
Goldfish Leptin-AI and Leptin-AII: Function and Central Mechanism in Feeding Control

 ,
,
Abstract
:
1. Introduction
2. Results
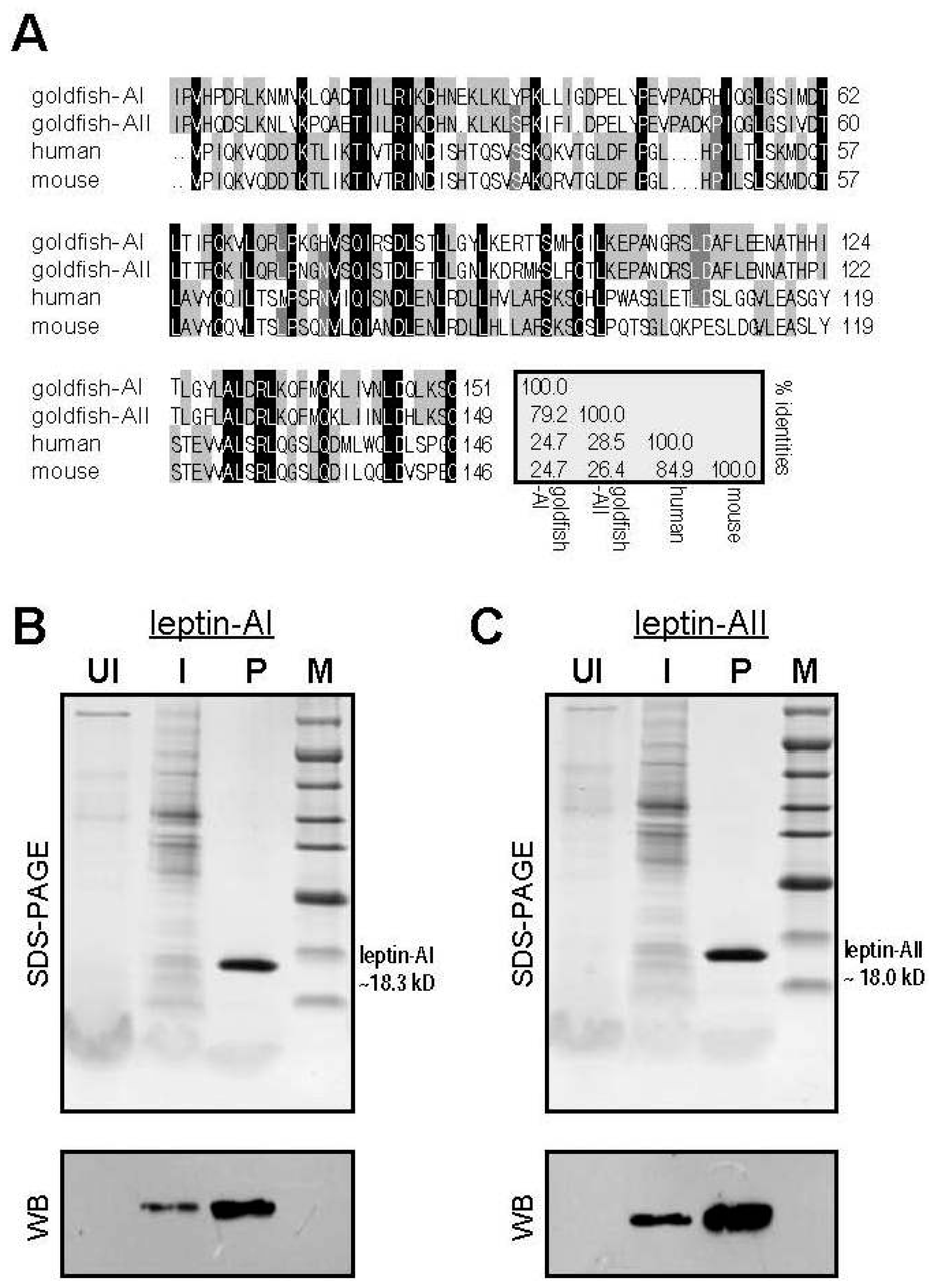
2.1. Alignment of Goldfish and Mammalian Leptin Mature Peptide Sequences
2.2. Expression and Purification of Goldfish Leptin-AI and Leptin-AII Protein
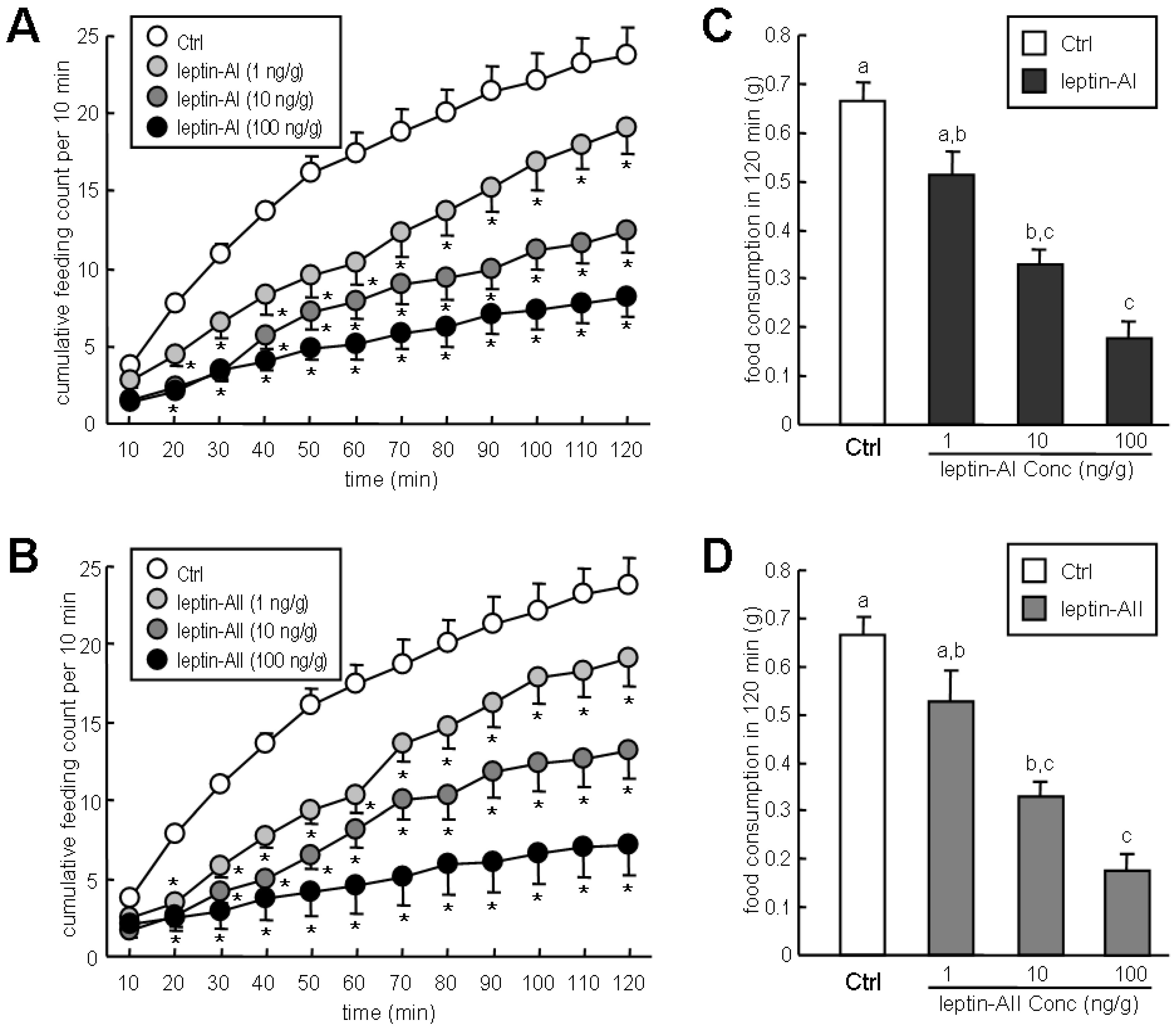
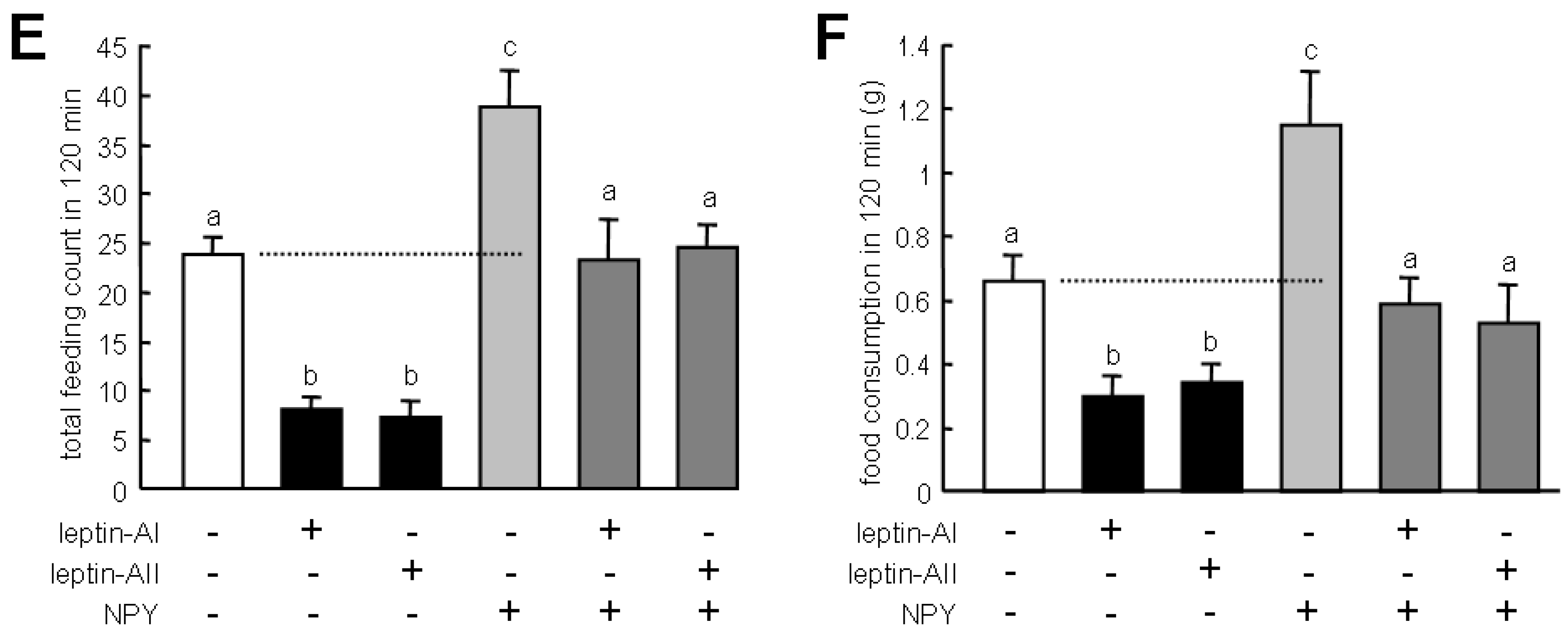
2.3. Effects of Leptin-AI and Leptin-AII on Goldfish Feeding Behavior and Food Consumption
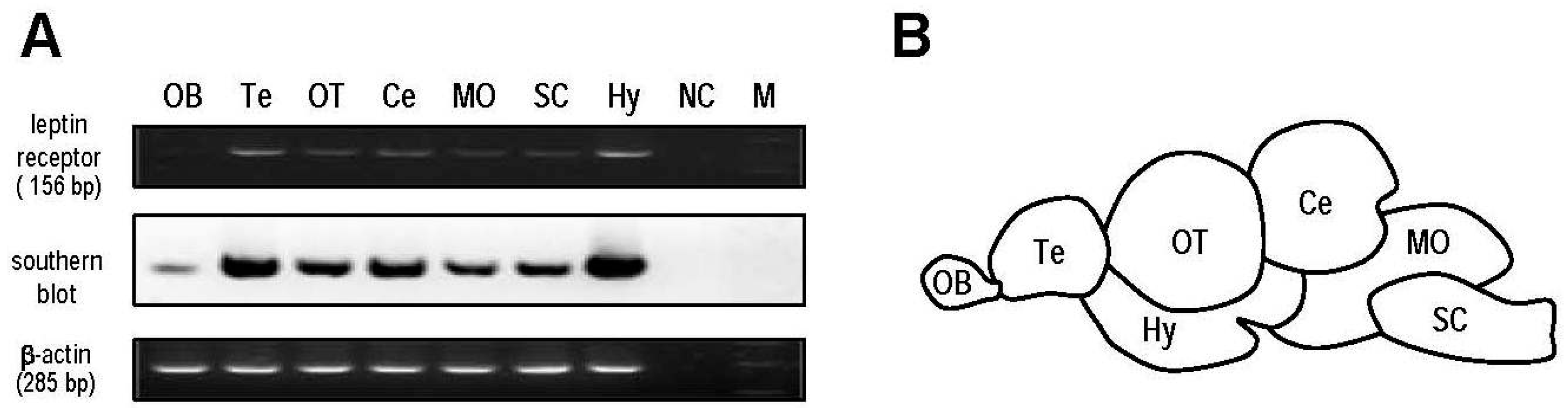
2.4. Expression Profile of Leptin Receptor in Different Brain Regions
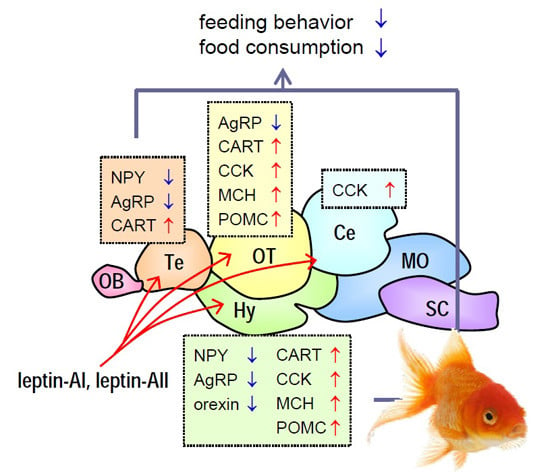
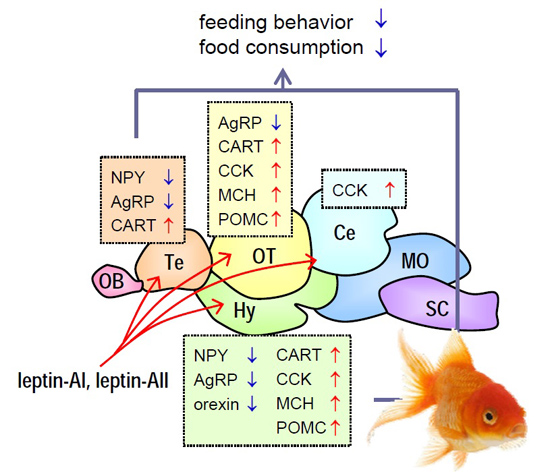
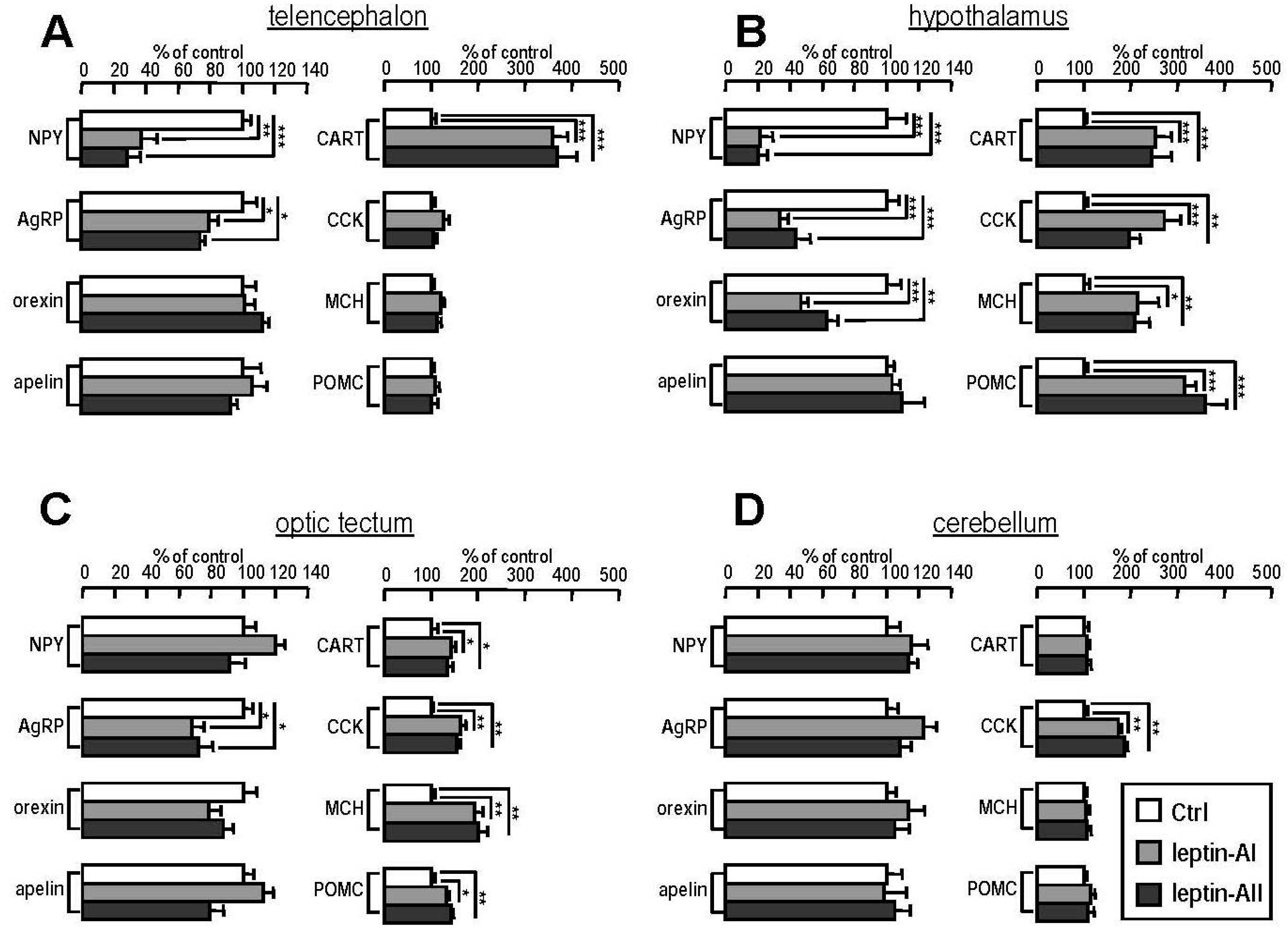
2.5. Transcript Expression of Appetite-Related Hormones in Selected Brain Regions after Leptin-AI and Leptin-AII Injection
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sequence Alignment and Identities Analysis
4.3. Production of Goldfish Leptin-AI and Leptin-AII Recombinant Protein in P. pastoris
4.4. Effect of Leptin-AI and Leptin-AII on Goldfish Feeding Behavior and Food Consumption
4.5. Semi-Quantitative Analysis of Leptin Receptor Expression in Different Brain Regions
4.6. Effects of Leptin-AI and Leptin-AII on Gene Expression of Appetite Regulators in Different Brain Regions
4.7. Measurement of Transcriptional Expression of Target Genes by Real-Time PCR
4.8. Data Transformation and Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Y.Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homolog. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.C.; Clegg, K.E.; Palmiter, R.D. Sensitivity to leptin and susceptibility to seizures of mice lacking neuropeptide Y. Nature 1996, 381, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.D.; Bagnol, D.; Kaelin, C.B.; Ollmann, M.M.; Gantz, I.; Watson, S.J.; Barsh, G.S. Physiological and anatomical circuitry between agouti-related protein and leptin signaling. Endocrinology 1999, 140, 2387–2397. [Google Scholar] [CrossRef] [PubMed]
- Boston, B.A.; Blaydon, K.M.; Varnerin, J.; Cone, R.D. Independent and additive effects of central POMC and leptin pathways on murine obesity. Science 1997, 278, 1641–1644. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, P.; Judge, M.E.; Thim, L.; Ribel, U.; Christjansen, K.N.; Wulff, B.S.; Clausen, J.T.; Jensen, P.B.; Madsen, O.D.; Vrang, N.; et al. Hypothalamic cart is a new anorectic peptide regulated by leptin. Nature 1998, 393, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, E.C.; Myers, M.G. Leptin receptor signaling and the regulation of mammalian physiology. Int. J. Obes. 2008, 32, S8–S12. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.A.; Clark, F.T.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef]
- Tartaglia, L.A. The leptin receptor. J. Biol. Chem. 1997, 272, 6093–6096. [Google Scholar] [CrossRef] [PubMed]
- Elmquist, J.K.; Bjorbaek, C.; Ahima, R.S.; Flier, J.S.; Saper, C.B. Distributions of leptin receptor mRNA isoforms in the rat brain. J. Comp. Neurol. 1998, 395, 535–547. [Google Scholar] [CrossRef]
- Leshan, R.L.; Bjornholm, M.; Munzberg, H.; Myers, M.G., Jr. Leptin receptor signaling and action in the central nervous system. Obesity 2006, 14, 208S–212S. [Google Scholar] [CrossRef] [PubMed]
- Elmquist, J.K.; Elias, C.F.; Saper, C.B. From lesions to leptin: Hypothalamic control of food intake and body weight. Neuron 1999, 22, 221–232. [Google Scholar] [CrossRef]
- Huising, M.O.; Geven, E.J.W.; Kruiswijk, C.P.; Nabuurs, S.B.; Stolte, E.H.; Spanings, F.A.T.; Verburg-Van Kemenade, B.M.L.; Flik, G. Increased leptin expression in common carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation. Endocrinology 2006, 147, 5786–5797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, T.; Uji, S.; Suzuki, T. Identification of cdna coding for a homologue to mammalian leptin from pufferfish, takifugu rubripes. Peptides 2005, 26, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Crespi, E.J.; Denver, R.J. Leptin (ob gene) of the south african clawed frog xenopus laevis. Proc. Natl. Acad. Sci. USA 2006, 103, 10092–10097. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. Leptin in teleostean fish, towards the origins of leptin physiology. J. Chem. Neuroanat. 2014, 61, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Denver, R.J.; Bonett, R.M.; Boorse, G.C. Evolution of leptin structure and function. Neuroendocrinology 2011, 94, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Bernier, N.J.; Nabuurs, S.B.; Flik, G.; Huising, M.O. Two divergent leptin paralogues in zebrafish (Danio rerio) that originate early in teleostean evolution. J. Endocrinol. 2009, 201, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, T.; Murashita, K. Genomic characterization of multiple leptin genes and a leptin receptor gene in the japanese medaka, oryzias latipes. Gen. Comp. Endocrinol. 2009, 161, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Shpilman, M.; Hollander-Cohen, L.; Ventura, T.; Gertler, A.; Levavi-Sivan, B. Production, gene structure and characterization of two orthologs of leptin and a leptin receptor in tilapia. Gen. Comp. Endocrinol. 2014, 207, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Chen, S.; Ren, C.; Hu, C.; Tang, D.; Yan, A. Two isoforms of leptin in the white-clouds mountain minnow (Tanichthys albonubes): Differential regulation by estrogen despite similar response to fasting. Gen. Comp. Endocrinol. 2016, 225, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Prokop, J.W.; Duff, R.J.; Ball, H.C.; Copeland, D.L.; Londraville, R.L. Leptin and leptin receptor: Analysis of a structure to function relationship in interaction and evolution from humans to fish. Peptides 2012, 38, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Ronnestad, I.; Nilsen, T.O.; Murashita, K.; Angotzi, A.R.; Moen, A.G.G.; Stefansson, S.O.; Kling, P.; Bjornsson, B.T.; Kurokawa, T. Leptin and leptin receptor genes in atlantic salmon: Cloning, phylogeny, tissue distribution and expression correlated to long-term feeding status. Gen. Comp. Endocrinol. 2010, 168, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, A.B.; Nisembaum, L.G.; Isorna, E.; Delgado, M.J.; de Pedro, N. Leptins and leptin receptor expression in the goldfish (Carassius auratus). Regulation by food intake and fasting/overfeeding conditions. Peptides 2012, 34, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Gautron, L.; Elmquist, J.K. Sixteen years and counting: An update on leptin in energy balance. J. Clin. Investig. 2011, 121, 2087–2093. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Eykelbosh, A.J.; Peter, R.E. Role of leptin in the control of feeding of goldfish Carassius auratus: Interactions with cholecystokinin, neuropeptide Y and orexin A, and modulation by fasting. Brain Res. 2003, 972, 90–109. [Google Scholar] [CrossRef]
- de Pedro, N.; Martinez-Alvarez, R.; Delgado, M.J. Acute and chronic leptin reduces food intake and body weight in goldfish (Carassius auratus). J. Endocrinol. 2006, 188, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Vivas, Y.; Azpeleta, C.; Feliciano, A.; Velarde, E.; Isorna, E.; Delgado, M.J.; de Pedro, N. Time-dependent effects of leptin on food intake and locomotor activity in goldfish. Peptides 2011, 32, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Peter, R.E. Characterization of two forms of cocaine- and amphetamine-regulated transcript (cart) peptide precursors in goldfish: Molecular cloning and distribution, modulation of expression by nutritional status, and interactions with leptin. Endocrinology 2001, 142, 5076–5088. [Google Scholar] [CrossRef] [PubMed]
- Murashita, K.; Uji, S.; Yamamoto, T.; Ronnestad, I.; Kurokawa, T. Production of recombinant leptin and its effects on food intake in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Phys. B 2008, 150, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Pfundt, B.; Sauerwein, H.; Mielenz, M. Leptin mrna and protein immunoreactivity in adipose tissue and liver of rainbow trout (Oncorhynchus mykiss) and immunohistochemical localization in liver. Anat. Histol. Embryol. 2009, 38, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Chisada, S.; Kurokawa, T.; Murashita, K.; Ronnestad, I.; Taniguchi, Y.; Toyoda, A.; Sakaki, Y.; Takeda, S.; Yoshiura, Y. Leptin receptor-deficient (knockout) medaka, oryzias latipes, show chronical up-regulated levels of orexigenic neuropeptides, elevated food intake and stage specific effects on growth and fat allocation. Gen. Comp. Endocrinol. 2014, 195, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerda-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. In vitro assessment of interactions between appetite-regulating peptides in brain of goldfish (Carassius auratus). Peptides 2014, 61, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Kojima, K.; Shimakura, S.I.; Miura, T.; Uchiyama, M.; Shioda, S.; Ando, H.; Takahashi, A. Relationship between melanin-concentrating hormone- and neuropeptide Y-containing neurons in the goldfish hypothalamus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 153, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Reverter, J.M.; Peter, R.E. Endogenous melanocortin antagonist in fish: Structure, brain mapping, and regulation by fasting of the goldfish agouti-related protein gene. Endocrinology 2003, 144, 4552–4561. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Peter, R.E. Interactions between orexin A, NPY and galanin in the control of food intake of the goldfish, Carassius auratus. Regul. Pept. 2001, 101, 59–72. [Google Scholar] [CrossRef]
- Nakamachi, T.; Shibata, H.; Sakashita, A.; Iinuma, N.; Wada, K.; Konno, N.; Matsuda, K. Orexin A enhances locomotor activity and induces anxiogenic-like action in the goldfish, Carassius auratus. Horm. Behav. 2014, 66, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Wyatt, J.L. Apelin in goldfish (Carassius auratus): Cloning, distribution and role in appetite regulation. Peptides 2009, 30, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Kojima, K.; Shimakura, S.; Wada, K.; Maruyama, K.; Uchiyama, M.; Kikuyama, S.; Shioda, S. Corticotropin-releasing hormone mediates α-melanocyte-stimulating hormone-induced anorexigenic action in goldfish. Peptides 2008, 29, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.H.; Stearns, W.H.; Dundon, T.A.; Schubert, M.; Tso, A.W.K.; Wang, Y.P.; Banks, A.S.; Lavery, H.J.; Haq, A.K.; Maratos-Flier, E.; et al. Stat3 signalling is required for leptin regulation of energy balance but not reproduction. Nature 2003, 421, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Douros, J.D.; Baltzegar, D.A.; Breves, J.P.; Lerner, D.T.; Seale, A.P.; Gordon Grau, E.; Borski, R.J. Prolactin is a major inhibitor of hepatic leptin A synthesis and secretion: Studies utilizing a homologous leptin A ELISA in the tilapia. Gen. Comp. Endocrinol. 2014, 207, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Laborde, C.; Chemardin, P.; Bigey, F.; Combarnous, Y.; Moulin, G.; Boze, H. Overexpression of ovine leptin in Pichia pastoris: Physiological yeast response to leptin production and characterization of the recombinant hormone. Yeast 2004, 21, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Li, W.; Lin, H. Cdna cloning and mrna expression of neuropeptide Y in orange spotted grouper, Epinephelus coioides. Comp. Biochem. Physiol. B 2005, 142, 79–89. [Google Scholar] [CrossRef] [PubMed]
- David, L.; Blum, S.; Feldman, M.W.; Lavi, U.; Hillel, J. Recent duplication of the common carp (Cyprinus carpio L.) genome as revealed by analyses of microsatellite loci. Mol. Biol. Evol. 2003, 20, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Hedges, S.B. A molecular timescale for vertebrate evolution. Nature 1998, 392, 917–920. [Google Scholar] [PubMed]
- Volff, J.N. Genome evolution and biodiversity in teleost fish. Heredity 2005, 94, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [PubMed]
- Li, Y.; Wu, S.; Ouyang, J.; Mao, L.; Li, W.; Lin, H. Expression of insulin-like growth factor-1 of orange-spotted grouper (Epinephelus coioides) in yeast Pichia pastoris. Protein Expr. Purif. 2012, 84, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, T.; Zhong, M.; Jiang, X.; Zhang, L.; Ren, C.; Hu, C. Differential regulation of hepatopancreatic vitellogenin (VTG) gene expression by two putative molt-inhibiting hormones (MIH1/2) in pacific white shrimp (Litopenaeus vannamei). Peptides 2015, 68, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K. Citation-classic-physiological salines for fresh-water teleosts. Curr. Contents Agric. Biol. Environ. Sci. 1986, 46, 20–20. [Google Scholar]
- Volkoff, H.; Peter, R.E. Effects of cart peptides on food consumption, feeding and associated behaviors in the goldfish, Carassius auratus: Actions on neuropeptide Y- and orexin A induced feeding. Brain Res. 2000, 887, 125–133. [Google Scholar] [CrossRef]
- Chen, T.; Ren, C.H.; Wang, Y.H.; Gao, Y.; Wong, N.K.; Zhang, L.P.; Hu, C.Q. Crustacean cardioactive peptide (CCAP) of the pacific white shrimp (Litopenaeus vannamei): Molecular characterization and its potential roles in osmoregulation and freshwater tolerance. Aquaculture 2016, 451, 405–412. [Google Scholar] [CrossRef]
- Wong, M.K.H.; Sze, K.H.; Chen, T.; Cho, C.K.; Law, H.C.H.; Chu, I.K.; Wong, A.O.L. Goldfish spexin: Solution structure and novel function as a satiety factor in feeding control. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E348–E366. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Target/Accession No. (Primer Sequences, 5′-3′) | PCR Condition | Cycle | Tm | Product Size | |||
|---|---|---|---|---|---|---|---|
| Denaturing | Annealing | Extension | Detection | ||||
| NPY/M87297 | |||||||
| GTAGTGTTGCGGGTAGCGA | 94 °C | 64 °C | 72 °C | 88 °C | ×35 | 92 °C | 234 bp |
| CAGACACCCCGACCCAAG | 30 s | 30 s | 30 s | 20 s | |||
| AgRP/AJ555492 | |||||||
| TGGCATCACATCCAAACCT | 94 °C | 64 °C | 72 °C | 82 °C | ×35 | 88 °C | 230 bp |
| CAGGTGATGACCCAAGCAG | 30 s | 30 s | 30 s | 20 s | |||
| Orexin/DQ923590 | |||||||
| GCAGAGCTGCTCATTGTTGACGTT | 94 °C | 64 °C | 72 °C | 84 °C | ×35 | 82 °C | 286 bp |
| AACCTTGTGATTACCTCAGGAGT | 30 s | 30 s | 30 s | 20 s | |||
| Apelin/FJ755698 | |||||||
| GAGCATAGCAAAGAGCTGGA | 94 °C | 64 °C | 72 °C | 89 °C | ×35 | 94 °C | 340 bp |
| GCTGAGGATGAGTGGCTTGT | 30 s | 30 s | 30 s | 20 s | |||
| CART/AF288810 | |||||||
| CCAAAGGACCCGAATCTGA | 94 °C | 64 °C | 72 °C | 82 °C | ×35 | 90 °C | 171 bp |
| TTTGCCGATTCTTGACCCT | 30 s | 30 s | 30 s | 20 s | |||
| CCK/CAU70865 | |||||||
| CCGCAGTCTCAGAAGATGGG | 94 °C | 64 °C | 72 °C | 87 °C | ×35 | 91 °C | 197 bp |
| GGAGGGGCTTCTGCGATA | 30 s | 30 s | 30 s | 20 s | |||
| MCH/AM403730 | |||||||
| AGGCTTGAGCGAGAACTTGG | 94 °C | 64 °C | 72 °C | 86 °C | ×35 | 91 °C | 272 bp |
| CCCAGAAGACCTACACCTCCC | 30 s | 30 s | 30 s | 20 s | |||
| POMC/AJ431209 | |||||||
| AAGCGCTCCTACTCCATGGA | 94 °C | 60 °C | 72 °C | 83 °C | ×35 | 85 °C | 282 bp |
| CTCGTCCCAGGACTTCATGAA | 30 s | 30 s | 30 s | 20 s | |||
| β-actin/AB039726 | |||||||
| CTGGTATCGTGATGGACTCT | 94 °C | 56 °C | 72 °C | 87 °C | ×35 | 91 °C | 285 bp |
| AGCTCATAGCTCTTCTCCAG | 30 s | 30 s | 30 s | 20 s | |||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, A.-F.; Chen, T.; Chen, S.; Ren, C.-H.; Hu, C.-Q.; Cai, Y.-M.; Liu, F.; Tang, D.-S. Goldfish Leptin-AI and Leptin-AII: Function and Central Mechanism in Feeding Control. Int. J. Mol. Sci. 2016, 17, 783. https://doi.org/10.3390/ijms17060783
Yan A-F, Chen T, Chen S, Ren C-H, Hu C-Q, Cai Y-M, Liu F, Tang D-S. Goldfish Leptin-AI and Leptin-AII: Function and Central Mechanism in Feeding Control. International Journal of Molecular Sciences. 2016; 17(6):783. https://doi.org/10.3390/ijms17060783
Chicago/Turabian StyleYan, Ai-Fen, Ting Chen, Shuang Chen, Chun-Hua Ren, Chao-Qun Hu, Yi-Ming Cai, Fang Liu, and Dong-Sheng Tang. 2016. "Goldfish Leptin-AI and Leptin-AII: Function and Central Mechanism in Feeding Control" International Journal of Molecular Sciences 17, no. 6: 783. https://doi.org/10.3390/ijms17060783
APA StyleYan, A. -F., Chen, T., Chen, S., Ren, C. -H., Hu, C. -Q., Cai, Y. -M., Liu, F., & Tang, D. -S. (2016). Goldfish Leptin-AI and Leptin-AII: Function and Central Mechanism in Feeding Control. International Journal of Molecular Sciences, 17(6), 783. https://doi.org/10.3390/ijms17060783