
Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
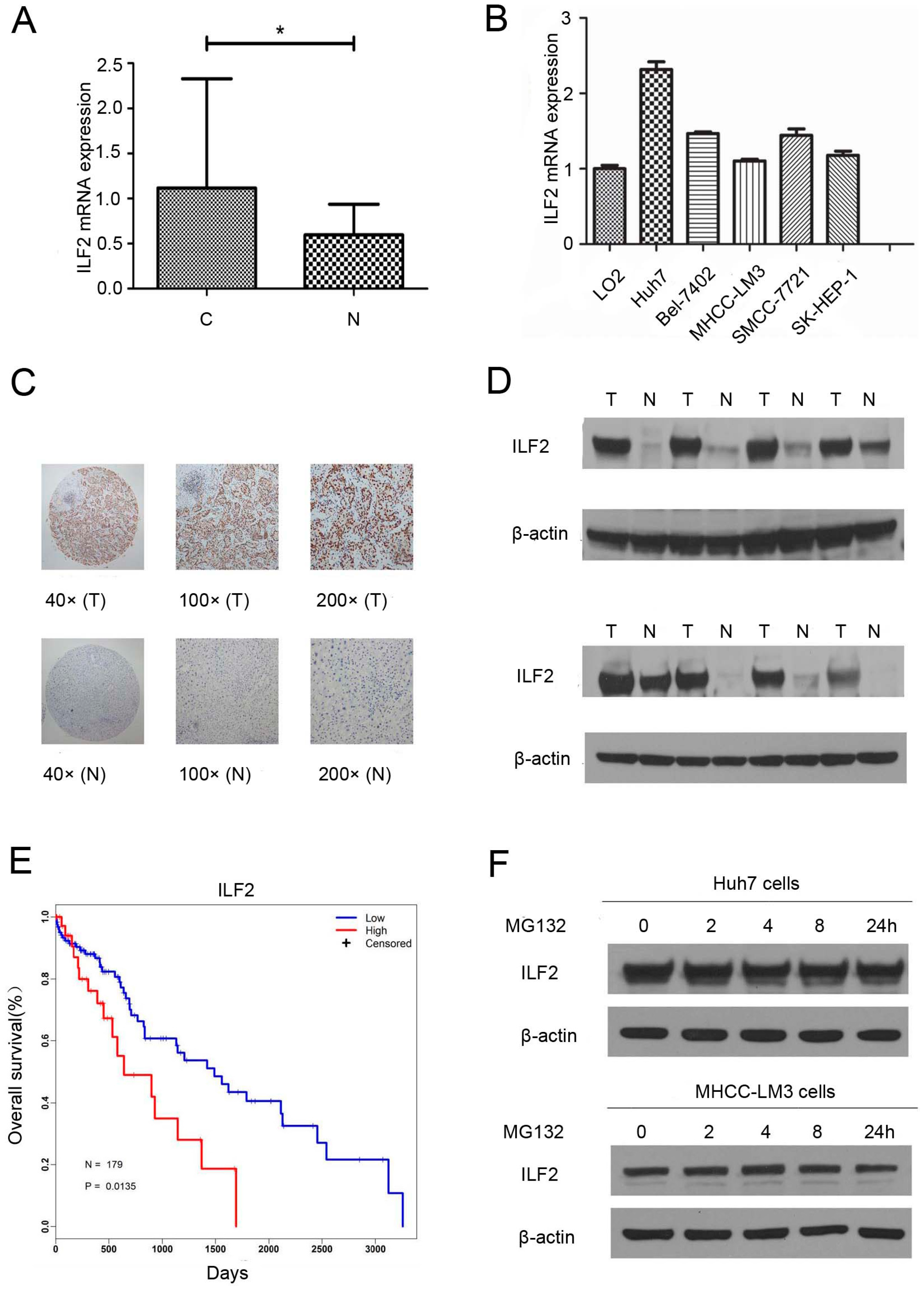
2.1. Interleukin Enhancer Binding Factor 2 Is Upregulated in Human Hepatocellular Carcinoma
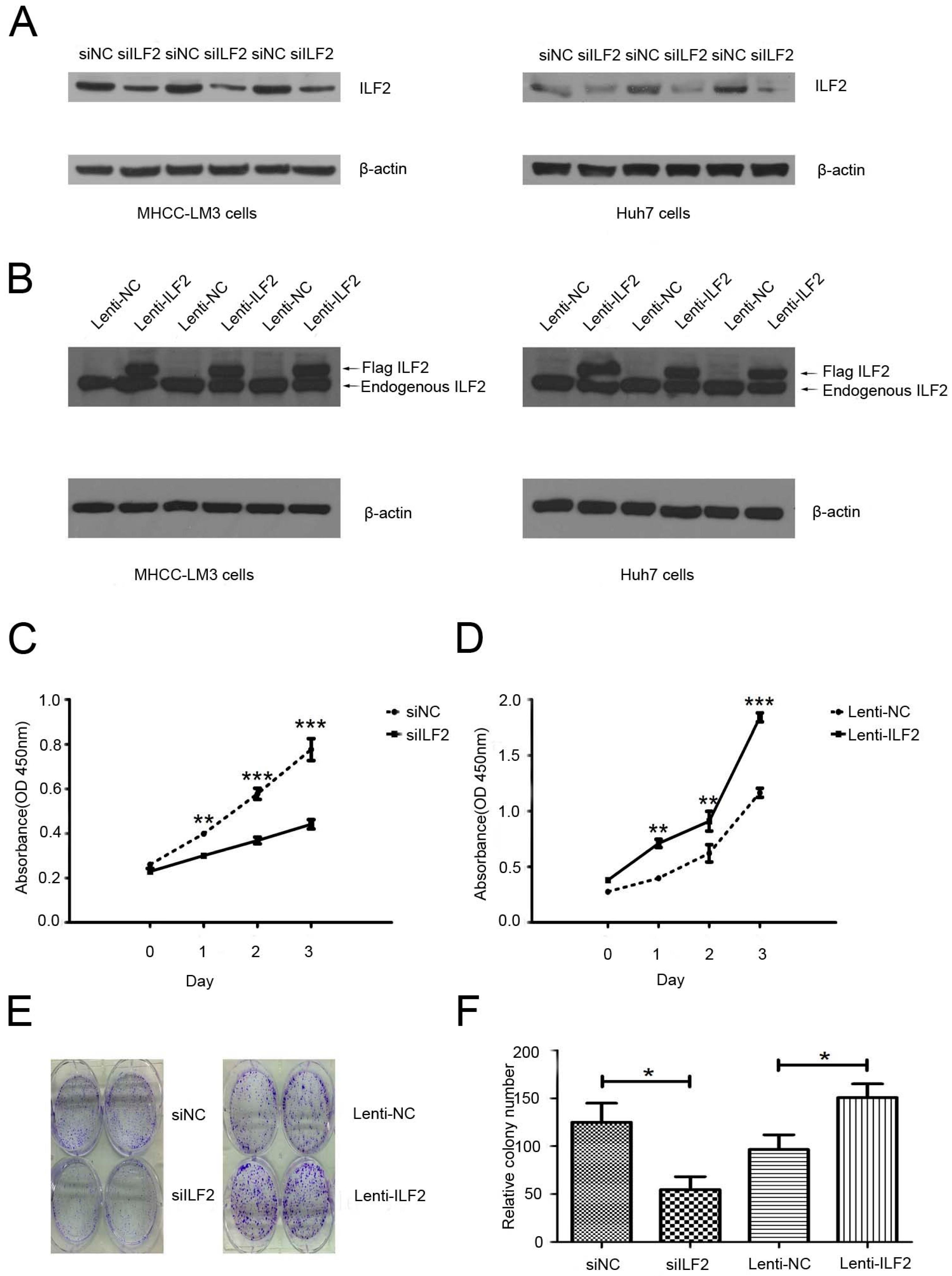
2.2. Effects of Interleukin Enhancer Binding Factor 2 on the Proliferation of Liver Cancer Cells
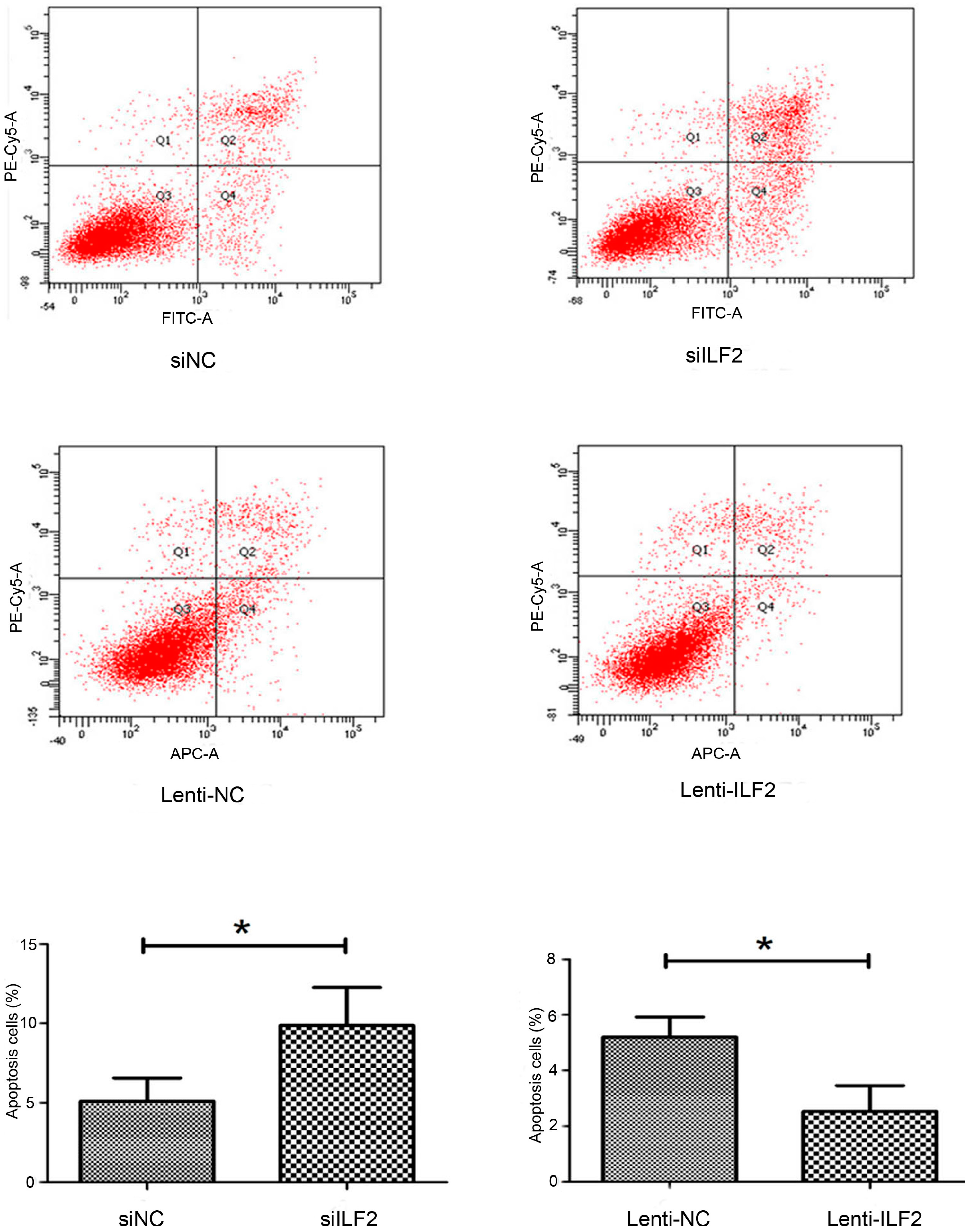
2.3. Effects of Interleukin Enhancer Binding Factor 2 on Apoptosis of Liver Cancer Cells
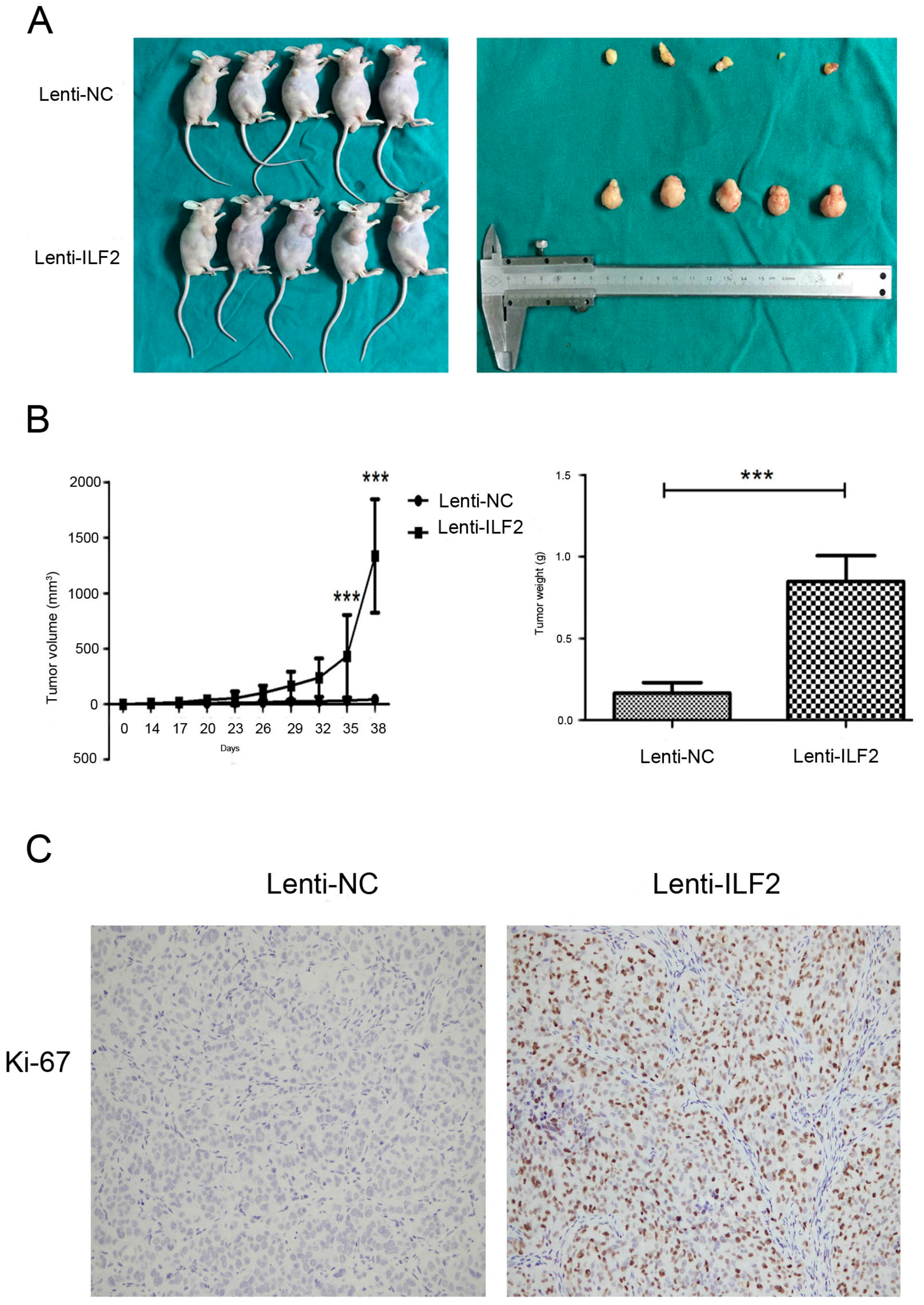
2.4. Overexpression of Interleukin Enhancer Binding Factor 2 Promotes Tumor Growth in a Xenograft Model
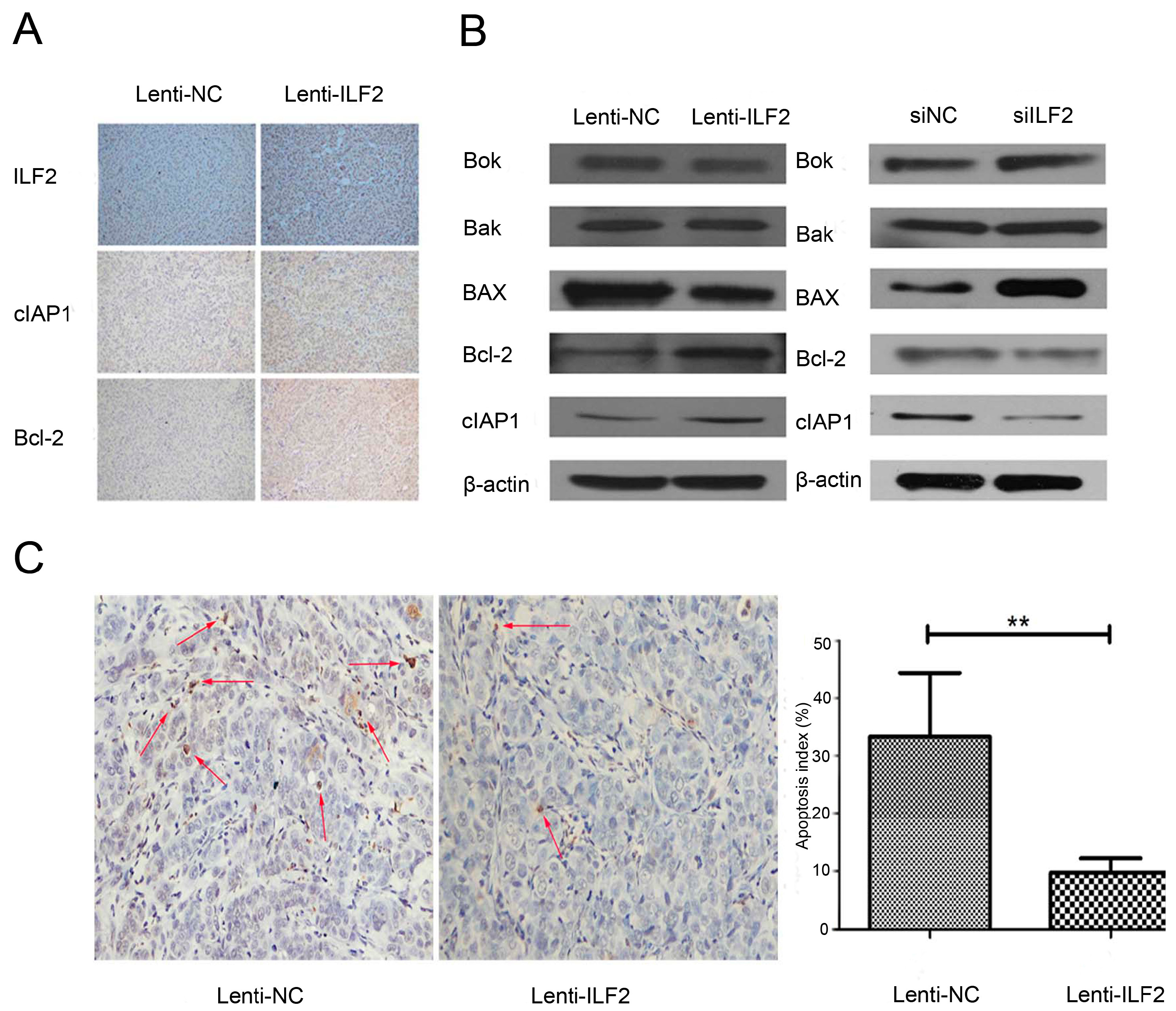
2.5. Interleukin Enhancer Binding Factor 2 May Suppress Apoptosis in Tumors via Regulation of Pro-Apoptotic Proteins and Anti-Apoptotic Proteins
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cultures
4.2. Lentiviral Transfection and siRNA Knockdown
4.3. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling Assay
4.4. Immunohistochemistry
4.5. Cell Viability Assay
4.6. Western Blotting
4.7. Colony Formation Assay
4.8. Tumor Xenografts
4.9. Apoptosis Assay
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Maluccio, M.; Covey, A. Recent progress in understanding, diagnosing, and treating hepatocellular carcinoma. CA Cancer J. Clin. 2012, 62, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Marcoulatos, P.; Koussidis, G.; Mamuris, Z.; Velissariou, V.; Vamvakopoulos, N.C. Mapping interleukin enhancer binding factor 2 gene (ILF2) to human chromosome 1 (1q11-qter and 1p11-p12) by polymerase chain reaction amplification of human-rodent somatic cell hybrid DNA templates. J. Interferon Cytokine Res. 1996, 16, 1035–1038. [Google Scholar] [PubMed]
- Zhao, G.; Shi, L.; Qiu, D.; Hu, H.; Kao, P.N. NF45/ILF2 tissue expression, promoter analysis, and interleukin-2 transactivating function. Exp. Cell Res. 2005, 305, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Kao, P.N.; Chen, L.; Brock, G.; Ng, J.; Kenny, J.; Smith, A.J.; Corthésy, B. Cloning and expression of cyclosporin A- and FK506-sensitive nuclear factor of activated T-cells: NF45 and NF90. J. Biol. Chem. 1994, 269, 20691–20699. [Google Scholar] [PubMed]
- Faye, M.D.; Graber, T.E.; Liu, P.; Thakor, N.; Baird, S.D.; Durie, D.; Holcik, M. Nucleotide composition of cellular internal ribosome entry sites defines dependence on NF45 and predicts a posttranscriptional mitotic regulon. Mol. Cell. Biol. 2013, 33, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Graber, T.E.; Baird, S.D.; Kao, P.N.; Mathews, M.B.; Holcik, M. NF45 functions as an IRES trans-acting factor that is required for translation of cIAP1 during the unfolded protein response. Cell Death Differ. 2010, 17, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Fernandez, L.A.; Parraga, M.; del Mazo, J. ILF2 is regulated during meiosis and associated to transcriptionally active chromatin. Mech. Dev. 2002, 111, 153–157. [Google Scholar] [CrossRef]
- Curtis, D.; Lehmann, R.; Zamore, P.D. Translational regulation in development. Cell 1995, 81, 171–178. [Google Scholar] [CrossRef]
- Meric, F.; Searfoss, A.M.; Wormington, M.; Wolffe, A.P. Masking and unmasking maternal mRNA. The role of polyadenylation, transcription, splicing, and nuclear history. J. Biol. Chem. 1996, 271, 30804–30810. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Aoki, K.; Higuchi, T.; Todaka, H.; Morisawa, K.; Tamaki, N.; Hatano, E.; Fukushima, A.; Taniguchi, T.; Agata, Y. The NF90–NF45 complex functions as a negative regulator in the microRNA processing pathway. Mol. Cell. Biol. 2009, 29, 3754–3769. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Licklider, L.J.; Gygi, S.P.; Reed, R. Comprehensive proteomic analysis of the human spliceosome. Nature 2002, 419, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Scherl, A.; Coute, Y.; Deon, C.; Calle, A.; Kindbeiter, K.; Sanchez, J.C.; Greco, A.; Hochstrasser, D.; Diaz, J.J. Functional proteomic analysis of human nucleolus. Mol. Biol. Cell 2002, 13, 4100–4109. [Google Scholar] [CrossRef] [PubMed]
- Langland, J.O.; Kao, P.N.; Jacobs, B.L. Nuclear factor-90 of activated T-cells: A double-stranded RNA-binding protein and substrate for the double-stranded RNA-dependent protein kinase, PKR. Biochemistry 1999, 38, 6361–6368. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.C.; Vestal, D.J.; Xu, Z.; Bandyopadhyay, S.; Guo, W.; Erme, S.M.; Williams, B.R.; Sen, G.C. DRBP76, a double-stranded RNA-binding nuclear protein, is phosphorylated by the interferon-induced protein kinase, PKR. J. Biol. Chem. 1999, 274, 20432–20437. [Google Scholar] [CrossRef] [PubMed]
- Saunders, L.R.; Perkins, D.J.; Balachandran, S.; Michaels, R.; Ford, R.; Mayeda, A.; Barber, G.N. Characterization of two evolutionarily conserved, alternatively spliced nuclear phosphoproteins, NFAR-1 and -2, that function in mRNA processing and interact with the double-stranded RNA-dependent protein kinase, PKR. J. Biol. Chem. 2001, 276, 32300–32312. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; He, X.; Qiu, X.; Liu, X.; Sun, G.; Guo, J.; Ding, Z.; Yang, L.; Ban, N.; Tao, T.; et al. Expression of NF45 correlates with malignant grade in gliomas and plays a pivotal role in tumor growth. Tumour Biol. 2014, 35, 10149–10157. [Google Scholar] [CrossRef] [PubMed]
- Ni, T.; Mao, G.; Xue, Q.; Liu, Y.; Chen, B.; Cui, X.; Lv, L.; Jia, L.; Wang, Y.; Ji, L. Upregulated expression of ILF2 in non-small cell lung cancer is associated with tumor cell proliferation and poor prognosis. J. Mol. Histol. 2015, 46, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Zhu, J.; Zhang, J.; Zhang, S.; Li, M.; Ni, R.; Liu, J.; Qiu, H.; Chen, W.; Wang, H.; et al. Expression and clinical role of NF45 as a novel cell cycle protein in esophageal squamous cell carcinoma (ESCC). Tumour Biol. 2015, 36, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Haskins, W.E.; Eedala, S.; Jadhav, Y.L.; Labhan, M.S.; Pericherla, V.C.; Perlman, E.J. Insights on neoplastic stem cells from gel-based proteomics of childhood germ cell tumors. Pediatr. Blood Cancer 2012, 58, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Shamanna, R.A.; Hoque, M.; Pe’Ery, T.; Mathews, M.B. Induction of p53, p21 and apoptosis by silencing the NF90/NF45 complex in human papilloma virus-transformed cervical carcinoma cells. Oncogene 2013, 32, 5176–5185. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; McKinney, K.Q.; Ghosh, S.; Iannitti, D.A.; Martinie, J.B.; Caballes, F.R.; Russo, M.W.; Ahrens, W.A.; Lundgren, D.H.; Han, D.K.; et al. Subcellular tissue proteomics of hepatocellular carcinoma for molecular signature discovery. J. Proteome Res. 2011, 10, 5070–5083. [Google Scholar] [CrossRef] [PubMed]
- Tomczak, K.; Czerwińska, P.; Wiznerowicz, M. The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge. Contemp. Oncol. (Pozn.) 2015, 19, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Girish, V.; Vijayalakshmi, A. Affordable image analysis using NIH Image/ImageJ. Indian J. Cancer 2004, 41, 47. [Google Scholar] [PubMed]
- Satoh, M.; Shaheen, V.M.; Kao, P.N.; Okano, T.; Shaw, M.; Yoshida, H.; Richards, H.B.; Reeves, W.H. Autoantibodies define a family of proteins with conserved double-stranded RNA-binding domains as well as DNA binding activity. J. Biol. Chem. 1999, 274, 34598–34604. [Google Scholar] [CrossRef] [PubMed]
- Ting, N.S.; Kao, P.N.; Chan, D.W.; Lintott, L.G.; Lees-Miller, S.P. DNA-dependent protein kinase interacts with antigen receptor response element binding proteins NF90 and NF45. J. Biol. Chem. 1998, 273, 2136–2145. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Gong, C.; Ji, L.; Liu, X.; Wang, Y.; Wang, L.; Shao, M.; Yang, L.; Fan, S.; Xiao, Y.; et al. NF45 overexpression is associated with poor prognosis and enhanced cell proliferation of pancreatic ductal adenocarcinoma. Mol. Cell. Biochem. 2015, 410, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Renault, T.T.; Manon, S. Bax: Addressed to kill. Biochimie 2011, 93, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, T.; Kitagawa, M.; Hasegawa, M.; Ikeda, S.; Akashi, T.; Takizawa, T.; Hirokawa, K.; Koike, M. Expression of IAP family proteins in esophageal cancer. Exp. Mol. Pathol. 2004, 76, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Abe, S.; Seidlar, H.B.; Nagaoka, S.; Takemura, T.; Utsuyama, M.; Kitagawa, M.; Hirokawa, K. Expression of IAP family proteins in colon cancers from patients with different age groups. Cancer Immunol. Immunother. 2004, 53, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, M.; Cantú, D.; Herrera, N.; Lopez, C.M.; de la Garza, J.G.; Maldonado, V.; Melendez-Zajgla, J. Inhibitors of apoptosis proteins in human cervical cancer. BMC Cancer 2006, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Berriguete, G.; Fraile, B.; de Bethencourt, F.R.; Prieto-Folgado, A.; Bartolome, N.; Nuñez, C.; Prati, B.; Martinez-Onsurbe, P.; Olmedilla, G.; Paniagua, R.; et al. Role of IAPs in prostate cancer progression: Immunohistochemical study in normal and pathological (benign hyperplastic, prostatic intraepithelial neoplasia and cancer) human prostate. BMC Cancer 2010, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, C.; Qian, L.; Zhang, C.; Wu, K.; Yang, C.; Yan, D.; Wu, X.; Shi, J. NF45 inhibits cardiomyocyte apoptosis following myocardial ischemia-reperfusion injury. Pathol. Res. Pract. 2015, 211, 955–962. [Google Scholar] [CrossRef] [PubMed]
- EI-Emshaty, H.M.; Saad, E.A.; Toson, E.A.; Abdel, M.C.; Gadelhak, N.A. Apoptosis and cell proliferation: Correlation with Bcl-2 and P53 oncoprotein expression in human hepatocellular carcinoma. Hepatogastroenterology 2014, 61, 1393–1401. [Google Scholar] [PubMed]
- Zender, L.; Spector, M.S.; Xue, W.; Flemming, P.; Cordon-Cardo, C.; Silke, J.; Fan, S.T.; Luk, J.M.; Wigler, M.; Hannon, G.J.; et al. Identification and validation of oncogenes in liver cancer using an integrative oncogenomic approach. Cell 2006, 125, 1253–1267. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Altan-Bonnet, N.; Parrott, A.M.; Arrigo, C.J.; Li, Q.; Khaleduzzaman, M.; Li, H.; Lee, C.G.; Pe’ery, T.; Mathews, M.B. Nuclear factor 45 (NF45) is a regulatory subunit of complexes with NF90/110 involved in mitotic control. Mol. Cell. Biol. 2008, 28, 4629–4641. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Huang, H.; Ding, L.; Zhu, P.; Saiyin, H.; Ji, G.; Zuo, J.; Han, D.; Pan, Y.; Ding, D.; et al. Regulation of cell cycle of hepatocellular carcinoma by NF90 through modulation of cyclin E1 mRNA stability. Oncogene 2015, 34, 4460–4470. [Google Scholar] [CrossRef] [PubMed]
- Barber, G.N. The NFAR’s (nuclear factors associated with dsRNA): Evolutionarily conserved members of the dsRNA binding protein family. RNA Biol. 2009, 6, 35–39. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Patients (Total = 72) | ILF2 | p-Value a | ||
|---|---|---|---|---|---|
| Negative | Positive | ||||
| Gender | Male | 61 | 24 | 37 | |
| Female | 11 | 6 | 5 | 0.347 | |
| Age (years) | ≥50 | 42 | 16 | 26 | |
| <50 | 30 | 14 | 16 | 0.467 | |
| Tumor size (cm) | ≥10 | 24 | 6 | 18 | |
| <10 | 48 | 24 | 24 | 0.043 * | |
| TNM stage | I–II | 41 | 15 | 26 | |
| III–IV | 31 | 15 | 16 | 0.315 | |
| Vascular invasion | Yes | 23 | 11 | 12 | |
| No | 49 | 19 | 30 | 0.468 | |
| Histopathologic grading | Good/moderate | 45 | 18 | 27 | |
| Poor | 27 | 12 | 15 | 0.711 | |
| Cirrhosis | Present | 27 | 11 | 16 | |
| Absent | 45 | 19 | 26 | 0.902 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, S.; Jiang, X.; Ding, C.; Du, C.; Owusu-Ansah, K.G.; Weng, X.; Hu, W.; Peng, C.; Lv, Z.; Tong, R.; et al. Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2016, 17, 1373. https://doi.org/10.3390/ijms17081373
Cheng S, Jiang X, Ding C, Du C, Owusu-Ansah KG, Weng X, Hu W, Peng C, Lv Z, Tong R, et al. Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2016; 17(8):1373. https://doi.org/10.3390/ijms17081373
Chicago/Turabian StyleCheng, Shaobing, Xu Jiang, Chaofeng Ding, Chengli Du, Kwabena Gyabaah Owusu-Ansah, Xiaoyu Weng, Wendi Hu, Chuanhui Peng, Zhen Lv, Rongliang Tong, and et al. 2016. "Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in Hepatocellular Carcinoma" International Journal of Molecular Sciences 17, no. 8: 1373. https://doi.org/10.3390/ijms17081373
APA StyleCheng, S., Jiang, X., Ding, C., Du, C., Owusu-Ansah, K. G., Weng, X., Hu, W., Peng, C., Lv, Z., Tong, R., Xiao, H., Xie, H., Zhou, L., Wu, J., & Zheng, S. (2016). Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in Hepatocellular Carcinoma. International Journal of Molecular Sciences, 17(8), 1373. https://doi.org/10.3390/ijms17081373