
Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels

Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
2.2. Correlation of Serum Sphingolipids (SLs) with Hepatitis C Virus (HCV)/Human Immunodeficiency Virus (HIV) Coinfection, Angiopoietin-Like 3 (ANGPTL3) and Biochemical Parameters
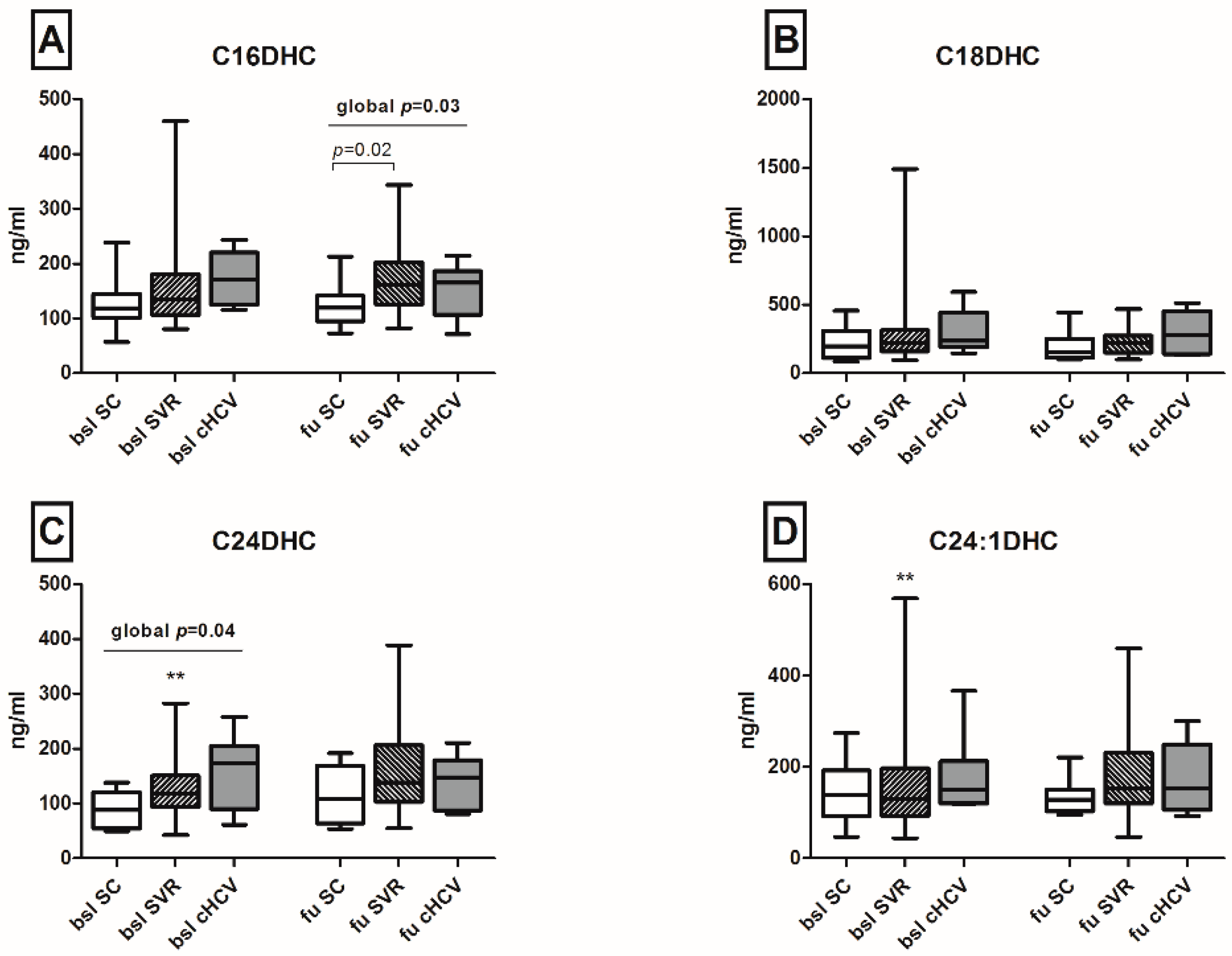
2.3. Spontaneous Clearance of HCV Restores Serum C24-Ceramide (C24-Cer) Levels
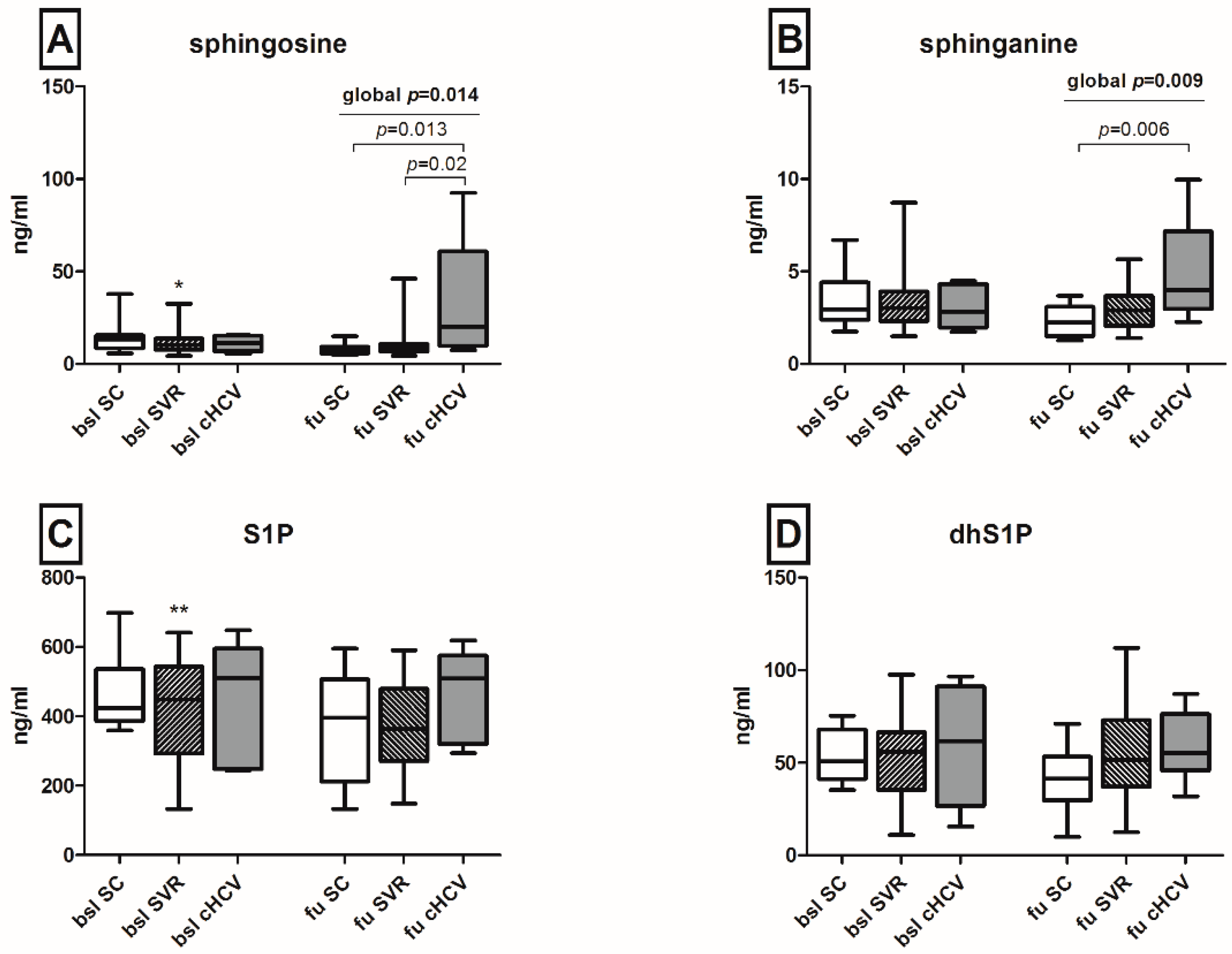
2.4. Upregulation of Sphingosine and Sphinganine Levels upon the Persistence of HCV
2.5. Univariate Analysis of Baseline Markers Associated with Spontaneous HCV Clearance
3. Discussion
4. Experimental Section
4.1. Patients with Acute HCV Infection
4.2. Determination of Sphingolipid Concentrations by High-Performance Liquid Chromatography Tandem Mass Spectrometry
4.3. Detection of ANGTL3 Serum Concentrations
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HCV | hepatitis C virus |
| IDU | injecting drug users |
| MSM | men having sex with men |
| HIV | human immunodeficiency virus |
| DAA | direct-acting antiviral |
| SL | sphingolipid |
| ANGPTL3 | angiopoietin-like 3 |
| Cer | ceramide |
| DHC | dihydroceramide |
| ALT | alanine aminotransferase |
| SVR | sustained viral response |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| S1P | sphingosine 1-phosphate |
| dhS1P | sphinganine 1-phosphate |
| APCI | atmospheric pressure chemical ionization |
| ESI | electrospray ionization |
| MRM | multiple reaction monitoring |
| CI | confidence interval |
| PEG-IFN | PEGylated interferon |
| RBV | ribavirin |
| INR | international normalized ratio |
References
- Hajarizadeh, B.; Grebely, J.; Dore, G.J. Epidemiology and natural history of HCV infection. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Hullegie, S.J.; Arends, J.E.; Rijnders, B.J.; Irving, W.L.; Salmon, D.; Prins, M.; Wensing, A.M.; Klenerman, P.; Leblebicioglu, H.; Boesecke, C.; et al. Current knowledge and future perspectives on acute hepatitis C infection. Clin. Microbiol. Infect. 2015, 21, 797. [Google Scholar] [CrossRef] [PubMed]
- Miyanari, Y.; Atsuzawa, K.; Usuda, N.; Watashi, K.; Hishiki, T.; Zayas, M.; Bartenschlager, R.; Wakita, T.; Hijikata, M.; Shimotohno, K. The lipid droplet is an important organelle for hepatitis C virus production. Nat. Cell Biol. 2007, 9, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.T.; Lee, K.J.; Aizaki, H.; Hwang, S.B.; Lai, M.M. Hepatitis C virus RNA replication occurs on a detergent-resistant membrane that cofractionates with caveolin-2. J. Virol. 2003, 77, 4160–4168. [Google Scholar] [CrossRef] [PubMed]
- Saxena, V.; Lai, C.K.; Chao, T.C.; Jeng, K.S.; Lai, M.M. Annexin A2 is involved in the formation of hepatitis C virus replication complex on the lipid raft. J. Virol. 2012, 86, 4139–4150. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Luo, G. Apolipoprotein E but not B is required for the formation of infectious hepatitis C virus particles. J. Virol. 2009, 83, 12680–12691. [Google Scholar] [CrossRef] [PubMed]
- Aizaki, H.; Morikawa, K.; Fukasawa, M.; Hara, H.; Inoue, Y.; Tani, H.; Saito, K.; Nishijima, M.; Hanada, K.; Matsuura, Y.; Lai, M.M.C.; et al. Critical role of virion-associated cholesterol and sphingolipid in hepatitis C virus infection 2. J. Virol. 2008, 82, 5715–5724. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Ikeda, K.; Sudoh, M.; Tokunaga, Y.; Suzuki, A.; Weng, L.; Ohta, M.; Tobita, Y.; Okano, K.; Ozeki, K.; et al. Self-enhancement of hepatitis C virus replication by promotion of specific sphingolipid biosynthesis 1. PLoS Pathog. 2012, 8, e1002860. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Okamoto, K.; Aoki, M.; Kato, H.; Katsume, A.; Ohta, A.; Tsukuda, T.; Shimma, N.; Aoki, Y.; Arisawa, M.; et al. Host sphingolipid biosynthesis as a target for hepatitis C virus therapy 2. Nat. Chem. Biol. 2005, 1, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Aizaki, H.; Lee, K.J.; Sung, V.M.; Ishiko, H.; Lai, M.M. Characterization of the hepatitis C virus RNA replication complex associated with lipid rafts. Virology 2004, 324, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Voisset, C.; Lavie, M.; Helle, F.; Op de, B.A.; Bilheu, A.; Bertrand-Michel, J.; Tercé, F.; Cocquerel, L.; Wychowski, C.; Vu-Dac, N.; et al. Ceramide enrichment of the plasma membrane induces CD81 internalization and inhibits hepatitis C virus entry. Cell Microbiol. 2008, 10, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Hirata, Y.; Arai, M.; Kohara, M.; Wakita, T.; Watashi, K.; Shimotohno, K.; He, Y.; Zhong, J.; Toyoda, T. Sphingomyelin activates hepatitis C virus RNA polymerase in a genotype-specific manner. J. Virol. 2010, 84, 11761–11770. [Google Scholar] [CrossRef] [PubMed]
- Yamane, D.; McGivern, D.R.; Wauthier, E.; Yi, M.; Madden, V.J.; Welsch, C.; Antes, I.; Wen, Y.; Chugh, P.E.; McGee, C.E.; et al. Regulation of the hepatitis C virus RNA replicase by endogenous lipid peroxidation 1. Nat. Med. 2014, 20, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Corey, K.E.; Kane, E.; Munroe, C.; Barlow, L.L.; Zheng, H.; Chung, R.T. Hepatitis C virus infection and its clearance alter circulating lipids: Implications for long-term follow-up. Hepatology 2009, 50, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Shimizugawa, T.; Ono, M.; Shimamura, M.; Yoshida, K.; Ando, Y.; Koishi, R.; Ueda, K.; Inaba, T.; Minekura, H.; Kohama, T.; et al. ANGPTL3 decreases very low density lipoprotein triglyceride clearance by inhibition of lipoprotein lipase. J. Biol. Chem. 2002, 277, 33742–33748. [Google Scholar] [CrossRef] [PubMed]
- Foka, P.; Karamichali, E.; Dalagiorgou, G.; Serti, E.; Doumba, P.P.; Pissas, G.; Kakkanas, A.2.; Kazazi, D.; Kochlios, E.; Gaitanou, M.; et al. Hepatitis C virus modulates lipid regulatory factor Angiopoietin-like 3 gene expression by repressing HNF-1α activity 1. J. Hepatol. 2014, 60, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Park, I.N.; Cho, I.J.; Kim, S.G. Ceramide negatively regulates glutathione S-transferase gene transactivation via repression of hepatic nuclear factor-1 that is degraded by the ubiquitin proteasome system 2. Mol. Pharmacol. 2004, 65, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Ramcharran, D.; Wahed, A.S.; Conjeevaram, H.S.; Evans, R.W.; Wang, T.; Belle, S.H.; Yee, L.J.; Virahep-C Study Group. Associations between serum lipids and hepatitis C antiviral treatment efficacy. Hepatology 2010, 52, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Sultanik, P.; Mallet, V.; Lagaye, S.; Casrouge, A.; Dorival, C.; Barthe, Y.; Fontaine, H.; Hézode, C.; Mottez, E.; Bronowicki, J.P.; et al. Plasma apolipoprotein H limits HCV replication and associates with response to NS3 protease inhibitors-based therapy. Liver Int. 2015, 35, 1833–1844. [Google Scholar] [CrossRef] [PubMed]
- Laird, M.E.; Mohsen, A.; Duffy, D.; Mamdouh, R.; LeFouler, L.; Casrouge, A.; El-Daly, M.; Rafik, M.; Abdel-Hamid, M.; Soulier, A.; et al. Apolipoprotein H expression is associated with IL28B genotype and viral clearance in hepatitis C virus infection. J. Hepatol. 2014, 61, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Muhle, C.; Ferreiros, N.; Schroeter, S.; Bogdanou, D.; Schwalm, S.; Hintereder, G.; Kornhuber, J.; Zeuzem, S.; Sarrazin, C.; et al. Serum acid sphingomyelinase is upregulated in chronic hepatitis C infection and non alcoholic fatty liver disease. Biochim. Biophys. Acta 2014, 1841, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Ferreiros, N.; Bon, D.; Schwalm, S.; Dietz, J.; Berkowski, C.; Fitting, D.; Herrmann, E.; Zeuzem, S.; Sarrazin, C.; et al. Variations in serum sphingolipid levels associate with liver fibrosis progression and poor treatment outcome in hepatitis C virus but not hepatitis B virus infection. Hepatology 2015, 61, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Ferreiros, N.; Waidmann, O.; Bon, D.; Schroeter, S.; Koch, A.; Herrmann, E.; Zeuzem, S.; Kronenberger, B.; Pfeilschifter, J. Serum sphingolipid variations associate with hepatic decompensation and survival in patients with cirrhosis 1. PLoS ONE 2015, 10, e0138130. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, P.; Munoz-Olaya, J.M.; Gagliostro, V.; Casas, J.; Ghidoni, R.; Fabrias, G. Dihydroceramide intracellular increase in response to resveratrol treatment mediates autophagy in gastric cancer cells. Cancer Lett. 2009, 282, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Mutimer, D.; Aghemo, A.; Diepolder, H.; Negro, F.; Robaeys, G.; Ryder, S.; Zoulim, F. EASL Clinical Practice Guidelines: Management of hepatitis C virus infection. J. Hepatol. 2014, 60, 392–420. [Google Scholar]
- Vieira, C.R.; Munoz-Olaya, J.M.; Sot, J.; Jimenez-Baranda, S.; Izquierdo-Useros, N.; Abad, J.L.; Apellániz, B.; Delgado, R.; Martinez-Picado, J.; Alonso, A.; et al. Dihydrosphingomyelin impairs HIV-1 infection by rigidifying liquid-ordered membrane domains. Chem. Biol. 2010, 17, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Nemoto-Sasaki, Y.; Tanikawa, T.; Oka, S.; Tsuchiya, K.; Zama, K.; Mitsutake, S.; Sugiura, T.; Yamashita, A. Sphingomyelin synthase 2, but not sphingomyelin synthase 1, is involved in HIV-1 envelope-mediated membrane fusion. J. Biol. Chem. 2014, 289, 30842–30856. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, C.M.; Rawat, S.S.; Puri, A.; Wang, J.M.; Ruscetti, F.W.; Blumenthal, R. Ceramide, a target for antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2004, 101, 15452–15457. [Google Scholar] [CrossRef] [PubMed]
- Pewzner-Jung, Y.; Brenner, O.; Braun, S.; Laviad, E.L.; Ben-Dor, S.; Feldmesser, E.; Horn-Saban, S.; Amann-Zalcenstein, D.; Raanan, C.; Berkutzki, T.; et al. A critical role for ceramide synthase 2 in liver homeostasis: II. insights into molecular changes leading to hepatopathy 2. J. Biol. Chem. 2010, 285, 10911–10923. [Google Scholar] [CrossRef] [PubMed]
- Zigdon, H.; Kogot-Levin, A.; Park, J.W.; Goldschmidt, R.; Kelly, S.; Merrill, A.H., Jr.; Scherz, A.; Pewzner-Jung, Y.; Saada, A.; Futerman, A.H. Ablation of ceramide synthase 2 causes chronic oxidative stress due to disruption of the mitochondrial respiratory chain. J. Biol. Chem. 2013, 288, 4947–4956. [Google Scholar] [CrossRef] [PubMed]
- Brault, C.; Levy, P.; Duponchel, S.; Michelet, M.; Salle, A.; Pecheur, E.I.; Plissonnier, M.L.; Parent, R.; Véricel, E.; Ivanov, A.V.; et al. Glutathione peroxidase 4 is reversibly induced by HCV to control lipid peroxidation and to increase virion infectivity1. Gut 2014, 65, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Farinati, F.; Cardin, R.; de, M.N.; Della, L.G.; Marafin, C.; Lecis, E. Iron storage, lipid peroxidation and glutathione turnover in chronic anti-HCV positive hepatitis 15. J. Hepatol. 1995, 22, 449–456. [Google Scholar] [CrossRef]
- Grosch, S.; Schiffmann, S.; Geisslinger, G. Chain length-specific properties of ceramides. Prog. Lipid Res. 2012, 51, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.A.; Syder, A.J.; Lederer, S.L.; Diamond, D.L.; Paeper, B.; Rice, C.M.; Katze, M.G. Genomic analysis reveals a potential role for cell cycle perturbation in HCV-mediated apoptosis of cultured hepatocytes. PLoS Pathog. 2009, 5, e1000269. [Google Scholar] [CrossRef] [PubMed]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid Res. 2013, 54, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Guillen, M.A.; Mejia, F.A.; Villena, J.; Turin, C.G.; Carcamo, C.P.; Ticse, R. Insulin resistance by homeostasis model assessment in HIV-infected patients on highly active antiretroviral therapy: Cross-sectional study. Diabetol. Metab. Syndr. 2015, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Conca, P.; Ariello, M.; Mastrolia, M. Does a lower insulin resistance affect antiviral therapy response in patients suffering from HCV related chronic hepatitis? Gut 2006, 55, 585. [Google Scholar] [PubMed]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Bui, H.H.; Sanders, P.; Siddall, P.; Kuo, M.S.; Perreault, L. Serum sphingolipids: Relationships to insulin sensitivity and changes with exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E398–E408. [Google Scholar] [CrossRef] [PubMed]
- Dietz, J.; Lutz, T.; Knecht, G.; Gute, P.; Berkowski, C.; Lange, C.M.; Khaykin, P.; Stephan, C.; Brodt, H.R.; Herrmann, E.; et al. Evolution and function of the HCV NS3 protease in patients with acute hepatitis C and HIV coinfection 1. Virology 2015, 485, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Benito, J.; Bhagani, S.; Boesecke, C.; Deterding, K.; Dominguez, S.; Fisher, M.; Fontanet, A.; Garcia, D.; Gilson, R.; et al. Acute hepatitis C in HIV-infected individuals: Recommendations from the European AIDS Treatment Network (NEAT) consensus conference. AIDS 2011, 25, 399–409. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Patients with Acute HCV Infection (n = 60) |
|---|---|
| Age, years: mean (range) | 42 (22–61) |
| Sex, male, n = (%) | 56 (93.4) |
| Female, n = (%) | 4 (6.6) |
| ALT, IU/L, mean (range) | 505 (28–2738) |
| AST, IU/L, mean (range) | 242 (21–1957) |
| γGT, IU/L, mean (range) | 250 (25–1064) |
| Bilirubin, mg/dL, mean (range) | 1.0 (0.2–13) |
| Thrombocytes, cells/µL, mean (range) | 210 (115–324) |
| INR, mean (range) | 1.05 (0.9–1.3) |
| IL28B-rs12979860 genotype, (CC/non_CC, n = (%)) | 26 (43.4)/34 (56.6) |
| HCV genotype | |
| 1, n = | 44 |
| 1a, n = | 39 |
| 1b, n = | 4 |
| Subtype not determined, n = | 1 |
| 2, n = | 2 |
| 3, n = | 3 |
| 4, n = | 11 |
| HCV viral load, IU/mL: mean (range) | 7.7 × 106 (15–(8.6 × 107)) |
| Estimated duration between infection to diagnosis (weeks) *: mean (range) | 7 (1–20) |
| Duration diagnosis to initiation of therapy (weeks): mean (range) | 14 (3–33) |
| Duration diagnosis to spontaneous clearance (weeks): mean (range) | 30 (4–100) |
| Spontaneous clearance, n = (%) | 11 (18.3) |
| Antiviral therapy of acute HCV, n = (%) | 45 (75) |
| SVR, n = (% of follow-up completed) | 43 (95.5) |
| Non-response, n = (% of follow-up completed) | 2 (4.5) |
| Lost to follow-up, n = | 1 (excluded from the study) |
| Persistence of HCV infection: n = (%) | 6 (10) |
| HIV coinfection: n = (%) | 51 (85) |
| Patients on ART: n = (%) | 38 (74.5) |
| CD4 count, cells/µL: mean (range) | 561 (207–1561) |
| HIV patients with <50 copies/mL: n = (% of patients on ART) | 32 (62.7) |
| Parameters | Spontaneous Clearance (n = 11) | Persistence of HCV (n = 6) | p-Value |
|---|---|---|---|
| Age, years: mean (range) | 42 (22–54) | 43.5 (37–56) | 0.2 |
| ALT, IU/L: mean (range) | 437 (113–2217) | 291.5 (43–922) | 0.4 |
| AST, IU/L: mean (range) | 224 (51–745) | 132 (33–402) | 0.1 |
| γGT, IU/L: mean (range) | 220 (31–310) | 205.5 (93–407) | 0.7 |
| Bilirubin, mg/dL: mean (range) | 1.35 (0.5–2.2) | 0.765 (0.5–2.64) | 0.4 |
| Thrombocytes, cells/µL: mean (range) | 202.5 (145–307) | 209 (153–255) | 1.0 |
| INR: mean (range) | 1.0 (0.9–1.1) | 0.9 (0.9–1.2) | 0.6 |
| HCV genotype (n =, 1/non-1) | 6/5 | 6/0 | 0.056 |
| HCV viral load, IU/mL: mean (range) | 1.4 × 106 (2780–(6.9 × 107)) | 1.2 × 106 (64,900–(8.6 × 107)) | 0.8 |
| HIV coinfection: (n =, yes/no) | 8/3 | 6/0 | 0.1 |
| HIV patients with <50 copies/mL: (n =, yes/no) | 5/3 | 5/1 | 0.4 |
| CD4 count, cells/µL: mean (range) | 564.7 (312–850) | 480 (207–766) | 0.5 |
| IL28B-rs12979860 genotype: (CC/non_CC, n = (%)) | 5/6 | 1/5 | 0.2 |
| C24Cer, ng/mL: mean (range) | 1710 (626–2270) | 1815 (808–2980) | 0.4 |
| C24DHC, ng/mL: mean (range) | 88.4 (49.8–138) | 173.5 (61.5–258) | 0.047 |
| C16Cer, ng/mL: mean (range) | 417 (218–743) | 459 (310–534) | 0.8 |
| C16DHC, ng/mL: mean (range) | 119 (57.1–239) | 171.5 (116–244) | 0.07 |
| Sphingosine, ng/mL: mean (range) | 13.3 (5.7–37.8) | 11.1 (5.5–15.6) | 0.6 |
| Sphinganine, ng/mL: mean (range) | 2.96 (1.75–6.71) | 2.83 (1.74–4.49) | 0.6 |
| S1P, ng/mL: mean (range) | 424 (359–698) | 511 (244–648) | 0.8 |
| dhS1P, ng/mL: mean (range) | 50.7 (35.3–75.3) | 61.6 (15.6–96.6) | 0.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grammatikos, G.; Dietz, J.; Ferreiros, N.; Koch, A.; Dultz, G.; Bon, D.; Karakasiliotis, I.; Lutz, T.; Knecht, G.; Gute, P.; et al. Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels. Int. J. Mol. Sci. 2016, 17, 922. https://doi.org/10.3390/ijms17060922
Grammatikos G, Dietz J, Ferreiros N, Koch A, Dultz G, Bon D, Karakasiliotis I, Lutz T, Knecht G, Gute P, et al. Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels. International Journal of Molecular Sciences. 2016; 17(6):922. https://doi.org/10.3390/ijms17060922
Chicago/Turabian StyleGrammatikos, Georgios, Julia Dietz, Nerea Ferreiros, Alexander Koch, Georg Dultz, Dimitra Bon, Ioannis Karakasiliotis, Thomas Lutz, Gaby Knecht, Peter Gute, and et al. 2016. "Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels" International Journal of Molecular Sciences 17, no. 6: 922. https://doi.org/10.3390/ijms17060922
APA StyleGrammatikos, G., Dietz, J., Ferreiros, N., Koch, A., Dultz, G., Bon, D., Karakasiliotis, I., Lutz, T., Knecht, G., Gute, P., Herrmann, E., Zeuzem, S., Mavromara, P., Sarrazin, C., & Pfeilschifter, J. (2016). Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels. International Journal of Molecular Sciences, 17(6), 922. https://doi.org/10.3390/ijms17060922