
New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State

 ,
, 

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of ERW
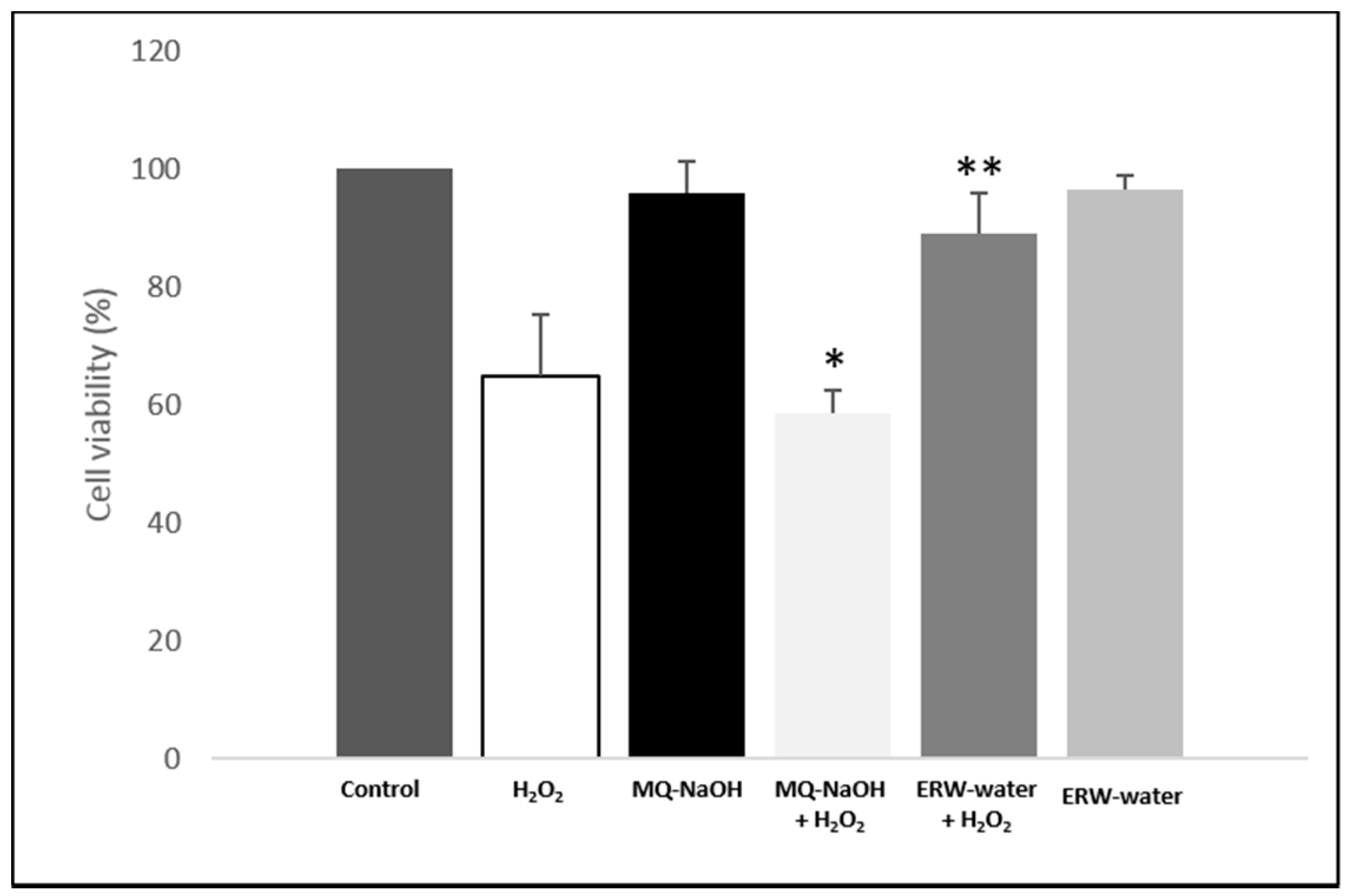
2.2. ERW Protects Against H2O2-Induced Toxicity
2.3. ERW Suppressed H2O2-Induced Oxidative Stress Production
2.4. Effect of ERW on Antioxidant Enzymes
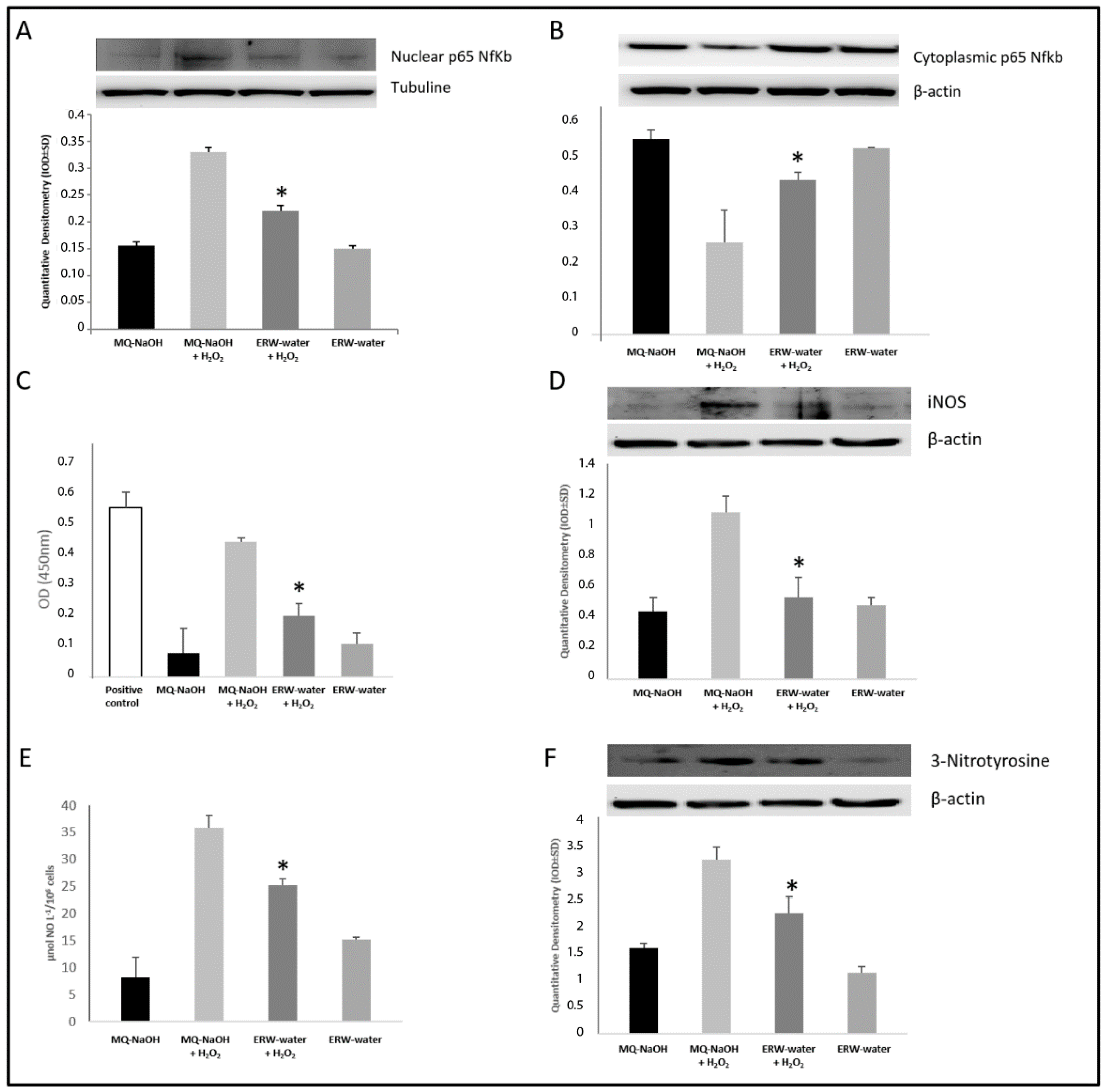
2.5. ERW Influence NF-κB/iNOS Pathway and 3-Nitrotyrosine Protein
3. Discussion
4. Materials and Methods
4.1. Apparatus Producing ERW and Procedure for the Measurement of ORP, pH, Eh and rH2
4.2. Preparation of Medium and Cell Culture
4.3. MTT Assay for Cell Viability and Cytotoxicity
4.4. NitroBlue-Tetrazolium (NBT) Assay
4.5. Western Blot Analysis
4.6. Cu, Zn-Superoxide Dismutase (SOD) and Catalase Activity
4.7. Glutathione Peroxidase (GPx) and Glutathione Reductase (GR) Activity
4.8. Measurement of GSH
4.9. Measurement of NO Release
4.10. Measurement of NF-κB p65 DNA Binding Activity
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Franceschelli, S.; Pesce, M.; Ferrone, A.; de Lutiis, M.A.; Patruno, A.; Grilli, A. Astaxanthin treatment confers protection against oxidative stress in U937 cells stimulated with lipopolysaccharide reducing O2− production. PLoS ONE 2014, 9, e88359. [Google Scholar] [CrossRef] [PubMed]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Vinciguerra, I.; de Lutiis, M.A.; Grilla, A.; Felaco, M.; Patruno, A. Phosphodiesterase type-5 inhibitor and oxidative stress. Int. J. Immunopathol. Pharmacol. 2008, 21, 879–889. [Google Scholar] [PubMed]
- Stohs, S. The role of free radicals in toxicity and disease. J. Basic Clin. Physiol. Pharmacol. 1995, 6, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Franceschelli, S.; Ferrone, A.; Pesce, M.; Riccioni, G.; Speranza, L. Biological functional relevance of asymmetric dimethylarginine (ADMA) in cardiovascular disease. Int. J. Mol. Sci. 2013, 14, 24412–24421. [Google Scholar] [CrossRef] [PubMed]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Menghini, L.; Patruno, A.; Vinciguerra, I.; de Lutiis, M.A.; Felaco, M.; Felaco, P.; Grilli, A. Anti-inflammatory properties of the plant Verbascum mallophorum. J. Biol. Regul. Homeost. Agents 2009, 23, 189–195. [Google Scholar] [PubMed]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Reale, M.; Menghini, L.; Vinciguerra, I.; de Lutiis, M.A.; Felaco, M.; Grilli, A. Antiinflammatory effects in THP-1 cells treated with verbascoside. Phytother. Res. 2010, 24, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Rizzuto, A.; La Fratta, I.; Tatangelo, R.; Campagna, G.; Iannasso, M.; Ferrone, A.; Franceschelli, S.; Speranza, L.; Patruno, A.; et al. Super-oxide anion production and antioxidant enzymatic activities associated with the executive functions in peripheral blood mononuclear cells of healthy adult samples. Neurosci. Res. 2016, 106, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.M.; Shafaie, Y. Dietary lutein modulates inducible nitric oxide synthase (iNOS) gene and protein expression in mouse macrophage cells (RAW 264.7). Mol. Nutr. Food Res. 2007, 51, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Speranza, L.; Pesce, M.; Franceschelli, S.; Mastrangelo, F.; Patruno, A.; de Lutiis, M.A.; Tete, S.; Felaco, M.; Grilli, A. The role of inducible nitric oxide synthase and haem oxygenase 1 in growth and development of dental tissue. Cell Biochem. Funct. 2012, 30, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo di Maio, F.; Lohinai, Z.; D’Arcangelo, C.; de Fazio, P.E.; Speranza, L.; de Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M. Nitric oxide synthase in healthy and inflamed human dental pulp. J. Dent. Res. 2004, 83, 312–336. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, X.; Chen, J.; Jiao, R.; Wang, L.; Li, Y.M.; Zuo, Y.; Liu, Y.; Lei, L.; Ma, K.Y.; et al. Biology of ageing and role of dietary antioxidants. BioMed Res. Int. 2014, 2014, 831841. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; D’Orazio, N.; Salvatore, C.; Franceschelli, S.; Pesce, M.; Speranza, L. Carotenoids and vitamins C and E in the prevention of cardiovascular disease. Int. J. Vitam. Nutr. Res. 2012, 82, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Yan, H.; Hamasaki, T.; Kinjo, T.; Nakamichi, N.; Teruya, K.; Kabaiama, S.; Shirahata, S. Electrochemically reduced water protects neural cells from oxidative damage. Oxid. Med. Cell. Longev. 2014, 2014, 869121. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Hsu, Y.W.; Chen, W.K.; Ho, Y.C.; Lu, F.J. Enhanced induction of mitochondrial damage and apoptosis in human leukemia HL-60 cells due to electrolyzed-reduced water and glutathione. Biosci. Biotechnol. Biochem. 2009, 73, 280–287. [Google Scholar] [CrossRef]
- Shirahata, S.; Kabayama, S.; Nakano, M.; Miura, T.; Kusumoto, K.; Gotoh, M.; Hayashi, H.; Otsubo, K.; Morisawa, S.; Katakura, Y. Electrolyzed-reduced water scavenges active oxygen species and protects DNA from oxidative damage. Biochem. Biophys. Res. Commun. 1997, 8, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hamasaki, T.; Nakamichi, N.; Kashiwagi, T.; Ye, J.; Teruya, K.; Abe, M.; Yan, H.; Kinjo, T.; Kabayama, S.; et al. Suppressive effects of electrolyzed reduced water on alloxan-induced apoptosis and type 1 diabetes mellitus. Cytotechnology 2011, 63, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Nishimura, T.; Teruya, K.; Maki, T.; Komatsu, T.; Hamasaki, T.; Kashiwagi, T.; Kabayama, S.; Shim, S.Y.; Katakura, Y.; et al. Protective mechanism of reduced water against alloxan-induced pancreatic beta-cell damage: Scavenging effect against reactive oxygen species. Cytotechnology 2002, 40, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Spulber, S.; Edoff, K.; Hong, L.; Morisawa, S.; Shirahta, S.; Ceccatelli, S. Molecular hydrogen reduces LPS-induced neuroinflammation and promotes recovery from sickness behaviour in mice. PLoS ONE 2011, 7, e42078. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Kashiwaki, T.; Hamasaki, T.; Kinjo, T.; Teruya, K.; Kabayama, S.; Shirahata, S. The neuroprotective effects of electrolyzed reduced water and its model water containing molecular hydrogen and Pt nanoparticles. BMC Proc. 2011, 5 (Suppl. 8), 69. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Li, Y.; Hamasaki, T.; Komatsu, T.; Komatsu, T.; Kashiwagi, T.; Teruya, K.; Nishikawa, R.; Kawahara, T.; Osada, K.; et al. Inhibitory effect of electrolyzed reduced water on tumor angiogenesis. Biol. Pharm. Bull. 2008, 31, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Chambron, J. Biological and Therapeutic Characteristics of Electrolyzed Reduced Alkaline Water (ERAW). Water 2013, 5, 2094–2115. [Google Scholar] [CrossRef]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; Patruno, A.; Livia, P.; Carlucci, G.; Ferrone, V.; Carlucci, M.; de Lutiis, M.A.; Grilli, A.; et al. A novel biological role of α-mangostin in modulating inflammatory response through the activation of SIRT-1 signaling pathway. J. Cell. Physiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Suliman, H.B.; Piantadosi, C.A. Mitochondrial biogenesis: Regulation by endogenous gases during inflammation and organ stress. Curr. Pharm. Des. 2014, 20, 5653–5662. [Google Scholar] [CrossRef] [PubMed]
- Shirahata, S.; Hamasaki, T.; Teruya, K. Advanced research on the health benefit of reduced water. Trends Food Sci. Technol. 2012, 23, 124–131. [Google Scholar] [CrossRef]
- Huang, K.C.; Yang, C.C.; Lee, K.T.; Chien, C.T. Reduced hemodialysis-induced oxidative stress in end-stage renal disease patients by electrolyzed reduced water. Kidney Int. 2003, 64, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Hsu, S.P.; Yang, C.C.; Morisawa, S.; Otsubo, K.; Chien, C.T. Electrolysed-reduced water dialysate improves T-cell damage in end-stage renal disease patients with chronic haemodialysis. Nephrol. Dial. Transplant. 2010, 25, 2730–2737. [Google Scholar] [CrossRef]
- Kinjo, T.; Ye, J.; Yan, H.; Nakanishi, H.; Toh, K.; Nakamichi, N.; Kabayama, S.; Teruya, K.; Shirahata, S. Suppressive effects of electrochemically reduced water on matrix metalloproteinase-2 activities and in vitro invasion of human fibrosarcoma HT1080 cells. Cytotechnology 2012, 64, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-C.; Yang, C.C.; Hsu, S.P.; Lee, K.T.; Liu, H.W.; Morisawa, S.; Otsubo, K.; Chien, C.T. Electrolyzed-reduced water reduced hemodialysis-induced erythrocyte impairment in end-stage renal disease patients. Kidney Int. 2006, 70, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Schmatz, R.; Perreira, L.B.; Stefanello, N.; Mazzanti, C.; Spanevello, R.; Gutierres, J.; Bagatini, M.; Martins, C.C.; Abdalla, F.H.; Daci da Silva Serres, J.; et al. Effects of resveratrol on biomarkers of oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver and kidney of streptozotocin-induced diabetic rats. Biochimie 2012, 94, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Franceschelli, S.; Pesce, M.; Vinciguerra, I.; Ferrone, A.; Riccioni, G.; Patruno, A.; Grilli, A.; Felaco, M.; Speranza, L. Licocalchone-C extracted from Glycyrrhiza glabra inhibits lipopolysaccharide-interferon-γ inflammation by improving antioxidant conditions and regulating inducible nitric oxide synthase expression. Molecules 2011, 16, 5720–5734. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Ischiropoulos, H.; Cuzzocrea, S. Roles of nitric oxide and superoxide in inflammation. Methods Mol. Biol. 2003, 225, 291–303. [Google Scholar] [PubMed]
- Varga, Z.V.; Giricz, Z.; Liaudet, L.; Haskó, G.; Ferdinandy, P.; Pacher, P. Interplay of oxidative, nitrosative/nitrative stress, inflammation, cell death and autophagy in diabetic cardiomyopathy. Biochim. Biophys. Acta 2015, 1852, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, I.; Madar, Z.; Leikin-Frenkel, A.; Tamir, S. Glabridin, an isoflavan from licorice root, downregulates iNOS expression and activity under high-glucose stress and inflammation. Mol. Nutr. Food Res. 2015, 59, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Quijano, C.; Hernandez-Saavedra, D.; Castro, L.; McCord, J.M.; Freeman, B.A.; Radi, R. Reaction of peroxynitrite with Mn-superoxide dismutase. Role of the metal center in decomposition kinetics and nitration. J. Biol. Chem. 2001, 276, 11631–11638. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Nakano, H.; Hamada, H.I.; Ilami, N.; Nakazawa, R.; Ito, S. A novel bioactive haemodialysis System using dissolved dihydrogen (H2) produced by water electrolysis: A clinical trial. Nephrol. Dial. Transplant. 2010, 25, 3026–3033. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Toyoda, Y.; Sharma, P.; Evans, M.; Guthrie, N. Effectiveness of hydrogen rich water on antioxidant status of subjects with potential metabolic syndrome—An open label pilot study. J. Clin. Biochem. Nutr. 2010, 46, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Patruno, A.; Fornasari, E.; Di Stefano, A.; Cerasa, L.S.; Marinelli, L.; Baldassarre, L.; Sozio, P.; Turkez, H.; Franceschelli, S.; Ferrone, A.; et al. Synthesis of a novel cyclic prodrug of S-allyl-glutathione able to attenuate LPS-induced ROS production through the inhibition of MAPK pathways in U937 cells. Mol. Pharm. 2015, 12, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Reale, M.; de Lutiis, M.A.; Patruno, A.; Speranza, L. Modulation of MCP-1 and iNOS by 50-Hz sinusoidal electromagnetic field. Nitric Oxide 2006, 15, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Jollow, D.J.; Michell, J.R.; Zampaglionic Gillete, J.R. Bromoibenzene-induced Liver necrosis: Protective role of glutathione and evidence for 3,4-Bromobenzene oxide as hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Patruno, A.; Franceschelli, S.; Pesce, M.; Maccallini, C.; Fantacuzzi, M.; Speranza, L.; Ferrone, A.; de Lutiis, M.A.; Ricciotti, E.; Amoroso, R.; et al. Novel aminobenzyl-acetamidine derivative modulate the differential regulation of NOSs in LPS induced inflammatory response: Role of PI3K/Akt pathway. Biochim. Biophys. Acta 2012, 1820, 2095–2104. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eh (mV) | rH2 | pH | T (°C) | |
|---|---|---|---|---|
| Tap Water | 531.1 ± 17.67 | 33.34 ± 0.43 | 7.54 | 20 |
| MQ-water | 511.1 ± 34.64 | 30.18 ± 0.84 | 6.3 | 20 |
| MQ-NaOH | 477.6 ± 2.82 | 35.14 ± 0.06 | 9.5 | 20 |
| ERW | −210.73 ± 24.51 | 11.81 ± 0.58 | 9.47 | 20 |
| ERW-medium | 240.1 ± 4.94 | 22.91 ± 0.11 | 7.4 | 20 |
| MQ-NaOH medium | 450.1 ± 7.77 | 30.01 ± 0.18 | 7.4 | 20 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franceschelli, S.; Gatta, D.M.P.; Pesce, M.; Ferrone, A.; Patruno, A.; De Lutiis, M.A.; Grilli, A.; Felaco, M.; Croce, F.; Speranza, L. New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State. Int. J. Mol. Sci. 2016, 17, 1461. https://doi.org/10.3390/ijms17091461
Franceschelli S, Gatta DMP, Pesce M, Ferrone A, Patruno A, De Lutiis MA, Grilli A, Felaco M, Croce F, Speranza L. New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State. International Journal of Molecular Sciences. 2016; 17(9):1461. https://doi.org/10.3390/ijms17091461
Chicago/Turabian StyleFranceschelli, Sara, Daniela Maria Pia Gatta, Mirko Pesce, Alessio Ferrone, Antonia Patruno, Maria Anna De Lutiis, Alfredo Grilli, Mario Felaco, Fausto Croce, and Lorenza Speranza. 2016. "New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State" International Journal of Molecular Sciences 17, no. 9: 1461. https://doi.org/10.3390/ijms17091461
APA StyleFranceschelli, S., Gatta, D. M. P., Pesce, M., Ferrone, A., Patruno, A., De Lutiis, M. A., Grilli, A., Felaco, M., Croce, F., & Speranza, L. (2016). New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State. International Journal of Molecular Sciences, 17(9), 1461. https://doi.org/10.3390/ijms17091461