mesenchymal stem cells in patients with myelodysplastic syndromes. Stem Cells Dev. 2010, 19, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, R.A.; Wobus, M.; List, C.; Wehner, R.; Schönefeldt, C.; Brocard, B.; Mohr, B.; Rauner, M.; Schmitz, M.; Stiehler, M.; et al. Mesenchymal stromal cells from patients with myelodyplastic syndrome display distinct functional alterations that are modulated by lenalidomide. Haematologica 2013, 98, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Castro-Manrreza, M.E.; Montesinos, J.J. Immunoregulation by mesenchymal stem cells: Biological aspects and clinical applications. J. Immunol. Res. 2015, 2015, 394917. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, X.; Xu, W.; Cao, Z.; Sun, L.; Li, W.; Li, Q.; Zou, P.; Zhao, Z. The different immunoregulatory functions on dendritic cells between mesenchymal stem cells derived from bone marrow of patients with low-risk or high-risk myelodysplastic syndromes. PLoS ONE 2013, 8, e57470. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Dexter, T.M.; Wright, E.G.; Krizsa, F.; Lajtha, L.G. Regulation of haemopoietic stem cell proliferation in long term bone marrow cultures. Biomed. Publiée Pour AAICIG 1977, 27, 344–349. [Google Scholar]
- Dexter, T.M.; Moore, M.A.; Sheridan, A.P. Maintenance of hemopoietic stem cells and production of differentiated progeny in allogeneic and semiallogeneic bone marrow chimeras in vitro. J. Exp. Med. 1977, 145, 1612–1616. [Google Scholar] [CrossRef] [PubMed]
- Walkley, C.R.; Olsen, G.H.; Dworkin, S.; Fabb, S.A.; Swann, J.; McArthur, G.A.; Westmoreland, S.V.; Chambon, P.; Scadden, D.T.; Purton, L.E. A microenvironment-induced myeloproliferative syndrome caused by retinoic acid receptor gamma deficiency. Cell 2007, 129, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Walkley, C.R.; Shea, J.M.; Sims, N.A.; Purton, L.E.; Orkin, S.H. Rb regulates interactions between hematopoietic stem cells and their bone marrow microenvironment. Cell 2007, 129, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Neaves, W.B. Normal stem cells and cancer stem cells: The niche matters. Cancer Res. 2006, 66, 4553–4557. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, M.H.; Mukherjee, S.; Guo, S.; Zhang, S.; Kobayashi, T.; Schoonmaker, J.A.; Ebert, B.L.; Al-Shahrour, F.; Hasserjian, R.P.; Scadden, E.O.; et al. Bone progenitor dysfunction induces myelodysplasia and secondary leukaemia. Nature 2010, 464, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Kinney, M.C.; Scott, L.M.; Zinkel, S.S.; Rebel, V.I. Revisiting the case for genetically engineered mouse models in human myelodysplastic syndrome research. Blood 2015, 126, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Muguruma, Y.; Matsushita, H.; Yahata, T.; Yumino, S.; Tanaka, Y.; Miyachi, H.; Ogawa, Y.; Kawada, H.; Ito, M.; Ando, K. Establishment of a xenograft model of human myelodysplastic syndromes. Haematologica 2011, 96, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Medyouf, H.; Mossner, M.; Jann, J.-C.; Nolte, F.; Raffel, S.; Herrmann, C.; Lier, A.; Eisen, C.; Nowak, V.; Zens, B.; et al. Myelodysplastic cells in patients reprogram mesenchymal stromal cells to establish a transplantable stem cell niche disease unit. Cell Stem Cell 2014, 14, 824–837. [Google Scholar] [PubMed]
- Chen, Y.; Jacamo, R.; Shi, Y.; Wang, R.; Battula, V.L.; Konoplev, S.; Strunk, D.; Hofmann, N.A.; Reinisch, A.; Konopleva, M.; et al. Human extramedullary bone marrow in mice: A novel in vivo model of genetically controlled hematopoietic microenvironment. Blood 2012, 119, 4971–4980. [Google Scholar] [CrossRef] [PubMed]
- Reinisch, A.; Etchart, N.; Thomas, D.; Hofmann, N.A.; Fruehwirth, M.; Sinha, S.; Chan, C.K.; Senarath-Yapa, K.; Seo, E.-Y.; Wearda, T.; et al. Epigenetic and in vivo comparison of diverse MSC sources reveals an endochondral signature for human hematopoietic niche formation. Blood 2015, 125, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Reinisch, A.; Thomas, D.; Corces, R.M.; Zhang, X.; Gratzinger, D.; Hong, W.-J.; Schallmoser, K.; Strunk, D.; Majeti, R. A Humanized ossicle niche Xenotransplantation model with improved hematopoietic engraftment identifies Leukemia-initiating cells in APL and myelofibrosis. Nat. Med. 2016, in press. [Google Scholar]
- Vardiman, J.W.; Thiele, J.; Arber, D.A.; Brunning, R.D.; Borowitz, M.J.; Porwit, A.; Harris, N.L.; Le Beau, M.M.; Hellström-Lindberg, E.; Tefferi, A.; et al. The 2008 revision of the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia: Rationale and important changes. Blood 2009, 114, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Tothova, Z.; Steensma, D.P.; Ebert, B.L. New strategies in myelodysplastic syndromes: Application of molecular diagnostics to clinical practice. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Genovese, G.; Kähler, A.K.; Handsaker, R.E.; Lindberg, J.; Rose, S.A.; Bakhoum, S.F.; Chambert, K.; Mick, E.; Neale, B.M.; Fromer, M.; et al. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N. Engl. J. Med. 2014, 371, 2477–2487. [Google Scholar] [CrossRef] [PubMed]
- Kwok, B.; Hall, J.M.; Witte, J.S.; Xu, Y.; Reddy, P.; Lin, K.; Flamholz, R.; Dabbas, B.; Yung, A.; Al-Hafidh, J.; et al. MDS-associated somatic mutations and clonal hematopoiesis are common in idiopathic cytopenias of undetermined significance. Blood 2015, 126, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- Mahaira, L.G.; Katsara, O.; Pappou, E.; Iliopoulou, E.G.; Fortis, S.; Antsaklis, A.; Fotinopoulos, P.; Baxevanis, C.N.; Papamichail, M.; Perez, S.A. IGF2BP1 expression in human mesenchymal stem cells significantly affects their proliferation and is under the epigenetic control of TET1/2 demethylases. Stem Cells Dev. 2014, 23, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, N.; Zhang, L.-N.; Huang, N.; Song, T.-F.; Li, Z.-Z.; Li, M.; Luo, X.-G.; Zhou, H.; He, H.-P.; et al. Knockdown of Dnmt1 and Dnmt3a Promotes the Angiogenesis of hMSCs Leading to Arterial Specific Differentiation. Stem Cells Dayt. Ohio 2016. [Google Scholar] [CrossRef] [PubMed]
- Balderman, S.R.; Li, A.J.; Hoffman, C.M.; Frisch, B.J.; Goodman, A.N.; LaMere, M.W.; Georger, M.A.; Evans, A.G.; Liesveld, J.L.; Becker, M.W.; et al. Targeting of the bone marrow microenvironment improves outcome in a murine model of myelodysplastic syndrome. Blood 2016, 127, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montero, A.C.; Jara-Acevedo, M.; Alvarez-Twose, I.; Teodosio, C.; Sanchez-Muñoz, L.; Muñiz, C.; Muñoz-Gonzalez, J.I.; Mayado, A.; Matito, A.; Caldas, C.; et al. KIT D816V-mutated bone marrow mesenchymal stem cells in indolent systemic mastocytosis are associated with disease progression. Blood 2016, 127, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Muntión, S.; Ramos, T.L.; Diez-Campelo, M.; Rosón, B.; Sánchez-Abarca, L.I.; Misiewicz-Krzeminska, I.; Preciado, S.; Sarasquete, M.-E.; de Las Rivas, J.; González, M.; et al. Microvesicles from Mesenchymal Stromal Cells Are Involved in HPC-Microenvironment Crosstalk in Myelodysplastic Patients. PLoS ONE 2016, 11, e0146722. [Google Scholar] [CrossRef] [PubMed]
- Della Porta, M.G.; Malcovati, L. Myelodysplastic syndromes with bone marrow fibrosis. Haematologica 2011, 96, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.L.; Chen, E.; Sasane, M.; Paley, C.; Guo, A.; Laouri, M. Economic impact on US Medicare of a new diagnosis of myelodysplastic syndromes and the incremental costs associated with blood transfusion need. Transfusion (Paris) 2012, 52, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Gattermann, N.; Finelli, C.; Della Porta, M.; Fenaux, P.; Stadler, M.; Guerci-Bresler, A.; Schmid, M.; Taylor, K.; Vassilieff, D.; Habr, D.; et al. Hematologic responses to deferasirox therapy in transfusion-dependent patients with myelodysplastic syndromes. Haematologica 2012, 97, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zhao, M.; Rajbhandary, S.; Xie, F.; Chai, X.; Mu, J.; Meng, J.; Liu, Y.; Jiang, Y.; Xu, X.; et al. Free iron catalyzes oxidative damage to hematopoietic cells/mesenchymal stem cells in vitro and suppresses hematopoiesis in iron overload patients. Eur. J. Haematol. 2013, 91, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Charbord, P. Stromal Support of Hematopoiesis. In Stem Cells Handbook; MD, S.S., Ed.; Humana Press: New York City, NY, USA, 2004; pp. 143–154. [Google Scholar]
- Wolf, N.S.; Trentin, J.J. The restorative effect of erythropoietic stimulation upon the sublethally irradiated (SLI) hematopoietic stem cell and/or its progeny. Exp. Hematol. 1975, 3, 57–64. [Google Scholar] [PubMed]
- Till, J.E.; Mcculloch, E.A.; Siminovitch, L. A stochastic model of stem cell proliferation, BASED ON THE GROWTH OF SPLEEN COLONY-FORMING CELLS. Proc. Natl. Acad. Sci. USA 1964, 51, 29–36. [Google Scholar] [CrossRef] [PubMed]
- La Pushin, R.W.; Trentin, J.J. Identification of distinctive stromal elements in erythroid and neutrophil granuloid spleen colonies: Light and electron microscopic study. Exp. Hematol. 1977, 5, 505–522. [Google Scholar] [PubMed]
- Dexter, T.M.; Allen, T.D.; Lajtha, L.G. Conditions controlling the proliferation of haemopoietic stem cells in vitro. J. Cell. Physiol. 1977, 91, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Edelman, G.M. Topobiology: An Introduction to Molecular Embryology; Basic Books: New York, NY, USA, 1988. [Google Scholar]
- Naeim, F.; Moatamed, F.; Sahimi, M. Morphogenesis of the bone marrow: Fractal structures and diffusion-limited growth. Blood 1996, 87, 5027–5031. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Stromal Cultures | MSC | Macrophages |
|---|---|---|---|
| TNFα | increased | normal | increased |
| IL-1B | increased | increased | – |
| IL-6 | increased | normal | – |
| IFNγ | increased | increased | increased-subset |
| TGFβ | increased | increased (HR) | correlate with number |
| SCF | – | increased (2) reduced (2) | – |
| CXCL12/SDF1 | – | increased (2) reduced (3) | – |
| Hematopoietic support | normal (1) deficient (1) | normal (2) deficient (6) | – |
| Proliferation | normal | normal (5) deficient (7) | NA |
| CFU-F | NA | normal (1) deficient (3) | NA |
| Karyotype | – | abnormal | – |
| Osteoblasts | NA | normal (5) abnormal (9) | NA |
| Adipocytes | NA | normal (4) abnormal (3) higher (1) | NA |
| Immunomodulation | NA | normal (1) abnormal (3) | NA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Figueroa, E.; Gratzinger, D. Beyond the Niche: Myelodysplastic Syndrome Topobiology in the Laboratory and in the Clinic. Int. J. Mol. Sci. 2016, 17, 553. https://doi.org/10.3390/ijms17040553
Flores-Figueroa E, Gratzinger D. Beyond the Niche: Myelodysplastic Syndrome Topobiology in the Laboratory and in the Clinic. International Journal of Molecular Sciences. 2016; 17(4):553. https://doi.org/10.3390/ijms17040553
Chicago/Turabian StyleFlores-Figueroa, Eugenia, and Dita Gratzinger. 2016. "Beyond the Niche: Myelodysplastic Syndrome Topobiology in the Laboratory and in the Clinic" International Journal of Molecular Sciences 17, no. 4: 553. https://doi.org/10.3390/ijms17040553
APA StyleFlores-Figueroa, E., & Gratzinger, D. (2016). Beyond the Niche: Myelodysplastic Syndrome Topobiology in the Laboratory and in the Clinic. International Journal of Molecular Sciences, 17(4), 553. https://doi.org/10.3390/ijms17040553