
Role of the Renin-Angiotensin-Aldosterone System beyond Blood Pressure Regulation: Molecular and Cellular Mechanisms Involved in End-Organ Damage during Arterial Hypertension

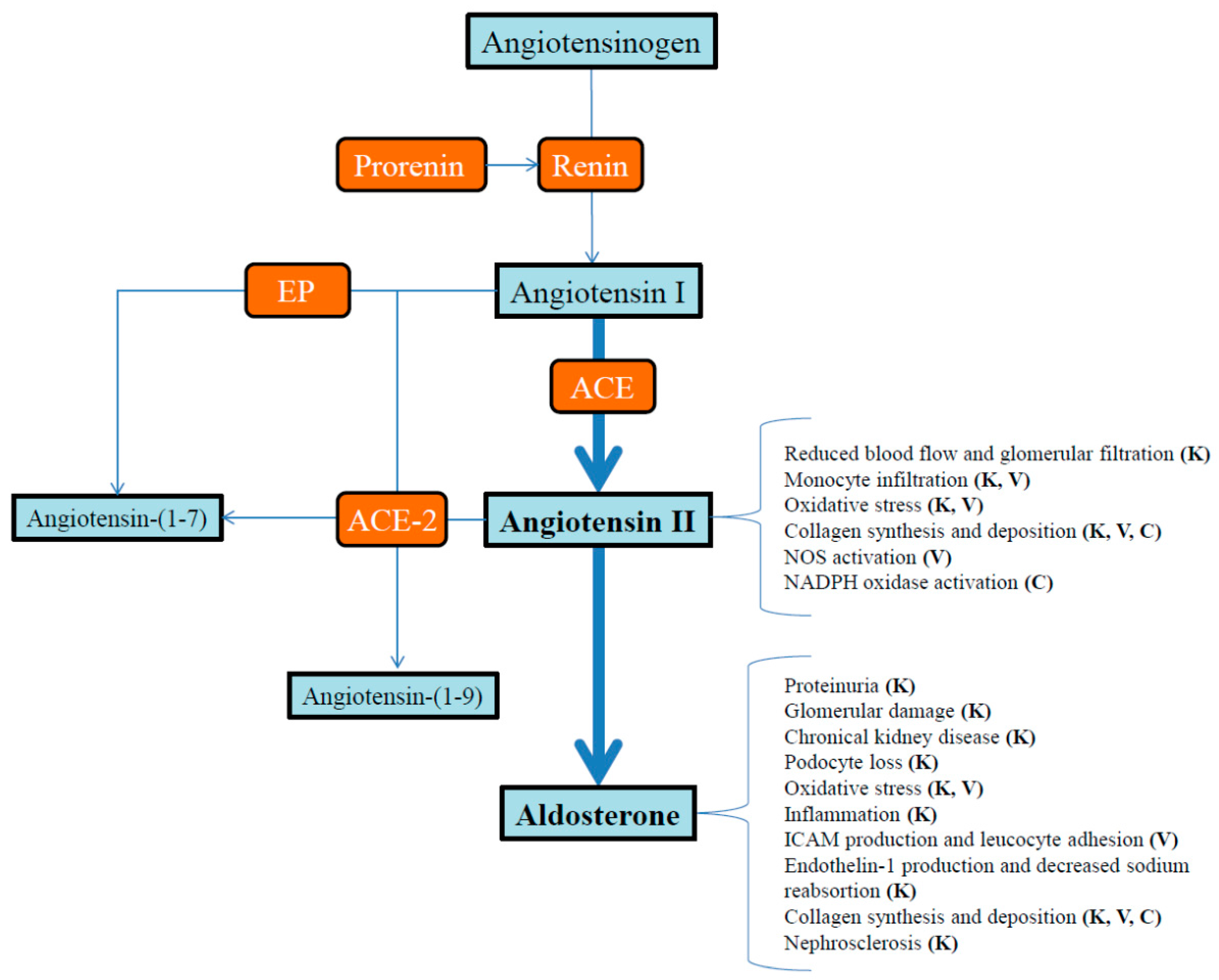{kind=link}
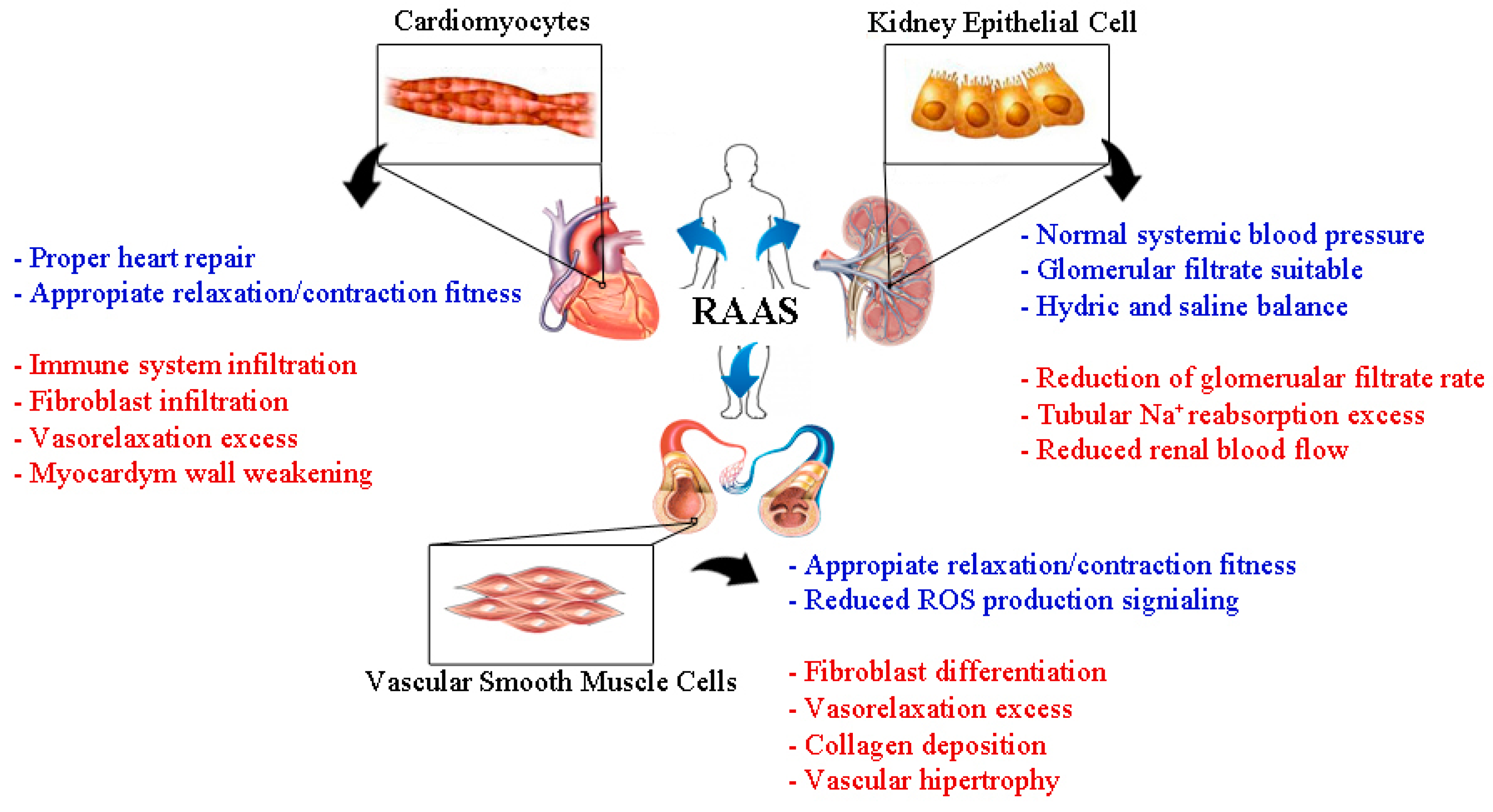{kind=link}
Abstract
:1. Introduction
2. The Role of Angiotensin II and Aldosterone-Dependent End-Organ Damage during Arterial Hypertension
3. Fibrosis, Inflammation, and Their Relation to Arterial Hypertension
4. RAAS Molecules Implicated in Kidney Damage in Arterial Hypertension
5. Vascular System and Mycoardium Damage by RAAS
6. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
Abbreviations
| Ang II | Angiotensin II |
| ARTS-HF | Mineralocorticoid Receptor antagonist Tolerability Study-Heart Failure |
| AT1-R | Angiotensin II type 1 receptor |
| AT2-R | Angiotensin II type 2 receptor |
| EPHESUS | Eplerenone post-acute myocardial infarction heart and survival study |
| MR | Mineralocorticoid receptor |
| RAAS | Renin-Angiotensin-Aldosterone system |
| RALES | Randomized Aldactone evaluation study |
| ROS | Reactive oxygen species |
References
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Milliez, P.; Girerd, X.; Plouin, P.F.; Blacher, J.; Safar, M.E.; Mourad, J.J. Evidence for an increased rate of cardiovascular events in patients with primary aldosteronism. J. Am. Coll. Cardiol. 2005, 45, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Born-Frontsberg, E.; Reincke, M.; Rump, L.C.; Hahner, S.; Diederich, S.; Lorenz, R.; Allolio, B.; Seufert, J.; Schirpenbach, C.; Beuschlein, F.; et al. Cardiovascular and cerebrovascular comorbidities of hypokalemic and normokalemic primary aldosteronism: Results of the German Conn’s registry. J. Clin. Endocrinol. Metab. 2009, 94, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Lifton, R.P.; Gharavi, A.G.; Geller, D.S. Molecular mechanisms of human hypertension. Cell 2001, 104, 545–556. [Google Scholar] [CrossRef]
- Martins, L.C.; Figueiredo, V.N.; Quinaglia, T.; Boer-Martins, L.; Yugar-Toledo, J.C.; Martin, J.F.; Demacq, C.; Pimenta, E.; Calhoun, D.A.; Moreno, H., Jr. Characteristics of resistant hypertension: Ageing, body mass index, hyperaldosteronism, cardiac hypertrophy and vascular stiffness. J. Hum. Hypertens. 2011, 25, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, D.A.; Sharma, K. The role of aldosteronism in causing obesity-related cardiovascular risk. Cardiol. Clin. 2010, 28, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Conn, J.W. Aldosterone in clinical medicine: Past, present, and future. AMA Arch. Intern. Med. 1956, 97, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Te Riet, L.; van Esch, J.H.; Roks, A.J.; van den Meiracker, A.H.; Danser, A.H. Hypertension: Renin-angiotensin-aldosterone system alterations. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Jan Danser, A.H.; Batenburg, W.W.; van Esch, J.H. Prorenin and the (pro)renin receptor—An update. Nephrol. Dial. Transplant. 2007, 22, 1288–1292. [Google Scholar] [CrossRef] [PubMed]
- Putnam, K.; Shoemaker, R.; Yiannikouris, F.; Cassis, L.A. The renin-angiotensin system: A target of and contributor to dyslipidemias, altered glucose homeostasis, and hypertension of the metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1219–H1230. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Trask, A.J.; Jessup, J.A. Advances in biochemical and functional roles of angiotensin-converting enzyme 2 and angiotensin-(1-7) in regulation of cardiovascular function. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2281–H2290. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M. Angiotension-(1-7) and antihypertensive mechanisms. J. Nephrol. 1998, 11, 278–283. [Google Scholar] [PubMed]
- Spat, A.; Hunyady, L. Control of aldosterone secretion: A model for convergence in cellular signaling pathways. Physiol. Rev. 2004, 84, 489–539. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.O. Mechanisms of salt and water retention in congestive heart failure. The importance of aldosterone. Am. J. Med. 1960, 29, 486–507. [Google Scholar] [CrossRef]
- Gros, R.; Ding, Q.; Sklar, L.A.; Prossnitz, E.E.; Arterburn, J.B.; Chorazyczewski, J.; Feldman, R.D. GPR30 expression is required for the mineralocorticoid receptor-independent rapid vascular effects of aldosterone. Hypertension 2011, 57, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, C.; Benesic, A.; Krug, A.W.; Freudinger, R.; Mildenberger, S.; Gassner, B.; Gekle, M. Human mineralocorticoid receptor expression renders cells responsive for nongenotropic aldosterone actions. Mol. Endocrinol. 2005, 19, 1697–1710. [Google Scholar] [CrossRef] [PubMed]
- Pacurari, M.; Kafoury, R.; Tchounwou, P.B.; Ndebele, K. The renin-angiotensin-aldosterone system in vascular inflammation and remodeling. Int. J. Inflamm. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Buglioni, A.; Cannone, V.; Cataliotti, A.; Sangaralingham, S.J.; Heublein, D.M.; Scott, C.G.; Bailey, K.R.; Rodeheffer, R.J.; Dessi-Fulgheri, P.; Sarzani, R.; et al. Circulating aldosterone and natriuretic peptides in the general community: Relationship to cardiorenal and metabolic disease. Hypertension 2015, 65, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Bauersachs, J.; Jaisser, F.; Toto, R. Mineralocorticoid receptor activation and mineralocorticoid receptor antagonist treatment in cardiac and renal diseases. Hypertension 2015, 65, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Tomaschitz, A.; Pilz, S.; Ritz, E.; Obermayer-Pietsch, B.; Pieber, T.R. Aldosterone and arterial hypertension. Nat. Rev. Endocrinol. 2010, 6, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, I.Z.; Mendelsohn, M.E. Angiotensin II and aldosterone regulate gene transcription via functional mineralocortocoid receptors in human coronary artery smooth muscle cells. Circ. Res. 2005, 96, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Derfoul, A.; Robertson, N.M.; Lingrel, J.B.; Hall, D.J.; Litwack, G. Regulation of the human Na/K-ATPase β1 gene promoter by mineralocorticoid and glucocorticoid receptors. J. Biol. Chem. 1998, 273, 20702–20711. [Google Scholar] [CrossRef] [PubMed]
- Naray-Fejes-Toth, A.; Fejes-Toth, G. The SGK, an aldosterone-induced gene in mineralocorticoid target cells, regulates the epithelial sodium channel. Kidney Int. 2000, 57, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Zannad, F.; Remme, W.J.; Cody, R.; Castaigne, A.; Perez, A.; Palensky, J.; Wittes, J. The effect of spironolactone on morbidity and mortality in patients with severe heart failure. Randomized aldactone evaluation study investigators. N. Engl. J. Med. 1999, 341, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Remme, W.; Zannad, F.; Neaton, J.; Martinez, F.; Roniker, B.; Bittman, R.; Hurley, S.; Kleiman, J.; Gatlin, M.; et al. Eplerenone, a selective aldosterone blocker, in patients with left ventricular dysfunction after myocardial infarction. N. Engl. J. Med. 2003, 348, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, P.; Menard, J.; Fay, R.; Gustafsson, F.; Pitt, B.; Zannad, F. Eplerenone survival benefits in heart failure patients post-myocardial infarction are independent from its diuretic and potassium-sparing effects. Insights from an ephesus (eplerenone post-acute myocardial infarction heart failure efficacy and survival study) substudy. J. Am. Coll. Cardiol. 2011, 58, 1958–1966. [Google Scholar] [PubMed]
- Hansson, L.; Lindholm, L.H.; Niskanen, L.; Lanke, J.; Hedner, T.; Niklason, A.; Luomanmaki, K.; Dahlof, B.; de Faire, U.; Morlin, C.; et al. Effect of angiotensin-converting-enzyme inhibition compared with conventional therapy on cardiovascular morbidity and mortality in hypertension: The Captopril Prevention Project (CAPPP) randomised trial. Lancet 1999, 353, 611–616. [Google Scholar] [CrossRef]
- Hansson, L.; Lindholm, L.H.; Ekbom, T.; Dahlof, B.; Lanke, J.; Schersten, B.; Wester, P.O.; Hedner, T.; de Faire, U. Randomised trial of old and new antihypertensive drugs in elderly patients: Cardiovascular mortality and morbidity the swedish trial in old patients with hypertension-2 study. Lancet 1999, 354, 1751–1756. [Google Scholar] [CrossRef]
- Schmieder, R.E.; Martus, P.; Klingbeil, A. Reversal of left ventricular hypertrophy in essential hypertension. A meta-analysis of randomized double-blind studies. JAMA 1996, 275, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Viberti, G.; Wheeldon, N.M.; MicroAlbuminuria Reduction With VSI. Microalbuminuria reduction with valsartan in patients with type 2 diabetes mellitus: A blood pressure-independent effect. Circulation 2002, 106, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.D.; Gurley, S.B.; Herrera, M.J.; Ruiz, P.; Griffiths, R.; Kumar, A.P.; Kim, H.S.; Smithies, O.; Le, T.H.; Coffman, T.M. Angiotensin II causes hypertension and cardiac hypertrophy through its receptors in the kidney. Proc. Natl. Acad. Sci. USA 2006, 103, 17985–17990. [Google Scholar] [CrossRef] [PubMed]
- Funder, J.W.; Carey, R.M.; Fardella, C.; Gomez-Sanchez, C.E.; Mantero, F.; Stowasser, M.; Young, W.F., Jr.; Montori, V.M.; Endocrine, S. Case detection, diagnosis, and treatment of patients with primary aldosteronism: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2008, 93, 3266–3281. [Google Scholar] [CrossRef] [PubMed]
- Fardella, C.E.; Mosso, L.; Gomez-Sanchez, C.; Cortes, P.; Soto, J.; Gomez, L.; Pinto, M.; Huete, A.; Oestreicher, E.; Foradori, A.; et al. Primary hyperaldosteronism in essential hypertensives: Prevalence, biochemical profile, and molecular biology. J. Clin. Endocrinol. Metab. 2000, 85, 1863–1867. [Google Scholar] [CrossRef] [PubMed]
- Fardella, C.E.; Mosso, L. Primary aldosteronism. Clin. Lab. 2002, 48, 181–190. [Google Scholar] [PubMed]
- Fardella, C.E.; Pinto, M.; Mosso, L.; Gomez-Sanchez, C.; Jalil, J.; Montero, J. Genetic study of patients with dexamethasone-suppressible aldosteronism without the chimeric CYP11B1/CYP11B2 gene. J. Clin. Endocrinol. Metab. 2001, 86, 4805–4807. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, C.A.; Campino, C.; Martinez-Aguayo, A.; Tichauer, J.E.; Bancalari, R.; Valdivia, C.; Trejo, P.; Aglony, M.; Baudrand, R.; Lagos, C.F.; et al. A new presentation of the chimeric CYP11B1/CYP11B2 gene with low prevalence of primary aldosteronism and atypical gene segregation pattern. Hypertension 2012, 59, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.; Fardella, C.; Mosso, L. Hipertensión arterial tratable con glucocorticoides: Comunicación de un caso. Rev. Méd. Chile 1977, 125, 1361–1365. [Google Scholar]
- Fardella, C.E.; Mosso, L.M.; Carvajal, C.A. Primary aldosteronism. Rev. Med. Chile 2008, 136, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Lalli, E.; Barhanin, J.; Zennaro, M.C.; Warth, R. Local control of aldosterone production and primary aldosteronism. Trends Endocrinol. Metab. 2016, 27, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.J.; Ruse, J.L.; Laidlaw, J.C. Hypertension, increased aldosterone secretion and low plasma renin activity relieved by dexamethasone. Can. Med. Assoc. J. 1966, 95, 1109–1119. [Google Scholar] [PubMed]
- Mulatero, P.; Tizzani, D.; Viola, A.; Bertello, C.; Monticone, S.; Mengozzi, G.; Schiavone, D.; Williams, T.A.; Einaudi, S.; La Grotta, A.; et al. Prevalence and characteristics of familial hyperaldosteronism: The patogen study (primary aldosteronism in torino-genetic forms). Hypertension 2011, 58, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Ogishima, T.; Shibata, H.; Shimada, H.; Mitani, F.; Suzuki, H.; Saruta, T.; Ishimura, Y. Aldosterone synthase cytochrome p-450 expressed in the adrenals of patients with primary aldosteronism. J. Biol. Chem. 1991, 266, 10731–10734. [Google Scholar] [PubMed]
- Gordon, R.D.; Stowasser, M.; Tunny, T.J.; Klemm, S.A.; Finn, W.L.; Krek, A.L. Clinical and pathological diversity of primary aldosteronism, including a new familial variety. Clin. Exp. Pharmacol. Physiol. 1991, 18, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, K.; Tomlins, S.A.; Kuick, R.; Cani, A.K.; Giordano, T.J.; Hovelson, D.H.; Liu, C.J.; Sanjanwala, A.R.; Edwards, M.A.; Gomez-Sanchez, C.E.; et al. Aldosterone-stimulating somatic gene mutations are common in normal adrenal glands. Proc. Natl. Acad. Sci. USA 2015, 112, E4591–E4599. [Google Scholar] [CrossRef] [PubMed]
- Geller, D.S.; Zhang, J.; Wisgerhof, M.V.; Shackleton, C.; Kashgarian, M.; Lifton, R.P. A novel form of human mendelian hypertension featuring nonglucocorticoid-remediable aldosteronism. J. Clin. Endocrinol. Metab. 2008, 93, 3117–3123. [Google Scholar] [CrossRef] [PubMed]
- Charmandari, E.; Sertedaki, A.; Kino, T.; Merakou, C.; Hoffman, D.A.; Hatch, M.M.; Hurt, D.E.; Lin, L.; Xekouki, P.; Stratakis, C.A.; et al. A novel point mutation in the KCNJ5 gene causing primary hyperaldosteronism and early-onset autosomal dominant hypertension. J. Clin. Endocrinol. Metab. 2012, 97, E1532–E1539. [Google Scholar] [CrossRef] [PubMed]
- Scholl, U.I.; Nelson-Williams, C.; Yue, P.; Grekin, R.; Wyatt, R.J.; Dillon, M.J.; Couch, R.; Hammer, L.K.; Harley, F.L.; Farhi, A.; et al. Hypertension with or without adrenal hyperplasia due to different inherited mutations in the potassium channel KCNJ5. Proc. Natl. Acad. Sci. USA 2012, 109, 2533–2538. [Google Scholar] [CrossRef] [PubMed]
- Scholl, U.I.; Goh, G.; Stolting, G.; de Oliveira, R.C.; Choi, M.; Overton, J.D.; Fonseca, A.L.; Korah, R.; Starker, L.F.; Kunstman, J.W.; et al. Somatic and germline cacna1d calcium channel mutations in aldosterone-producing adenomas and primary aldosteronism. Nat. Genet. 2013, 45, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.P.; Di Bello, V.; Ganzaroli, C.; Sacchetto, A.; Cesari, M.; Bertini, A.; Giorgi, D.; Scognamiglio, R.; Mariani, M.; Pessina, A.C. Excess aldosterone is associated with alterations of myocardial texture in primary aldosteronism. Hypertension 2002, 40, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Mosso, L.; Carvajal, C.; Gonzalez, A.; Barraza, A.; Avila, F.; Montero, J.; Huete, A.; Gederlini, A.; Fardella, C.E. Primary aldosteronism and hypertensive disease. Hypertension 2003, 42, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Stehr, C.B.; Mellado, R.; Ocaranza, M.P.; Carvajal, C.A.; Mosso, L.; Becerra, E.; Solis, M.; Garcia, L.; Lavandero, S.; Jalil, J.; et al. Increased levels of oxidative stress, subclinical inflammation, and myocardial fibrosis markers in primary aldosteronism patients. J. Hypertens. 2010, 28, 2120–2126. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Aguayo, A.; Aglony, M.; Campino, C.; Garcia, H.; Bancalari, R.; Bolte, L.; Avalos, C.; Loureiro, C.; Carvajal, C.A.; Avila, A.; et al. Aldosterone, plasma renin activity, and aldosterone/renin ratio in a normotensive healthy pediatric population. Hypertension 2010, 56, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, C.; Campino, C.; Martinez-Aguayo, A.; Godoy, I.; Aglony, M.; Bancalari, R.; Garcia, H.; Carvajal, C.A.; Fardella, C. Positive association between aldosterone-renin ratio and carotid intima-media thickness in hypertensive children. Clin. Endocrinol. (Oxf.) 2013, 78, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Rockey, D.C.; Bell, P.D.; Hill, J.A. Fibrosis—A common pathway to organ injury and failure. N. Engl. J. Med. 2015, 373. [Google Scholar] [CrossRef] [PubMed]
- Kagami, S.; Border, W.A.; Miller, D.E.; Noble, N.A. Angiotensin II stimulates extracellular matrix protein synthesis through induction of transforming growth factor-β expression in rat glomerular mesangial cells. J. Clin. Investig. 1994, 93, 2431–2437. [Google Scholar] [CrossRef] [PubMed]
- Arciniegas, E.; Sutton, A.B.; Allen, T.D.; Schor, A.M. Transforming growth factor β1 promotes the differentiation of endothelial cells into smooth muscle-like cells in vitro. J. Cell Sci. 1992, 103 Pt 2, 521–529. [Google Scholar] [PubMed]
- Zeisberg, E.M.; Tarnavski, O.; Zeisberg, M.; Dorfman, A.L.; McMullen, J.R.; Gustafsson, E.; Chandraker, A.; Yuan, X.; Pu, W.T.; Roberts, A.B.; et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat. Med. 2007, 13, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, J.; Zhang, J.Q.; Ramires, F.J. Local angiotensin II and transforming growth factor-β1 in renal fibrosis of rats. Hypertension 2000, 35, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- An, S.J.; Boyd, R.; Wang, Y.; Qiu, X.; Wang, H.D. Endothelin-1 expression in vascular adventitial fibroblasts. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H700–H708. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.D.; Chu, Y.F.; Li, X.D.; Gao, P.J. Angiotensin II induces expression of inflammatory mediators in vascular adventitial fibroblasts. Sheng Li Xue Bao 2015, 67, 603–610. [Google Scholar] [PubMed]
- McCarthy, C.G.; Wenceslau, C.F.; Goulopoulou, S.; Ogbi, S.; Baban, B.; Sullivan, J.C.; Matsumoto, T.; Webb, R.C. Circulating mitochondrial DNA and toll-like receptor 9 are associated with vascular dysfunction in spontaneously hypertensive rats. Cardiovasc. Res. 2015, 107, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Goulopoulou, S.; McCarthy, C.G.; Webb, R.C. Toll-like receptors in the vascular system: Sensing the dangers within. Pharmacol. Rev. 2016, 68, 142–167. [Google Scholar] [CrossRef] [PubMed]
- Eissler, R.; Schmaderer, C.; Rusai, K.; Kuhne, L.; Sollinger, D.; Lahmer, T.; Witzke, O.; Lutz, J.; Heemann, U.; Baumann, M. Hypertension augments cardiac toll-like receptor 4 expression and activity. Hypertens. Res. 2011, 34, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.; Li, Y.; Hua, F.; Ma, J.; Gao, X.; Kelley, J.; Zhao, A.; Haddad, G.E.; Williams, D.L.; William Browder, I.; et al. Reduced cardiac hypertrophy in toll-like receptor 4-deficient mice following pressure overload. Cardiovasc. Res. 2005, 68, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Hernanz, R.; Martinez-Revelles, S.; Palacios, R.; Martin, A.; Cachofeiro, V.; Aguado, A.; Garcia-Redondo, L.; Barrus, M.T.; de Batista, P.R.; Briones, A.M.; et al. Toll-like receptor 4 contributes to vascular remodelling and endothelial dysfunction in angiotensin II-induced hypertension. Br. J. Pharmacol. 2015, 172, 3159–3176. [Google Scholar] [CrossRef] [PubMed]
- Echem, C.; Bomfim, G.F.; Ceravolo, G.S.; Oliveira, M.A.; Santos-Eichler, R.A.; Bechara, L.R.; Veras, M.M.; Saldiva, P.H.; Ferreira, J.C.; Akamine, E.H.; et al. Anti-toll like receptor 4 (TLR4) therapy diminishes cardiac remodeling regardless of changes in blood pressure in spontaneously hypertensive rats (SHR). Int. J. Cardiol. 2015, 187, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, W.; Ao, X.; Dai, H.; Yuan, L.; Huang, X.; Zhou, Q. TAK-242, a toll-like receptor 4 antagonist, protects against aldosterone-induced cardiac and renal injury. PLoS ONE 2015, 10, e0142456. [Google Scholar] [CrossRef] [PubMed]
- Lubanski, M.S.; McCullough, P.A. Kidney’s role in hypertension. Minerva Cardioangiol. 2009, 57, 743–759. [Google Scholar] [PubMed]
- Calo, L.A.; Zaghetto, F.; Pagnin, E.; Davis, P.A.; de Mozzi, P.; Sartorato, P.; Martire, G.; Fiore, C.; Armanini, D. Effect of aldosterone and glycyrrhetinic acid on the protein expression of PAI-1 and p22(phox) in human mononuclear leukocytes. J. Clin. Endocrinol. Metab. 2004, 89, 1973–1976. [Google Scholar] [CrossRef] [PubMed]
- Walczak, C.; Gaignier, F.; Gilet, A.; Zou, F.; Thornton, S.N.; Ropars, A. Aldosterone increases VEGF-α production in human neutrophils through PI3K, ERK1/2 and p38 pathways. Biochim. Biophys. Acta 2011, 1813, 2125–2132. [Google Scholar] [CrossRef] [PubMed]
- Sekizawa, N.; Yoshimoto, T.; Hayakawa, E.; Suzuki, N.; Sugiyama, T.; Hirata, Y. Transcriptome analysis of aldosterone-regulated genes in human vascular endothelial cell lines stably expressing mineralocorticoid receptor. Mol. Cell. Endocrinol. 2011, 341, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Persson, F.; Frandsen, E.; Sugaya, T.; Hess, G.; Zdunek, D.; Shjoedt, K.J.; Parving, H.H.; Rossing, P. Spironolactone diminishes urinary albumin excretion in patients with type 1 diabetes and microalbuminuria: A randomized placebo-controlled crossover study. Diabet. Med. 2012, 29, e184–e190. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Schjoedt, K.J.; Rossing, K.; Persson, F.; Schalkwijk, C.G.; Stehouwer, C.D.; Parving, H.H.; Rossing, P. Levels of NT-proBNP, markers of low-grade inflammation, and endothelial dysfunction during spironolactone treatment in patients with diabetic kidney disease. J. Renin Angiotensin Aldosterone Syst. 2013, 14, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Mennuni, S.; Rubattu, S.; Pierelli, G.; Tocci, G.; Fofi, C.; Volpe, M. Hypertension and kidneys: Unraveling complex molecular mechanisms underlying hypertensive renal damage. J. Hum. Hypertens. 2014, 28, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Kurts, C.; Panzer, U.; Anders, H.J.; Rees, A.J. The immune system and kidney disease: Basic concepts and clinical implications. Nat. Rev. Immunol. 2013, 13, 738–753. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A.; Neilson, E.G. Chronic kidney disease progression. J. Am. Soc. Nephrol. 2006, 17, 2964–2966. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A. Overview of the cellular and molecular basis of kidney fibrosis. Kidney Int. Suppl. (2011) 2014, 4, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.C.; Chan, L.Y.; Tsang, A.W.; Liu, E.W.; Lam, M.F.; Tang, S.C.; Lai, K.N. Anti-macrophage migration inhibitory factor reduces transforming growth factor-β1 expression in experimental IgA nephropathy. Nephrol. Dial. Transplant. 2004, 19, 1976–1985. [Google Scholar] [CrossRef] [PubMed]
- Bruchfeld, A.; Wendt, M.; Miller, E.J. Macrophage migration inhibitory factor in clinical kidney disease. Front. Immunol. 2016, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Sussman, C.R. TGF-β, Notch, and HGF weave a tangled web of kidney repair. Am. J. Physiol. Ren. Physiol. 2016, 310, F744–F745. [Google Scholar] [CrossRef] [PubMed]
- Pueyo, M.E.; Gonzalez, W.; Nicoletti, A.; Savoie, F.; Arnal, J.F.; Michel, J.B. Angiotensin II stimulates endothelial vascular cell adhesion molecule-1 via nuclear factor-κb activation induced by intracellular oxidative stress. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Sowers, J.R. Metabolic risk factors and renal disease. Kidney Int. 2007, 71, 719–720. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.A.; Whaley-Connell, A.; Habibi, J.; Wei, Y.; Lastra, G.; Manrique, C.; Stas, S.; Sowers, J.R. Renin-angiotensin-aldosterone system and oxidative stress in cardiovascular insulin resistance. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2009–H2023. [Google Scholar] [CrossRef] [PubMed]
- Epstein, M. Aldosterone blockade: An emerging strategy for abrogating progressive renal disease. Am. J. Med. 2006, 119, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Nagase, M.; Yoshida, S.; Shibata, S.; Nagase, T.; Gotoda, T.; Ando, K.; Fujita, T. Enhanced aldosterone signaling in the early nephropathy of rats with metabolic syndrome: Possible contribution of fat-derived factors. J. Am. Soc. Nephrol. 2006, 17, 3438–3446. [Google Scholar] [CrossRef] [PubMed]
- Bomback, A.S.; Klemmer, P.J. Renal injury in extreme obesity: The important role of aldosterone. Kidney Int. 2008, 74, 1216, author reply 1216–1217. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R.; Rudolph, A.E.; Frierdich, G.E.; Nachowiak, D.A.; Kekec, B.K.; Blomme, E.A.; McMahon, E.G.; Delyani, J.A. Aldosterone induces a vascular inflammatory phenotype in the rat heart. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1802–H1810. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Andres, N.; Martin-Fernandez, B.; Rossignol, P.; Zannad, F.; Lahera, V.; Fortuno, M.A.; Cachofeiro, V.; Diez, J. A role for cardiotrophin-1 in myocardial remodeling induced by aldosterone. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2372–H2382. [Google Scholar] [CrossRef] [PubMed]
- Dartsch, T.; Fischer, R.; Gapelyuk, A.; Weiergraeber, M.; Ladage, D.; Schneider, T.; Schirdewan, A.; Reuter, H.; Mueller-Ehmsen, J.; Zobel, C. Aldosterone induces electrical remodeling independent of hypertension. Int. J. Cardiol. 2013, 164, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Han, J.S.; Choi, B.S.; Yang, C.W.; Kim, Y.S. Aldosterone-induced TGF-β1 expression is regulated by mitogen-activated protein kinases and activator protein-1 in mesangial cells. J. Korean Med. Sci. 2009, 24 (Suppl. 1), S195–S203. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Rudolph, A.E.; Bond, B.R.; Rocha, R.; Blomme, E.A.; Goellner, J.J.; Funder, J.W.; McMahon, E.G. Transgenic model of aldosterone-driven cardiac hypertrophy and heart failure. Circ. Res. 2003, 93, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Tsybouleva, N.; Zhang, L.; Chen, S.; Patel, R.; Lutucuta, S.; Nemoto, S.; DeFreitas, G.; Entman, M.; Carabello, B.A.; Roberts, R.; et al. Aldosterone, through novel signaling proteins, is a fundamental molecular bridge between the genetic defect and the cardiac phenotype of hypertrophic cardiomyopathy. Circulation 2004, 109, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Lemarie, C.A.; Simeone, S.M.; Nikonova, A.; Ebrahimian, T.; Deschenes, M.E.; Coffman, T.M.; Paradis, P.; Schiffrin, E.L. Aldosterone-induced activation of signaling pathways requires activity of angiotensin type 1a receptors. Circ. Res. 2009, 105, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Kobara, M.; Abe, M.; Tanaka, N.; Gouda, E.; Toba, H.; Yamada, H.; Tatsumi, T.; Nakata, T.; Matsubara, H. Aldosterone nongenomically produces NADPH oxidase-dependent reactive oxygen species and induces myocyte apoptosis. Hypertens. Res. 2008, 31, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Keidar, S.; Kaplan, M.; Pavlotzky, E.; Coleman, R.; Hayek, T.; Hamoud, S.; Aviram, M. Aldosterone administration to mice stimulates macrophage NADPH oxidase and increases atherosclerosis development: A possible role for angiotensin-converting enzyme and the receptors for angiotensin II and aldosterone. Circulation 2004, 109, 2213–2220. [Google Scholar] [CrossRef] [PubMed]
- Iwashima, F.; Yoshimoto, T.; Minami, I.; Sakurada, M.; Hirono, Y.; Hirata, Y. Aldosterone induces superoxide generation via Rac1 activation in endothelial cells. Endocrinology 2008, 149, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Hirono, Y.; Yoshimoto, T.; Suzuki, N.; Sugiyama, T.; Sakurada, M.; Takai, S.; Kobayashi, N.; Shichiri, M.; Hirata, Y. Angiotensin II receptor type 1-mediated vascular oxidative stress and proinflammatory gene expression in aldosterone-induced hypertension: The possible role of local renin-angiotensin system. Endocrinology 2007, 148, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Johar, S.; Cave, A.C.; Narayanapanicker, A.; Grieve, D.J.; Shah, A.M. Aldosterone mediates angiotensin II-induced interstitial cardiac fibrosis via a Nox2-containing NADPH oxidase. FASEB J. 2006, 20, 1546–1548. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.Y.; Bloem, L.J.; Pratt, J.H. Aldosterone inhibits inducible nitric oxide synthase in neonatal rat cardiomyocytes. Endocrinology 2003, 144, 1712–1717. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Lam, A.; Abraham, J.A.; Schreiner, G.F.; Joly, A.H. CTGF expression is induced by TGF-β in cardiac fibroblasts and cardiac myocytes: A potential role in heart fibrosis. J. Mol. Cell. Cardiol. 2000, 32, 1805–1819. [Google Scholar] [CrossRef] [PubMed]
- Martin-Fernandez, B.; de las Heras, N.; Miana, M.; Ballesteros, S.; Delgado, C.; Song, S.; Hintze, T.; Cachofeiro, V.; Lahera, V. Structural, functional, and molecular alterations produced by aldosterone plus salt in rat heart: Association with enhanced serum and glucocorticoid-regulated kinase-1 expression. J. Cardiovasc. Pharmacol. 2011, 57, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Habibi, J.; DeMarco, V.G.; Ma, L.; Pulakat, L.; Rainey, W.E.; Whaley-Connell, A.T.; Sowers, J.R. Mineralocorticoid receptor blockade improves diastolic function independent of blood pressure reduction in a transgenic model of raas overexpression. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1484–H1491. [Google Scholar] [CrossRef] [PubMed]
- Kagiyama, S.; Matsumura, K.; Goto, K.; Otsubo, T.; Iida, M. Role of rho kinase and oxidative stress in cardiac fibrosis induced by aldosterone and salt in angiotensin type 1a receptor knockout mice. Regul. Pept. 2010, 160, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Taira, M.; Toba, H.; Murakami, M.; Iga, I.; Serizawa, R.; Murata, S.; Kobara, M.; Nakata, T. Spironolactone exhibits direct renoprotective effects and inhibits renal renin-angiotensin-aldosterone system in diabetic rats. Eur. J. Pharmacol. 2008, 589, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Lacolley, P.; Regnault, V.; Nicoletti, A.; Li, Z.; Michel, J.B. The vascular smooth muscle cell in arterial pathology: A cell that can take on multiple roles. Cardiovasc. Res. 2012, 95, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Lombes, M.; Farman, N.; Bonvalet, J.P.; Zennaro, M.C. Identification and role of aldosterone receptors in the cardiovascular system. Ann. Endocrinol. (Paris) 2000, 61, 41–46. [Google Scholar] [PubMed]
- Kohan, D.E.; Rossi, N.F.; Inscho, E.W.; Pollock, D.M. Regulation of blood pressure and salt homeostasis by endothelin. Physiol. Rev. 2011, 91, 1–77. [Google Scholar] [CrossRef] [PubMed]
- Lynch, I.J.; Welch, A.K.; Kohan, D.E.; Cain, B.D.; Wingo, C.S. Endothelin-1 inhibits sodium reabsorption by ETA and ETB receptors in the mouse cortical collecting duct. Am. J. Physiol. Ren. Physiol. 2013, 305, F568–F573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Saito, Y.; Naya, N.; Imagawa, K.; Somekawa, S.; Kawata, H.; Takeda, Y.; Uemura, S.; Kishimoto, I.; Nakao, K. The specific mineralocorticoid receptor blocker eplerenone attenuates left ventricular remodeling in mice lacking the gene encoding guanylyl cyclase-A. Hypertens. Res. 2008, 31, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Tomita, M.; Yoshimi, H.; Kuramochi, M.; Ito, K.; Ikeda, M. Effect of synthetic human atrial natriuretic peptide on aldosterone secretion by dispersed aldosterone-producing adenoma cells in vitro. J. Clin. Endocrinol. Metab. 1985, 61, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Yoshimura, M.; Nakamura, S.; Nakayama, M.; Shimasaki, Y.; Harada, E.; Mizuno, Y.; Yamamuro, M.; Harada, M.; Saito, Y.; et al. Inhibitory effect of natriuretic peptides on aldosterone synthase gene expression in cultured neonatal rat cardiocytes. Circulation 2003, 107, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Hiroki, J.; Shimokawa, H.; Higashi, M.; Morikawa, K.; Kandabashi, T.; Kawamura, N.; Kubota, T.; Ichiki, T.; Amano, M.; Kaibuchi, K.; et al. Inflammatory stimuli upregulate Rho-kinase in human coronary vascular smooth muscle cells. J. Mol. Cell. Cardiol. 2004, 37, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, H.; Satoh, K. 2015 ATVB plenary lecture: Translational research on Rho-kinase in cardiovascular medicine. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1756–1769. [Google Scholar] [CrossRef] [PubMed]
- Vahebi, S.; Kobayashi, T.; Warren, C.M.; de Tombe, P.P.; Solaro, R.J. Functional effects of Rho-kinase-dependent phosphorylation of specific sites on cardiac troponin. Circ. Res. 2005, 96, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Li, H. Therapeutic effect of enhancing endothelial nitric oxide synthase (ENOS) expression and preventing enos uncoupling. Br. J. Pharmacol. 2011, 164, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Bakris, G.L.; Agarwal, R.; Chan, J.C.; Cooper, M.E.; Gansevoort, R.T.; Haller, H.; Remuzzi, G.; Rossing, P.; Schmieder, R.E.; Nowack, C.; et al. Effect of finerenone on albuminuria in patients with diabetic nephropathy: A randomized clinical trial. JAMA 2015, 314, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Kolkhof, P.; Delbeck, M.; Kretschmer, A.; Steinke, W.; Hartmann, E.; Barfacker, L.; Eitner, F.; Albrecht-Kupper, B.; Schafer, S. Finerenone, a novel selective nonsteroidal mineralocorticoid receptor antagonist protects from rat cardiorenal injury. J. Cardiovasc. Pharmacol. 2014, 64, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Anker, S.D.; Bohm, M.; Gheorghiade, M.; Kober, L.; Krum, H.; Maggioni, A.P.; Ponikowski, P.; Voors, A.A.; Zannad, F.; et al. Rationale and design of mineralocorticoid receptor antagonist tolerability study-heart failure (ARTS-HF): A randomized study of finerenone vs. Eplerenone in patients who have worsening chronic heart failure with diabetes and/or chronic kidney disease. Eur. J. Heart Fail. 2015, 17, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Naito, Y.; Weng, H.; Ma, X.; Endo, K.; Kito, N.; Yanagawa, N.; Yu, Y.; Li, J.; Iwai, N. Renoprotective mechanisms of pirfenidone in hypertension-induced renal injury: Through anti-fibrotic and anti-oxidative stress pathways. Biomed. Res. 2013, 34, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Albrektsen, T.; Frederiksen, K.S.; Holmes, W.E.; Boel, E.; Taylor, K.; Fleckner, J. Novel genes regulated by the insulin sensitizer rosiglitazone during adipocyte differentiation. Diabetes 2002, 51, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Burson, J.M.; Aguilera, G.; Gross, K.W.; Sigmund, C.D. Differential expression of angiotensin receptor 1a and 1b in mouse. Am. J. Physiol. 1994, 267, E260–E267. [Google Scholar] [PubMed]
- Crandall, D.L.; Herzlinger, H.E.; Saunders, B.D.; Armellino, D.C.; Kral, J.G. Distribution of angiotensin II receptors in rat and human adipocytes. J. Lipid Res. 1994, 35, 1378–1385. [Google Scholar] [PubMed]
- Engeli, S.; Gorzelniak, K.; Kreutz, R.; Runkel, N.; Distler, A.; Sharma, A.M. Co-expression of renin-angiotensin system genes in human adipose tissue. J. Hypertens. 1999, 17, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Whelan, J.; Claycombe, K.; Reath, D.B.; Moustaid-Moussa, N. Angiotensin II increases leptin secretion by 3T3-L1 and human adipocytes via a prostaglandin-independent mechanism. J. Nutr. 2002, 132, 1135–1140. [Google Scholar] [PubMed]
- Skurk, T.; Lee, Y.M.; Hauner, H. Angiotensin II and its metabolites stimulate PAI-1 protein release from human adipocytes in primary culture. Hypertension 2001, 37, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.M.; Janke, J.; Gorzelniak, K.; Engeli, S.; Luft, F.C. Angiotensin blockade prevents type 2 diabetes by formation of fat cells. Hypertension 2002, 40, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Goodfriend, T.L.; Ball, D.L.; Egan, B.M.; Campbell, W.B.; Nithipatikom, K. Epoxy-keto derivative of linoleic acid stimulates aldosterone secretion. Hypertension 2004, 43, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Ehrhart-Bornstein, M.; Lamounier-Zepter, V.; Schraven, A.; Langenbach, J.; Willenberg, H.S.; Barthel, A.; Hauner, H.; McCann, S.M.; Scherbaum, W.A.; Bornstein, S.R. Human adipocytes secrete mineralocorticoid-releasing factors. Proc. Natl. Acad. Sci. USA 2003, 100, 14211–14216. [Google Scholar] [CrossRef] [PubMed]
- Engeli, S.; Bohnke, J.; Gorzelniak, K.; Janke, J.; Schling, P.; Bader, M.; Luft, F.C.; Sharma, A.M. Weight loss and the renin-angiotensin-aldosterone system. Hypertension 2005, 45, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Vedani, P.; Manunta, P.; Pizzocri, P.; Marchi, M.; Paganelli, M.; Folli, F.; Pontiroli, A.E. Effect of weight loss through laparoscopic gastric banding on blood pressure, plasma renin activity and aldosterone levels in morbid obesity. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Tuck, M.L.; Sowers, J.; Dornfeld, L.; Kledzik, G.; Maxwell, M. The effect of weight reduction on blood pressure, plasma renin activity, and plasma aldosterone levels in obese patients. N. Engl. J. Med. 1981, 304, 930–933. [Google Scholar] [CrossRef] [PubMed]
- Northcott, C.A.; Fink, G.D.; Garver, H.; Haywood, J.R.; Laimon-Thomson, E.L.; McClain, J.L.; Pires, P.W.; Rainey, W.E.; Rigsby, C.S.; Dorrance, A.M. The development of hypertension and hyperaldosteronism in a rodent model of life-long obesity. Endocrinology 2012, 153, 1764–1773. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Durango, N.; Fuentes, C.A.; Castillo, A.E.; González-Gómez, L.M.; Vecchiola, A.; Fardella, C.E.; Kalergis, A.M. Role of the Renin-Angiotensin-Aldosterone System beyond Blood Pressure Regulation: Molecular and Cellular Mechanisms Involved in End-Organ Damage during Arterial Hypertension. Int. J. Mol. Sci. 2016, 17, 797. https://doi.org/10.3390/ijms17070797
Muñoz-Durango N, Fuentes CA, Castillo AE, González-Gómez LM, Vecchiola A, Fardella CE, Kalergis AM. Role of the Renin-Angiotensin-Aldosterone System beyond Blood Pressure Regulation: Molecular and Cellular Mechanisms Involved in End-Organ Damage during Arterial Hypertension. International Journal of Molecular Sciences. 2016; 17(7):797. https://doi.org/10.3390/ijms17070797
Chicago/Turabian StyleMuñoz-Durango, Natalia, Cristóbal A. Fuentes, Andrés E. Castillo, Luis Martín González-Gómez, Andrea Vecchiola, Carlos E. Fardella, and Alexis M. Kalergis. 2016. "Role of the Renin-Angiotensin-Aldosterone System beyond Blood Pressure Regulation: Molecular and Cellular Mechanisms Involved in End-Organ Damage during Arterial Hypertension" International Journal of Molecular Sciences 17, no. 7: 797. https://doi.org/10.3390/ijms17070797
APA StyleMuñoz-Durango, N., Fuentes, C. A., Castillo, A. E., González-Gómez, L. M., Vecchiola, A., Fardella, C. E., & Kalergis, A. M. (2016). Role of the Renin-Angiotensin-Aldosterone System beyond Blood Pressure Regulation: Molecular and Cellular Mechanisms Involved in End-Organ Damage during Arterial Hypertension. International Journal of Molecular Sciences, 17(7), 797. https://doi.org/10.3390/ijms17070797