
Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas?

Abstract
:1. Introduction
2. Results
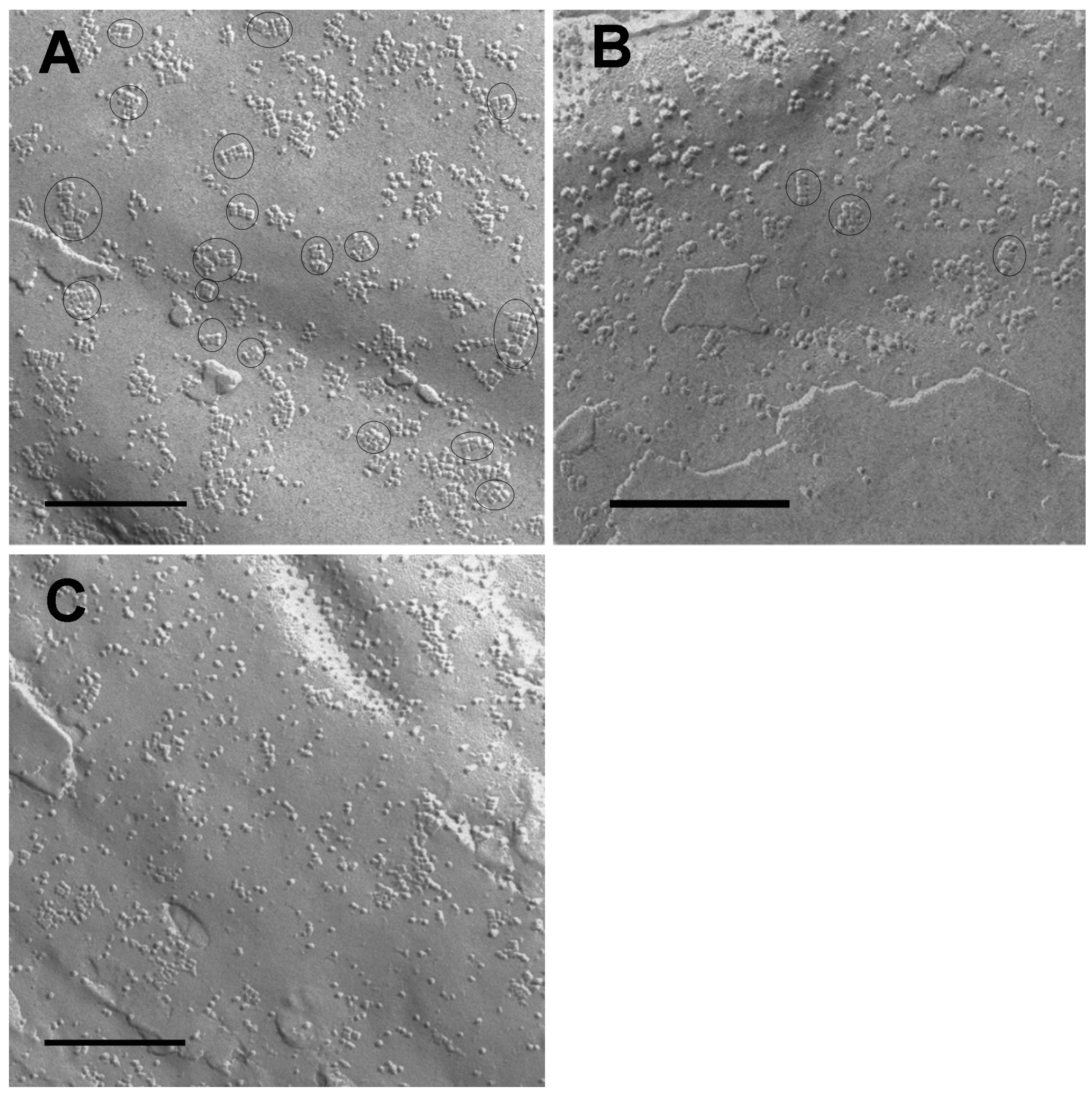
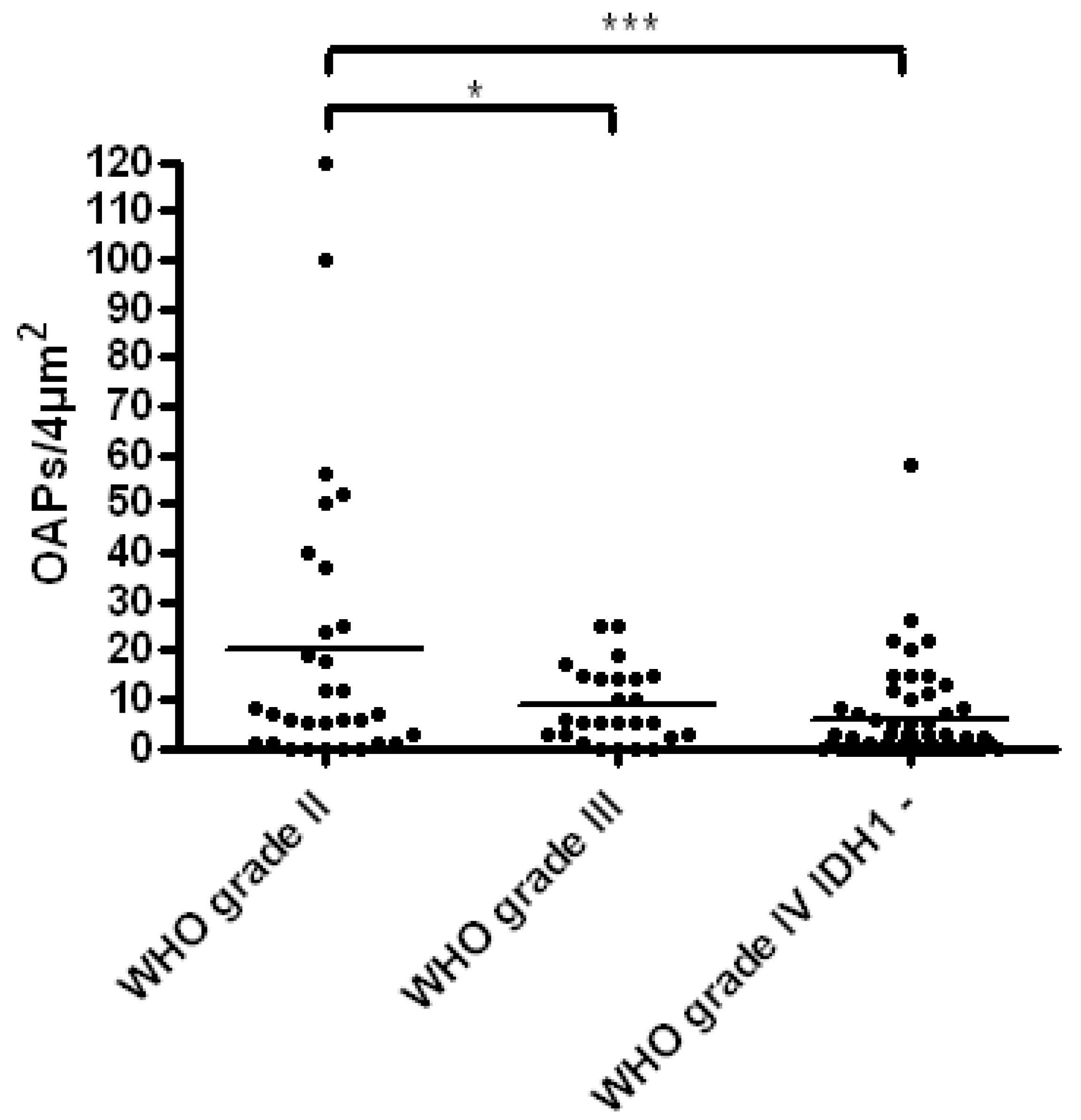
2.1. Determination of Orthogonal Arrays of Particles (OAPs) from Low-Grade to High-Grade Malignancies
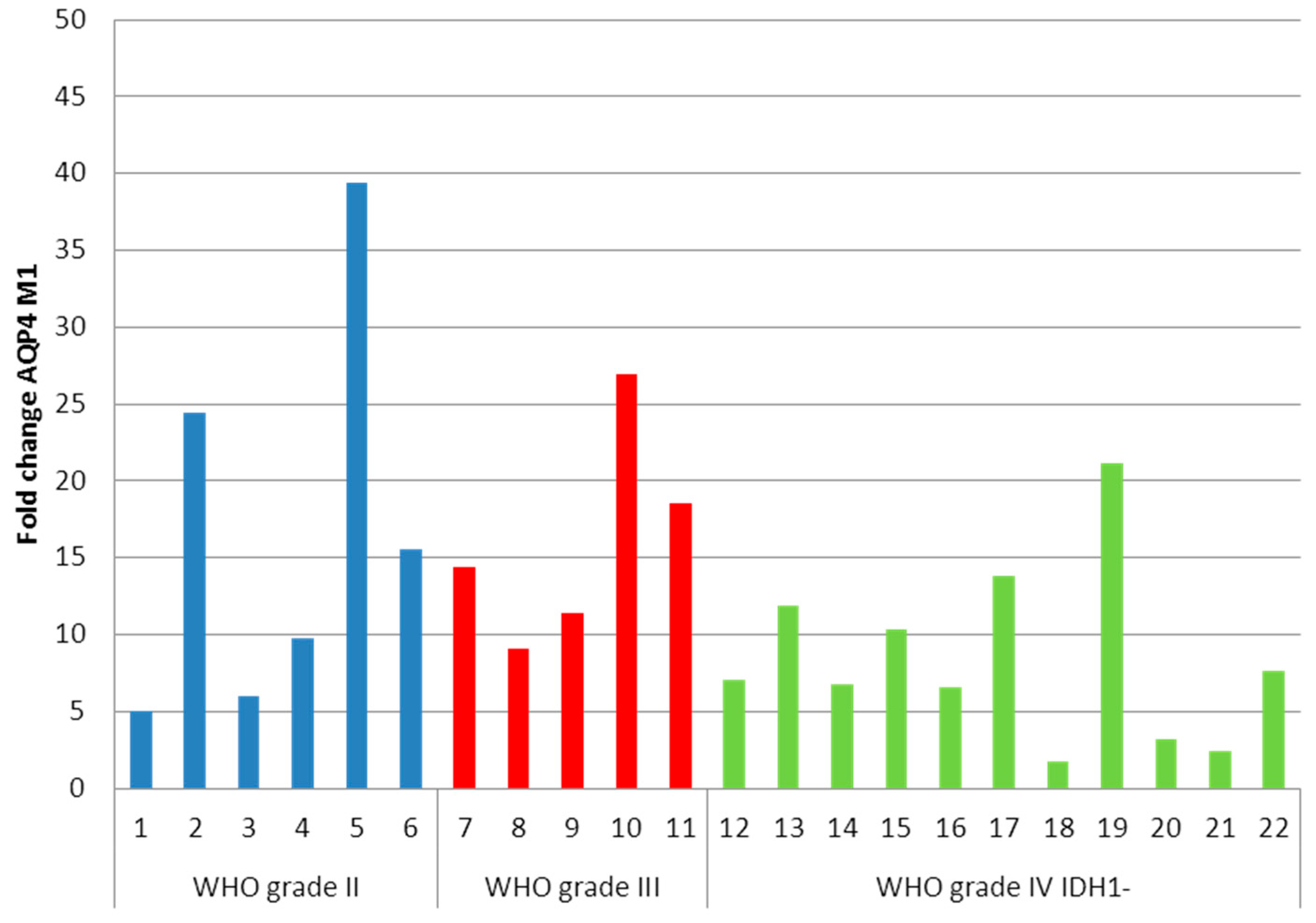
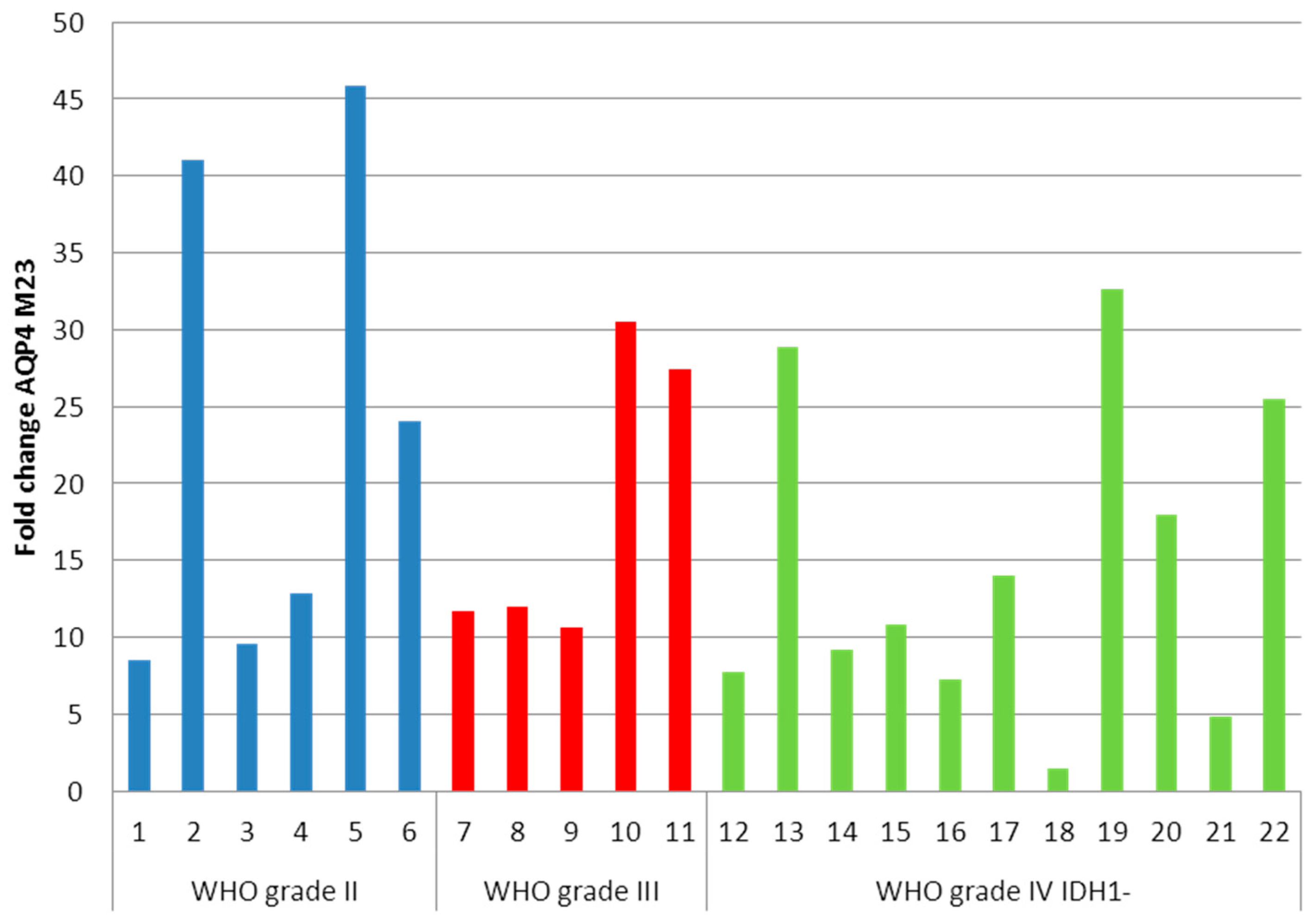
2.2. Determination of Aquaporin 4 (AQP4)-M1 and -M23 mRNA in Low-Grade and High-Grade Astrocytomas
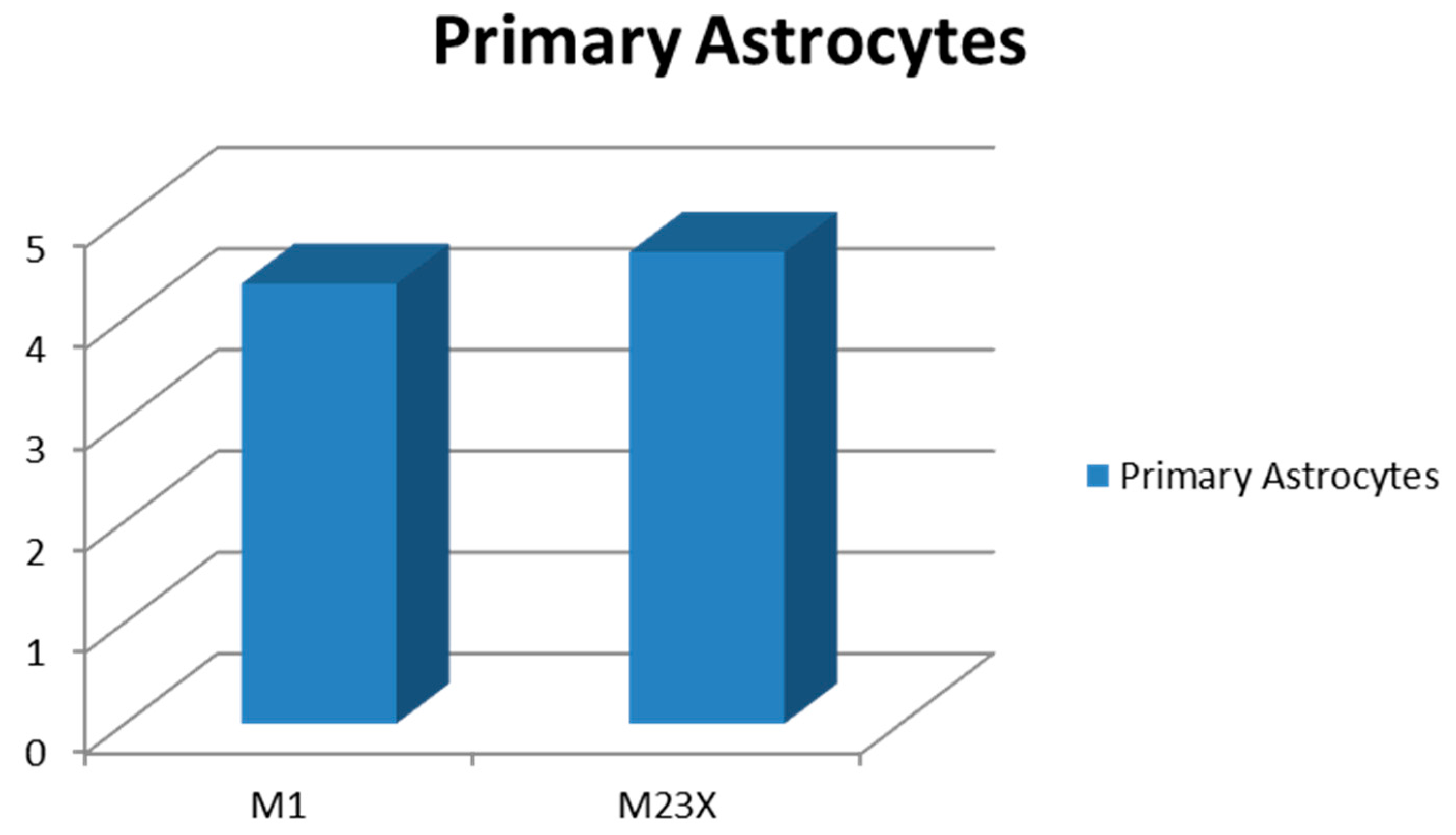
2.3. Determination of AQP4-M1 and -M23 mRNA in Cultures of Normal Astrocytes and Glioma Cells
3. Discussion
4. Materials and Methods
4.1. Tissue
4.2. Cell Culture
4.3. Freeze Fracture Technique and Determination of OAP Densities
4.4. RNA Extraction
4.5. cDNA Synthesis
4.6. Quantitative Real Time-PCR (qRT-PCR)
4.7. Data Analysis and Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rash, J.E.; Yasumura, T.; Hudson, C.S.; Agre, P.; Nielsen, S. Direct immunogold labeling of aquaporin-4 in square arrays of astrocyte and ependymocyte plasma membranes in rat brain and spinal cord. Proc. Natl. Acad. Sci. USA 1998, 95, 11981–11986. [Google Scholar] [CrossRef] [PubMed]
- Wolburg, H.; Wolburg-Buchholz, K.; Fallier-Becker, P.; Noell, S.; Mack, A.F. Structure and functions of aquaporin-4-based orthogonal arrays of particles. Int. Rev. Cell Mol. Biol. 2011, 287, 1–41. [Google Scholar] [PubMed]
- Nicchia, G.P.; Rossi, A.; Mola, M.G.; Pisani, F.; Stigliano, C.; Basco, D.; Mastrototaro, M.; Svelto, M.; Frigeri, A. Higher order structure of aquaporin-4. Neuroscience 2010, 168, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Rohlmann, A.; Gocht, A.; Wolburg, H. Reactive astrocytes in myelin-deficient rat optic-nerve reveal an altered distribution of orthogonal arrays of particles (OAP). Glia 1992, 5, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Saadoun, S.; Papadopoulos, M.C.; Davies, D.C.; Krishna, S.; Bell, B.A. Aquaporin-4 expression is increased in oedematous human brain tumours. J. Neurol. Neurosurg. Psychiatry 2002, 72, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Wolburg-Buchholz, K.; Mack, A.F.; Ritz, R.; Tatagiba, M.; Beschorner, R.; Wolburg, H.; Fallier-Becker, P. Dynamics of expression patterns of AQP4, dystroglycan, agrin and matrix metalloproteinases in human glioblastoma. Cell Tissue Res. 2012, 347, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Neely, J.D.; Christensen, B.M.; Nielsen, S.; Agre, P. Heterotetrameric composition of aquaporin-4 water channels. Biochemistry 1999, 38, 11156–11163. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Jin, B.J.; Ratelade, J.; Verkman, A.S. Aggregation state determines the localization and function of M1- and M23-aquaporin-4 in astrocytes. J. Cell Biol. 2014, 204, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Furman, C.S.; Gorelick-Feldman, D.A.; Davidson, K.G.; Yasumura, T.; Neely, J.D.; Agre, P.; Rash, J.E. Aquaporin-4 square array assembly: Opposing actions of M1 and M23 isoforms. Proc. Natl. Acad. Sci. USA 2003, 100, 13609–13614. [Google Scholar] [CrossRef] [PubMed]
- Fryer, J.P.; Lennon, V.A.; Pittock, S.J.; Jenkins, S.M.; Fallier-Becker, P.; Clardy, S.L.; Horta, E.; Jedynak, E.A.; Lucchinetti, C.F.; Shuster, E.A.; et al. Aqp4 autoantibody assay performance in clinical laboratory service. Neuroimmunol. Neuroinflamm. 2014, 1, 1e11. [Google Scholar] [CrossRef] [PubMed]
- Warth, A.; Simon, P.; Capper, D.; Goeppert, B.; Tabatabai, G.; Herzog, H.; Dietz, K.; Stubenvoll, F.; Ajaaj, R.; Becker, R.; et al. Expression pattern of the water channel aquaporin-4 in human gliomas is associated with blood-brain barrier disturbance but not with patient survival. J. Neurosci. Res. 2007, 85, 1336–1346. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.J.; Rossi, A.; Verkman, A.S. Model of aquaporin-4 supramolecular assembly in orthogonal arrays based on heterotetrameric association of M1-M23 isoforms. Biophys. J. 2011, 100, 2936–2945. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Ritz, R.; Wolburg-Buchholz, K.; Wolburg, H.; Fallier-Becker, P. An allograft glioma model reveals the dependence of aquaporin-4 expression on the brain microenvironment. PLoS ONE 2012, 7, e36555. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Fallier-Becker, P.; Deutsch, U.; Mack, A.F.; Wolburg, H. Agrin defines polarized distribution of orthogonal arrays of particles in astrocytes. Cell Tissue Res. 2009, 337, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Wolburg-Buchholz, K.; Mack, A.F.; Beedle, A.M.; Satz, J.S.; Campbell, K.P.; Wolburg, H.; Fallier-Becker, P. Evidence for a role of dystroglycan regulating the membrane architecture of astroglial endfeet. Eur. J. Neurosci. 2011, 33, 2179–2186. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Fallier-Becker, P.; Mack, A.F.; Hoffmeister, M.; Beschorner, R.; Ritz, R. Water channels aquaporin 4 and -1 expression in subependymoma depends on the localization of the tumors. PLoS ONE 2015, 10, e0131367. [Google Scholar] [CrossRef] [PubMed]
- Sole, S.; Petegnief, V.; Gorina, R.; Chamorro, A.; Planas, A.M. Activation of matrix metalloproteinase-3 and agrin cleavage in cerebral ischemia/reperfusion. J. Neuropathol. Exp. Neurol. 2004, 63, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Anderson, P.; Durbeej, M.; van Rooijen, N.; Ivars, F.; Opdenakker, G.; Sorokin, L.M. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. J. Exp. Med. 2006, 203, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Fallier-Becker, P.; Beyer, C.; Kroger, S.; Mack, A.F.; Wolburg, H. Effects of agrin on the expression and distribution of the water channel protein aquaporin-4 and volume regulation in cultured astrocytes. Eur. J. Neurosci. 2007, 26, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Fallier-Becker, P.; Sperveslage, J.; Wolburg, H.; Noell, S. The impact of agrin on the formation of orthogonal arrays of particles in cultured astrocytes from wild-type and agrin-null mice. Brain Res. 2011, 1367, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.M.; Tajima, M.; Verkman, A.S. Live-cell imaging of aquaporin-4 diffusion and interactions in orthogonal arrays of particles. Neuroscience 2010, 168, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Tani, K.; Mitsuma, T.; Hiroaki, Y.; Kamegawa, A.; Nishikawa, K.; Tanimura, Y.; Fujiyoshi, Y. Mechanism of aquaporin-4’s fast and highly selective water conduction and proton exclusion. J. Mol. Biol. 2009, 389, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Fenton, R.A.; Moeller, H.B.; Zelenina, M.; Snaebjornsson, M.T.; Holen, T.; MacAulay, N. Differential water permeability and regulation of three aquaporin 4 isoforms. Cell. Mol. Life Sci. 2010, 67, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Beyer, C.; Raab, H. Nongenomic effects of oestrogen: Embryonic mouse midbrain neurones respond with a rapid release of calcium from intracellular stores. Eur. J. Neurosci. 1998, 10, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, T.; Karolczak, M.; Beyer, C. Estradiol stimulates GDNF expression in developing hypothalamic neurons. Endocrinology 2002, 143, 3175–3178. [Google Scholar] [CrossRef] [PubMed]
- Zelenin, S.; Gunnarson, E.; Alikina, T.; Bondar, A.; Aperia, A. Identification of a new form of AQP4 mRNA that is developmentally expressed in mouse brain. Pediatr. Res. 2000, 48, 335–339. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Expression/Protein Complex | Healthy Brain | Glioblastoma |
|---|---|---|
| Agrin | + | − |
| Dystroglycan | + | − |
| Active MMP3 | − | + |
| Active MMP2/9 | − | + |
| OAPs | + | − |
| Patient Number | Astrocytoma WHO Grade | IDH1 |
|---|---|---|
| 1 | II | IDH1 (R132H) mutation (IDH1+) |
| 2 | II | IDH1 R132H) mutation (IDH1+) |
| 3 | II | IDH1 (R132H) mutation (IDH1+) |
| 4 | II | IDH1 (R132H) mutation (IDH1+) |
| 5 | II | IDH1 (R132H) mutation (IDH1+) |
| 6 | II | IDH1 (R132H) mutation (IDH1+) |
| 7 | III | IDH1 (R132H) mutation (IDH1+) |
| 8 | III | IDH1 (R132H) mutation (IDH1+) |
| 9 | III | IDH1 (R132H) mutation (IDH1+) |
| 10 | III | IDH1 (R132H) mutation (IDH1+) |
| 11 | III | IDH1 (R132H) mutation (IDH1+) |
| 12 | IV | IDH1 wildtype (IDH1−) |
| 13 | IV | IDH1 wildtype (IDH1−) |
| 14 | IV | IDH1 wildtype (IDH1−) |
| 15 | IV | IDH1 wildtype (IDH1−) |
| 16 | IV | IDH1 wildtype (IDH1−) |
| 17 | IV | IDH1 wildtype (IDH1−) |
| 18 | IV | IDH1 wildtype (IDH1−) |
| 19 | IV | IDH1 wildtype (IDH1−) |
| 20 | IV | IDH1 wildtype (IDH1−) |
| 21 | IV | IDH1 wildtype (IDH1−) |
| 22 | IV | IDH1 wildtype (IDH1−) |
| Primer | Sequence | Product Size |
|---|---|---|
| HPRT1 Ex6 for HPRT1 Ex7 rev | TGACACTGGCAAAACAATGC TTCGTGGGGTCCTTTTCACC | 101 bp |
| AQP4 M1 Ex1 for AQP4 M1 Ex2 rev | GGGGAAGGCATGAGTGACAG AAAGCTTGAGTCCAGACCCC | 110 bp |
| AQP4 M23 Ex1 for AQP4 M23 Ex2 rev | TCTCTTTTCAGTAAGTGTGGACCT CATGGCCAGAAATTCCGCTG | 114 bp |
| Primer | Sequence | Product Size |
|---|---|---|
| HPRT Ex6 Mus for HPRT Ex7 Mus rev | CAAACTTTGCTTTCCCTGGT GGCCTGTATCCAACACTTCG | 91 bp |
| AQP4 M1 Mus Ex1 for AQP4 M1 Mus Ex2 rev | AGGGAAGGCATGAGTGACAG GACTCCTTTGAAAGCCACCA | 96 bp |
| AQP4 M23X Mus for AQP4 M23X Mus rev | TATGGTTCACGGGTTTGGAT CCCTTTGTCACCTGCTCATT | 139 bp |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fallier-Becker, P.; Nieser, M.; Wenzel, U.; Ritz, R.; Noell, S. Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas? Int. J. Mol. Sci. 2016, 17, 1230. https://doi.org/10.3390/ijms17081230
Fallier-Becker P, Nieser M, Wenzel U, Ritz R, Noell S. Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas? International Journal of Molecular Sciences. 2016; 17(8):1230. https://doi.org/10.3390/ijms17081230
Chicago/Turabian StyleFallier-Becker, Petra, Maike Nieser, Ulrike Wenzel, Rainer Ritz, and Susan Noell. 2016. "Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas?" International Journal of Molecular Sciences 17, no. 8: 1230. https://doi.org/10.3390/ijms17081230
APA StyleFallier-Becker, P., Nieser, M., Wenzel, U., Ritz, R., & Noell, S. (2016). Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas? International Journal of Molecular Sciences, 17(8), 1230. https://doi.org/10.3390/ijms17081230