
Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses

Abstract
:1. Introduction
2. Results
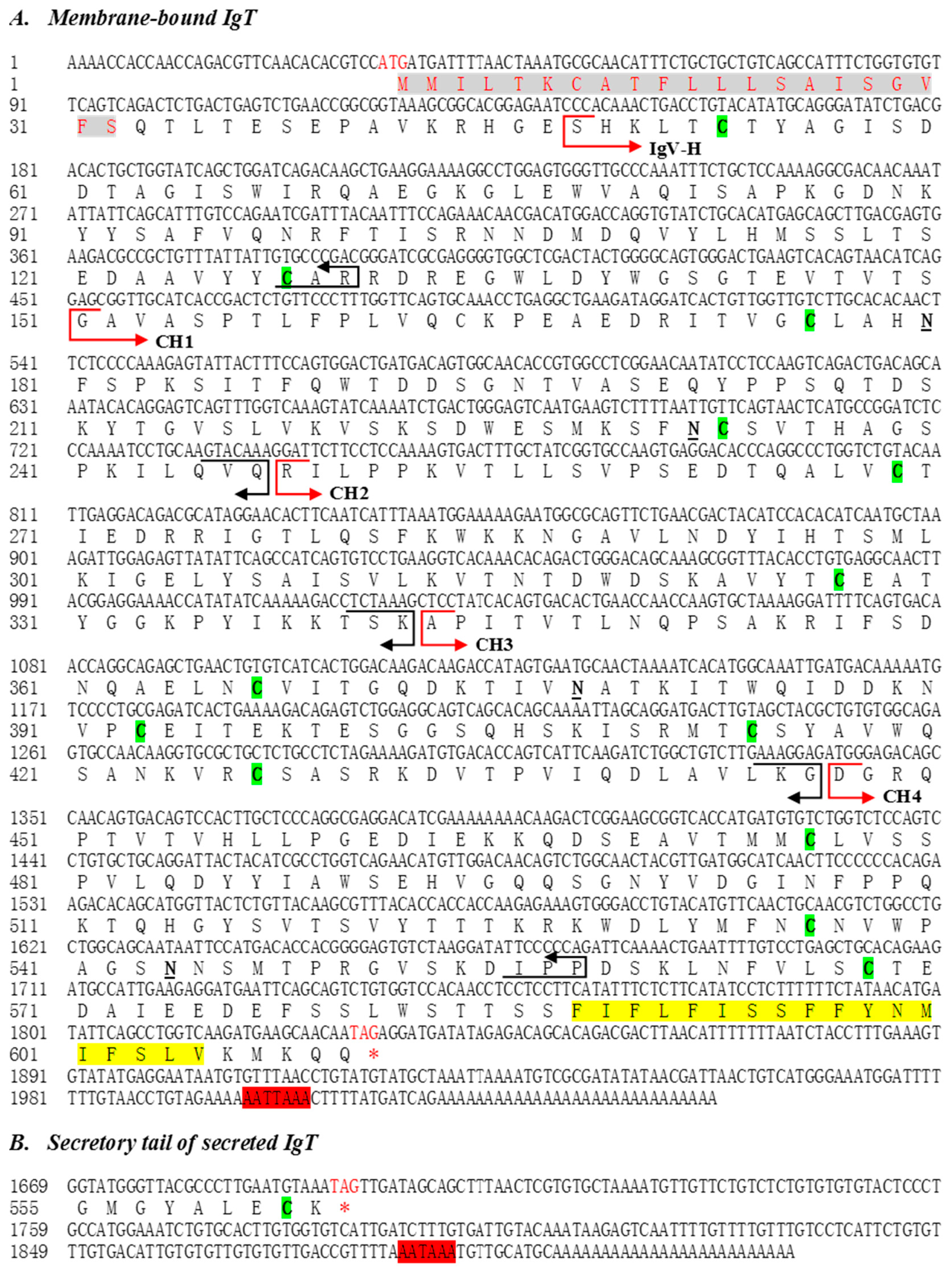
2.1. Molecular Characterization of Flounder IgT
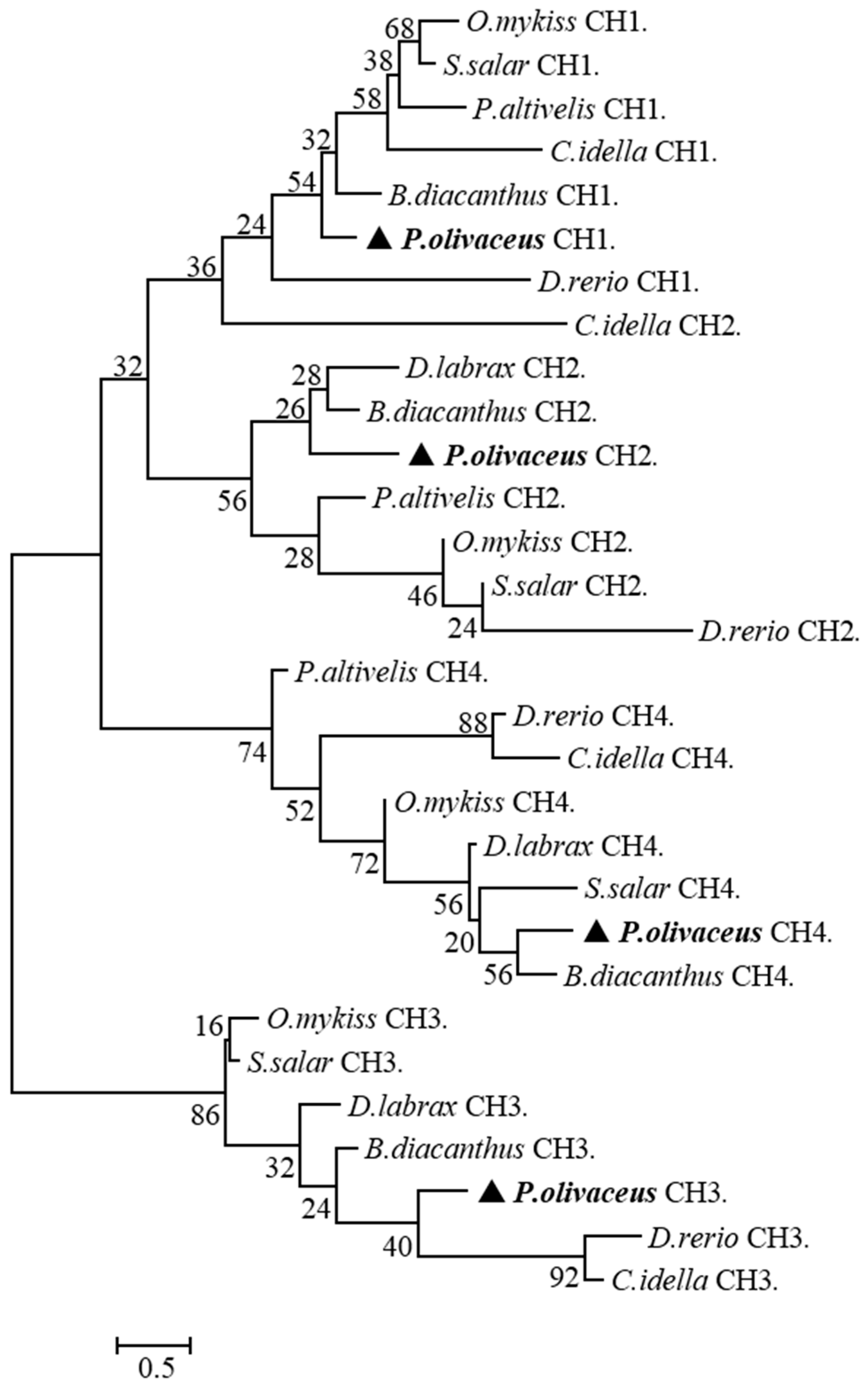
2.2. Multiple Sequence Alignment and Phylogenetic Analysis
2.3. Tissue Distribution of IgT Transcript in Healthy Flounder
2.4. Tissue Expression of IgT Post Bacterial Challenge
2.5. Expression of IgT after Inactivated E. tarda Immunization
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fish
4.3. Cloning of the Full-Length IgT
4.4. Domain Organization and Sequence Analyses
4.5. Quantitative Analysis of IgT Expression at the mRNA Level in Healthy Flounder
4.6. qRT-PCR Analysis of IgT Expression in Response to Live and Inactivated E. tarda
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Porter, R.R. The structure of the heavy chain of immunoglobulin and its relevance to the nature of the antibody-combining site. The second CIBA medal lecture. Biochem. J. 1967, 105, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Kabat, E.A.; Wu, T.T. Construction of a three-dimensional model of the polypeptide backbone of the variable region of kappa immunoglobulin light chains. Proc. Natl Acad. Sci. USA 1972, 69, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Kabat, E.A.; Wu, T.T.; Bilofsky, H. Variable region genes for the immunoglobulin framework are assembled from small segments of DNA-a hypothesis. Proc. Natl. Acad. Sci. USA 1978, 75, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.Z.; Xu, G.J.; Tang, X.Q.; Zhan, W.B. Monoclonal antibodies recognizing mucus immunoglobulin and surface immunoglobulin-positive cells of flounder (Paralichthys olivaceus). Vet. Immunol. Immunopathol. 2012, 145, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.J.; Zhan, W.B.; Ding, B.J.; Sheng, X.Z. Molecular cloning and expression analysis of polymeric immunoglobulin receptor in flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2013, 35, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Edholm, E.S.; Bengten, E.; Wilson, M. Insights into the function of IgD. Dev. Comp. Immunol. 2011, 35, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Wei, Z.G.; Hammarstrom, L.; Zhao, Y.F. The immunoglobulin δ gene in jawed vertebrates: A comparative overview. Dev. Comp. Immunol. 2011, 35, 975–981. [Google Scholar]
- Zhang, Y.A.; Salinas, I.; Sunyer, J.O. Recent findings on the structure and function of teleost IgT. Fish Shellfish Immunol. 2011, 31, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gomez, D.; Parra, D.; Takizawa, F.; Sunyer, J.O. IgT plays a prominent role in gill immune response of rainbow trout. Fish Shellfish Immunol. 2013, 34, 1686. [Google Scholar] [CrossRef]
- Rouaud, P.; Saintamand, A.; Saad, F.; Carrion, C.; Lecardeur, S.; Cogné, M.; Denizot, Y. Elucidation of the enigmatic IgD class-switch recombination via germline deletion of the IgH 3′ regulatory region. J. Exp. Med. 2014, 211, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Danilova, N.; Bussmann, J.; Jekosch, K.; Steiner, L.A. The immunoglobulin heavy-chain locus in zebrafish: Identification and expression of a previously unknown isotype, immunoglobulin Z. Nat. Immunol. 2005, 6, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Gambón-Deza, F.; Sánchez-Espinel, C. Presence of an unique IgT on the IgH locus in three-spined stickleback fish (Gasterosteus aculeatus) and the very recent generation of a repertoire of VH genes. Dev. Comp. Immunol. 2010, 34, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Aman, A.; Nakao, M.; Watanuki, H.; Sakai, M. Discovery of a novel immunoglobulin heavy chain gene chimera from common carp (Cyprinus carpio L.). Immunogenetics 2005, 57, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.D.; Cooper, M.D. Discovery of a unique Ig heavy-chain isotype (IgT) in rainbow trout: Implications for a distinctive B cell developmental pathway in teleost fish. Proc. Natl. Acad. Sci. USA 2005, 102, 6919–6924. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.H.; Wang, T.; Guo, Y.C.; Zhao, Z.H.; Li, N.; Zhao, Y.F. The immunoglobulin gene loci in the teleost Gasterosteus aculeatus. Fish Shellfish Immunol. 2010, 28, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Aman, A.; Sato, K.; Yamaguchi, R.; Sakai, M. Discovery of a new class of immunoglobulin heavy chain from fugu. Eur. J. Immunol. 2005, 35, 3320–3331. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Xiang, L.X.; Shao, J.Z. Identification and characterization of a novel immunoglobulin Z isotype in zebrafish: Implications for a distinct B cell receptor in lower vertebrates. Mol. Immunol. 2010, 47, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Ryo, S.; Wijdeven, R.H.M.; Tyagi, A.; Hermsen, T.; Kono, T.; Karunasagar, I.; Rombout, J.H.; Sakai, M.; Kemenade, B.M.L.V.; Savan, R. Common carp have two subclasses of bony fish specific antibody IgZ showing differential expression in response to infection. Dev. Comp. Immunol. 2010, 34, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Young, M.; Li, E.S.; Branham, E.; Browne, Z.; Koufteros, C.A.; Pham, S.E.; Kim, J.C.; Stebbins, E.A.; Wenck, F.E.; Zhang, A.L.; et al. Investigating distinct immunization routes in zebrafish on mucosal immunity. J. Immunol. 2016, 196, 137–145. [Google Scholar]
- Fillatreau, S.; Six, A.; Magadan, S.; Castro, R.; Sunyer, J.O.; Boudinot, P. The astonishing diversity of Ig classes and B cell repertoires in teleost fish. Front. Immunol. 2013, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Yasuike, M.; de Boer, J.; von Schalburg, K.R.; Cooper, G.A.; McKinnel, L.; Messmer, A.; So, S.; Davidson, W.S.; Koop, B.F. Evolution of duplicated IgH loci in Atlantic salmon, Salmo salar. BMC Genom. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Henkel, C.V.; Dirks, R.P.; Jansen, H.J.; Forlenza, M.; Wiegertjes, G.F.; Howe, K.; van den Thillart, G.E.E.J.M.; Spaink, H.P. Comparison of the exomes of common carp (Cyprinus carpio) and zebrafish (Danio rerio). Zebrafish 2012, 9, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Mashoof, S.; Pohlenz, C.; Chen, P.L.; Deiss, T.C.; Iii, D.G.; Buentello, A.; Criscitiello, M.F. Expressed IgH μ and τ transcripts share diversity segment in ranched Thunnus orientalis. Dev. Comp. Immunol. 2014, 43, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Rombout, J.H.W.M.; Yang, G.; Kiron, V. Adaptive immune responses at mucosal surfaces of teleost fish. Fish Shellfish Immunol. 2014, 40, 634–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Parra, D.; Gómez, D.; Salinas, I.; Zhang, Y.A.; Von, G.J.L.; Heinecke, R.D.; Buchmann, K.; LaPatra, S.; Sunyer, J.O. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Liu, W.J.; Wu, K.; Wang, W.M.; Zhang, X.Z. sIgZ exhibited maternal transmission in embryonic development and played a prominent role in mucosal immune response of Megalabrama amblycephala. Fish Shellfish Immunol. 2016, 54, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, S.; Buonocore, F.; Albanese, F.; Scapigliati, G.; Gerdol, M.; Oreste, U.; Coscia, M.R. New insights into evolution of IgT genes coming from Antarctic teleosts. Mar. Genom. 2015, 24, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, X.H. Edwardsiella tarda: An intriguing problem in aquaculture. Aquaculture 2014, 431, 129–135. [Google Scholar] [CrossRef]
- Aoki, T.; Hirono, I. Immune relevant genes of Japanese flounder, Paralichthys olivaceus. Comp. Biochem. Physiol. D 2006, 1, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.H.; Pack, H.J.; Do, J.W.; Bang, J.D.; Cho, H.R.; Ko, B.K. Flounder (Paralichthys olivaceus) cDNA encoding a secreted immunoglobulin M heavy chain. Fish Shellfish Immunol. 2001, 11, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Bengtén, E.; Quiniou, S.M.; Stuge, T.B.; Katagiri, T.; Miller, N.W.; Clem, L.W.; Warr, G.W.; Wilson, M. The IgH locus of the channel catfish, Ictalurus punctatus, contains multiple constant region gene sequences: Different genes encode heavy chains of membrane and secreted IgD. J. Immunol. 2002, 169, 2488–2497. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.Y.; Sun, B.J.; Luo, Y.P.; Zhang, Y.A.; Nie, P. Distribution of IgM, IgD and IgZ in mandarin fish, Siniperca chuatsi lymphoid tissues and their transcriptional changes after Flavobacterium columnare stimulation. Aquaculture 2009, 288, 14–21. [Google Scholar] [CrossRef]
- Woof, J.M.; Mestecky, J. Mucosal immunoglobulins. Immunol. Rev. 2005, 206, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Salinas, I.; Zhang, Y.A.; Sunyer, J.O. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef] [PubMed]
- Hirono, I.; Nam, B.H.; Enomoto, J.; Uchino, K.; Aoki, T. Cloning and characterisation of a cDNA encoding Japanese flounder Paralichthys olivaceus IgD. Fish Shellfish Immunol. 2003, 15, 63–70. [Google Scholar] [CrossRef]
- Srisapoome, P.; Ohira, T.; Hirono, I.; Aoki, T. Genes of the constant regions of functional immunoglobulin heavy chain of Japanese flounder, Paralichthys olivaceus. Immunogenetics 2004, 56, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.; Feinstein, A. Structure and function of the constant regions of immunoglobulins. Q. Rev. Biophys. 1976, 9, 135–180. [Google Scholar] [CrossRef] [PubMed]
- Elfakess, R.; Dikstein, R. A translation initiation element specific to mRNAs with very short 5′-UTR that also regulates transcription. PLoS ONE 2008, 3, e3094. [Google Scholar] [CrossRef] [PubMed]
- Moor, C.H.; Meijer, H.; Lissenden, S. Mechanisms of translational control by the 3′-UTR in development and differentiation. Semin. Cell Dev. Boil. 2005, 16, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, G.S.; Dickson, K.S.; Gray, N.K. Regulation of mRNA translation by 5′-and 3′-UTR-binding factors. Trends Biochem. Sci. 2003, 28, 182–188. [Google Scholar] [CrossRef]
- Mazumder, B.; Seshadri, V.; Fox, P.L. Translational control by the 3′-UTR: The ends specify the means. Trends Biochem. Sci. 2003, 28, 91–98. [Google Scholar] [CrossRef]
- Kato, G.; Takano, T.; Sakai, T.; Matsuyama, T.; Sano, N.; Nakayasu, C. Cloning and expression analyses of a unique IgT in ayu Plecoglossus altivelis. Fish. Sci. 2014, 81, 29–36. [Google Scholar] [CrossRef]
- Tariku Markos, T.; Kristoffer, L.K.; Ivar, H. Molecular cloning of IgT from Atlantic salmon, and analysis of the relative expression of τ, μ, and δ in different tissues. Vet. Immunol. Immunopathol. 2011, 139, 17–26. [Google Scholar]
- Liu, Y.; Zhang, S.C.; Jiang, G.L.; Yang, D.; Lian, J.H.; Yang, Y.W. The development of the lymphoid organs of flounder, Paralichthys olivaceus, from hatching to 13 months. Fish Shellfish Immunol. 2004, 16, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Press, C.M.; Evensen, Ø. The morphology of the immune system in teleost fishes. Fish Shellfish Immunol. 1999, 9, 309–318. [Google Scholar] [CrossRef]
- Ototake, M.; Iwama, G.K.; Nakanishi, T. The uptake of bovine serum albumin by the skin of bath-immunised rainbow trout Oncorhynchus mykiss. Fish Shellfish Immunol. 1996, 6, 321–333. [Google Scholar] [CrossRef]
- Kato, G.; Takano, T.; Sakai, T.; Matsuyama, T.; Nakayasu, C. Vibrio anguillarum bacterin uptake via the gills of Japanese flounder and subsequent immune responses. Fish Shellfish Immunol. 2013, 35, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.; Wrathmell, A.B.; Harris, J.E.; Grayson, T.H. Gut immunology in fish: A review. Dev. Comp. Immunol. 1988, 12, 453–480. [Google Scholar] [CrossRef]
- Castro, R.; Jouneau, L.; Pham, H.P.; Bouchez, O.; Giudicelli, V.; Lefranc, M.P.; Fillatreau, S. Teleost fish mount complex clonal IgM and IgT responses in spleen upon systemic viral infection. PLoS Pathog. 2013, 9, e1003098. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhan, W.B.; Xing, J.; Sheng, X.Z. Production, characterization and applicability of monoclonal antibodies to immunoglobulin of Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2007, 23, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.J.; Sheng, X.Z.; Xing, J.; Zhan, W.B. Effect of temperature on immune response of Japanese flounder (Paralichthys olivaceus) to inactivated lymphocystis disease virus (LCDV). Fish Shellfish Immunol. 2011, 30, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Tang, X.Q.; Sheng, X.Z.; Xing, J.; Zhan, W.B. Immune responses of flounder Paralichthys olivaceus vaccinated by immersion of formalin-inactivated Edwardsiella tarda following hyperosmotic treatment. Dis. Aquat. Organ. 2015, 116, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tang, X.Q.; Sheng, X.Z.; Xing, J.; Zhan, W.B. Immune response of flounder (Paralichthys olivaceus ) was associated with the concentration of inactivated Edwardsiella tarda, and immersion time. Vet. Immunol. Immunopathol. 2015, 167, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Falquet, L.; Pagni, M.; Bucher, P.; Hulo, N.; Sigrist, C.J.; Hofmann, K.; Bairoch, A. The PROSITE database, its status in 2002. Nucleic Acids Res. 2002, 30, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Tusnády, G.E.; Dosztányi, Z.; Simon, I. TMDET: Web server for detecting transmembrane regions of proteins by using their 3D coordinates. Bioinformatics 2005, 21, 1276–1277. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Jung, E.; Brunak, S. NetNGlyc 1.0 Server. Center for biological sequence analysis, technical university of Denmark. Available online: http://www.cbs.dtudk/services/NetNGlyc (accessed on 15 September 2016).
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ingerslev, H.C.; Pettersen, E.F.; Jakobsen, R.A.; Petersen, C.B.; Wergeland, H.I. Expression profiling and validation of reference gene candidates in immune relevant tissues and cells from Atlantic salmon (Salmo salar, L.). Mol. Immunol. 2006, 43, 1194–1201. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1. P. olivaceus | 2. D. rerio | 3. O. mykiss | 4. S. salar | 5. D. labrax | 6. B. diacanthus | 7. C. idella | 8. P. altivelis | |
|---|---|---|---|---|---|---|---|---|
| Constant regions (CH1–CH4) | ||||||||
| 1. P. olivaceus | ●22.3 | 43 | 42.1 | 42.2 | 52 | ●23.5 | ●25.4 | |
| 2. D. rerio | ●24.5 | ●23.2 | ●17.9 | ●19.3 | 36.3 | ●17.6 | ||
| 3. O. mykiss | 79.8 | ●31.6 | 39 | ●27.5 | 34.1 | |||
| 4. S. salar | 33.1 | 39.5 | ●26.2 | 33 | ||||
| 5. D. labrax | 44.3 | ●17.3 | ●17.7 | |||||
| 6. B. diacanthus | ●21.5 | ●25.8 | ||||||
| 7. C. idella | ●20.5 | |||||||
| 8. P. altivelis | ||||||||
| CH1-τ/ζ | ||||||||
| 1. P. olivaceus | ●23.2 | 43.1 | 43.0 | 48.9 | ●28.0 | 35.0 | ||
| 2. D. rerio | ●16.6 | ●17.0 | ●17.3 | ●18.8 | ●20.2 | |||
| 3. O. mykiss | 79.8 | 37.2 | 34.8 | 48.0 | ||||
| 4. S. salar | 39.1 | 36.5 | 47.4 | |||||
| 5. D. labrax | ||||||||
| 6. B. diacanthus | ●27.1 | 31.9 | ||||||
| 7. C. idella | 32.6 | |||||||
| 8. P. altivelis | ||||||||
| CH2-τ/ζ | ||||||||
| 1. P. olivaceus | ●23.7 | 40.6 | 36.4 | 51.0 | 52.0 | ●23.1 | 34.3 | |
| 2. D. rerio | 35.4 | 34.3 | ●27.8 | ●28.8 | ●16.1 | ●31.2 | ||
| 3. O. mykiss | 88.4 | 42.7 | 38.5 | ●19.4 | 42.1 | |||
| 4. S. salar | 42.7 | 36.4 | ●17.5 | 41.0 | ||||
| 5. D. labrax | 58.9 | ●19.4 | 34.3 | |||||
| 6. B. diacanthus | ●20.3 | 38.5 | ||||||
| 7. C. idella | ●20.3 | |||||||
| 8. P. altivelis | ||||||||
| CH3-τ/ζ | ||||||||
| 1. P. olivaceus | ●21.6 | 42.4 | 43.3 | 55.1 | 52.8 | ●18.1 | ||
| 2. D. rerio | ●22.6 | ●20.7 | ●23.3 | ●19.8 | 47.8 | |||
| 3. O. mykiss | 83.9 | 37.9 | 40.5 | ●21.5 | ||||
| 4. S. salar | 39.8 | 44.3 | ●22.4 | |||||
| 5. D. labrax | 54.2 | ●18.6 | ||||||
| 6. B. diacanthus | ●15.5 | |||||||
| 7. C. idella | ||||||||
| 8. P. altivelis | ||||||||
| CH4-τ/ζ | ||||||||
| 1. P. olivaceus | ●26.7 | 45.6 | 43.9 | 57.0 | 55.6 | ●24.1 | 36.7 | |
| 2. D. rerio | ●30.6 | ●27.9 | ●25.6 | ●17.7 | 52.3 | ●23.2 | ||
| 3. O. mykiss | 78.7 | 45.6 | 39.3 | 33.6 | 46.3 | |||
| 4. S. salar | 45.6 | 37.6 | ●31.8 | 43.6 | ||||
| 5. D. labrax | 51.3 | ●26.3 | 33.6 | |||||
| 6. B. diacanthus | ●19.4 | ●32.2 | ||||||
| 7. C. idella | ●30.3 | |||||||
| 8. P. altivelis | ||||||||
| τ1 | τ2 | τ3 | τ4 | μ1 | μ2 | μ3 | μ4 | δ1 | δ2 | δ3 | δ4 | δ5 | δ6 | δ7 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| τ1 | 19.1 | ●12.0 | ●13.2 | 38.0 | ●14.7 | ●11.9 | ●12.6 | ●13.2 | 18.2 | ●10.1 | ●15.2 | ●11.9 | ●13.7 | 17.6 | |
| τ2 | 16.0 | ●14.0 | 24.7 | 19.2 | ●8.10 | ●13.3 | 21.6 | ●12.3 | ●8.30 | ●13.5 | ●11.8 | 15.7 | 16.6 | ||
| τ3 | ●12.9 | ●11.1 | ●13.5 | ●12.7 | ●10.7 | ●11.1 | ●13.0 | ●12.7 | ●12.3 | ●14.2 | 16.0 | ●14.8 | |||
| τ4 | ●13.2 | ●9.20 | ●13.9 | ●12.8 | ●8.80 | ●10.6 | ●9.50 | ●14.1 | ●14.7 | ●14.2 | 16.8 | ||||
| μ1 | 15.6 | ●12.8 | ●13.4 | 16.0 | 19.2 | 16.6 | ●15.2 | ●14.6 | ●15.5 | ●14.7 | |||||
| μ2 | ●9.20 | ●10.6 | ●14.6 | ●10.2 | ●11.9 | ●14.1 | ●11.7 | ●11.3 | ●13.1 | ||||||
| μ3 | ●13.8 | ●10.0 | ●15.2 | ●11.2 | ●12.2 | ●13.7 | ●12.5 | ●11.9 | |||||||
| μ4 | ●13.2 | ●14.0 | ●12.4 | ●15.0 | 18.4 | 20.0 | ●15.0 | ||||||||
| δ1 | ●14.1 | ●12.8 | ●15.0 | ●12.8 | 18.3 | 17.9 | |||||||||
| δ2 | ●10.2 | ●11.5 | 17.4 | ●9.00 | ●11.5 | ||||||||||
| δ3 | 16.3 | ●14.6 | ●13.6 | ●10.3 | |||||||||||
| δ4 | ●13.7 | 19.6 | 17.4 | ||||||||||||
| δ5 | 16.0 | ●14.6 | |||||||||||||
| δ6 | 16.6 | ||||||||||||||
| δ7 |
| Primer Name | Oligonucleotide Sequence |
|---|---|
| Core segment PCR | |
| IgT-F | 5′-ACAGTGACACTGAAMCMWCCAAGT-3′ |
| IgT-R | 5′-GTYACMAGYGTYTWCACCATC-3′ |
| RACE PCR | |
| IgT 3′-RACE 1st | 5′-CCCTACTGACCTTTAAGCCA-3′ |
| IgT 3′-RACE nested | 5′-CGATCACCAATGTTCACTGC-3′ |
| IgT 5′-RACE 1st | 5′-AGGCAGCACAGGTAACAGGT-3′ |
| IgT 5′-RACE nested | 5′-TTGGTGTGTATGCTGAAAGAT-3′ |
| 3′-CDS | 5′-AAGCAGTGGTATCAACGCAGAGTAC(T)30VN-3′ |
| UPM | 5′-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3′ |
| NUP | 5′-AAGCAGTGGTATCAACGCAGAGT-3′ |
| 5-AP-DG | 5′-AAGCAGTGGTATCAACGCAGAGTACGCGGGGGGGGGG-3′ |
| Full-length validation | |
| qIgT-F | 5′-CCACCAACCAGACGTTCAAC-3′ |
| mIgT-R | 5′-TTGCTTCATCTTGACCAGGC-3′ |
| sIgT-R | 5′-GGCGTAACCCATACCCTTAG-3′ |
| RT-PCR | |
| IgT1-F | 5′-CGACTCTGTTCCCTTTGGTTC-3′ |
| IgT1-R | 5′-GTTTTCCTCCGTAAGTTGCCT-3′ |
| qRT-PCR | |
| IgT2-F | 5′-TAATTGTTCAGTAACTCATGCCG-3′ |
| IgT2-R | 5′-GATTGAAGTGTTCCTATGCGTCT-3′ |
| β-actin F | 5′-AGAGCAAGAGAGGCATCCTGAC-3′ |
| β-actin R | 5′-CGATGGGTGATGACCTGTCC-3′ |
| 18sRNA-F | 5′-GGTCTGTGATGCCCTTAGATGTC-3′ |
| 18sRNA-R | 5′-AGTGGGGTTCAGCGGGTTAC-3′ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Tang, X.; Zhan, W.; Xing, J.; Sheng, X. Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses. Int. J. Mol. Sci. 2016, 17, 1571. https://doi.org/10.3390/ijms17091571
Du Y, Tang X, Zhan W, Xing J, Sheng X. Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses. International Journal of Molecular Sciences. 2016; 17(9):1571. https://doi.org/10.3390/ijms17091571
Chicago/Turabian StyleDu, Yang, Xiaoqian Tang, Wenbin Zhan, Jing Xing, and Xiuzhen Sheng. 2016. "Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses" International Journal of Molecular Sciences 17, no. 9: 1571. https://doi.org/10.3390/ijms17091571
APA StyleDu, Y., Tang, X., Zhan, W., Xing, J., & Sheng, X. (2016). Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses. International Journal of Molecular Sciences, 17(9), 1571. https://doi.org/10.3390/ijms17091571