
In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development

 ,
,
Abstract
:
1. Introduction
2. Results
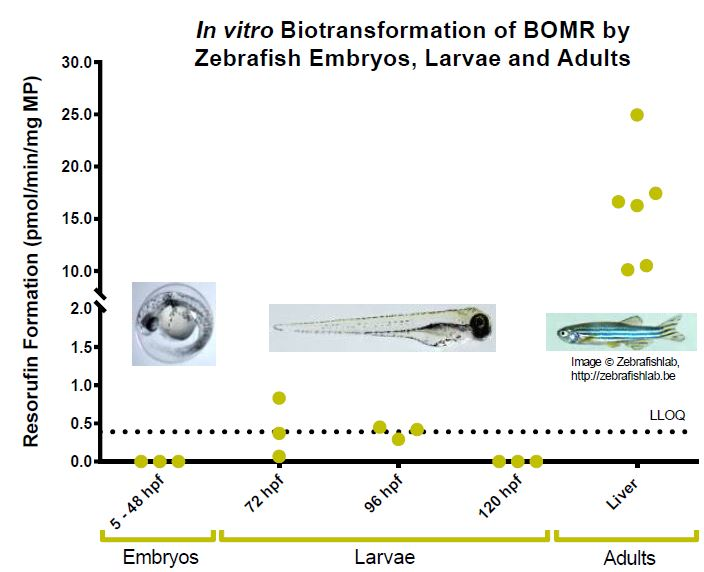
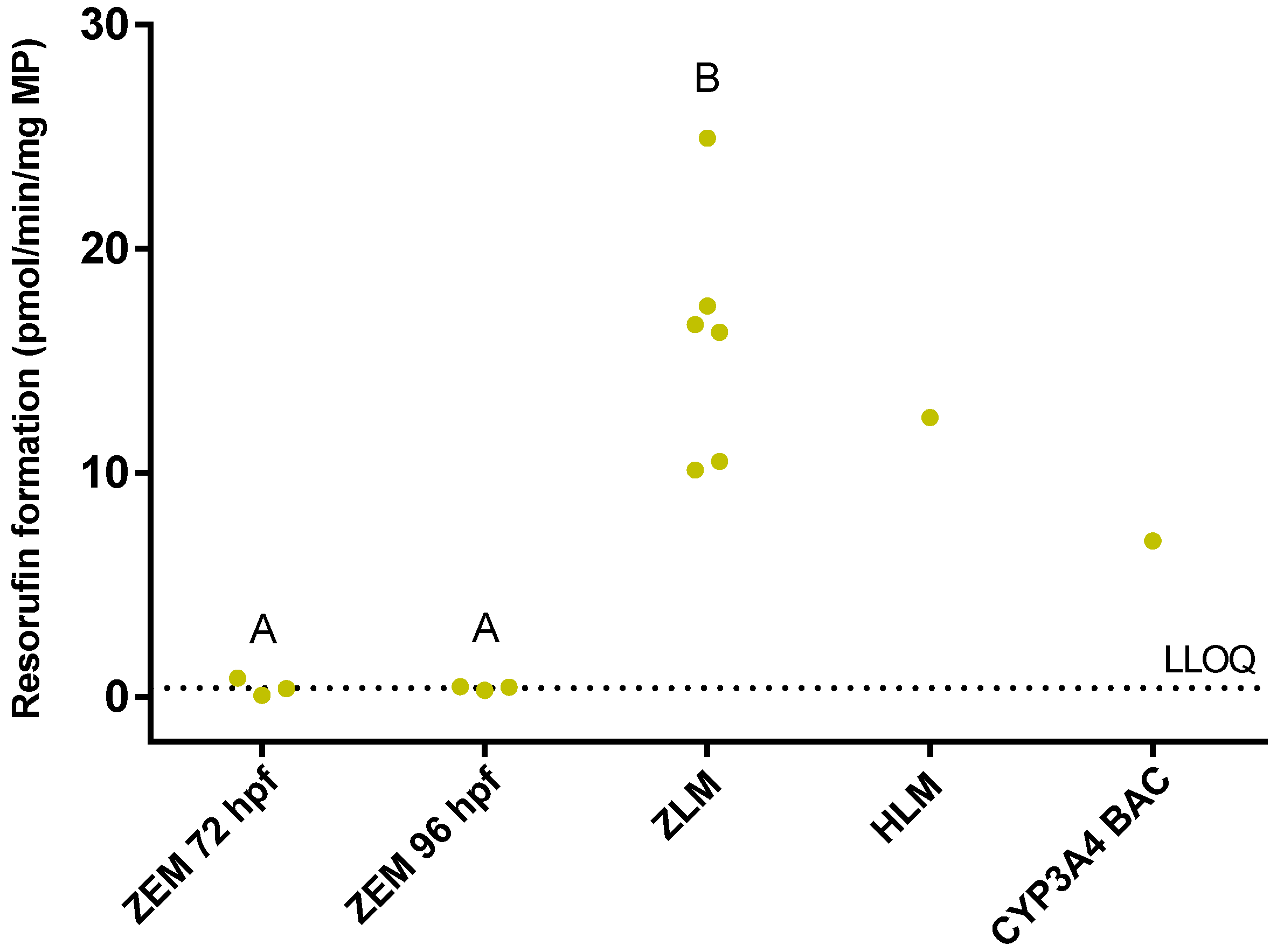
2.1. Benzyloxy-Methyl-Resorufin Assay in Adult Zebrafish Liver Microsomes and in Microsomes from Whole Zebrafish Embryo Homogenates
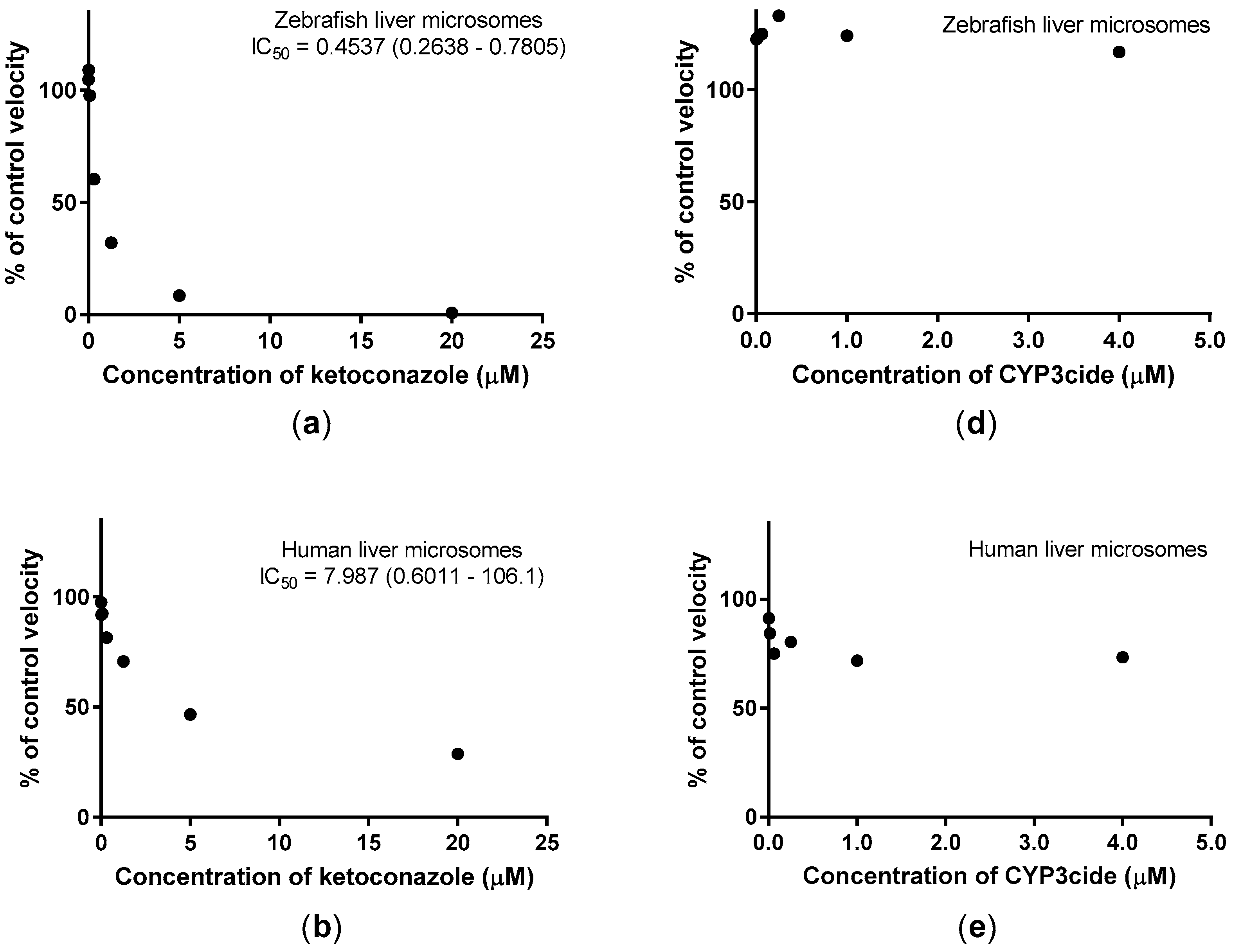
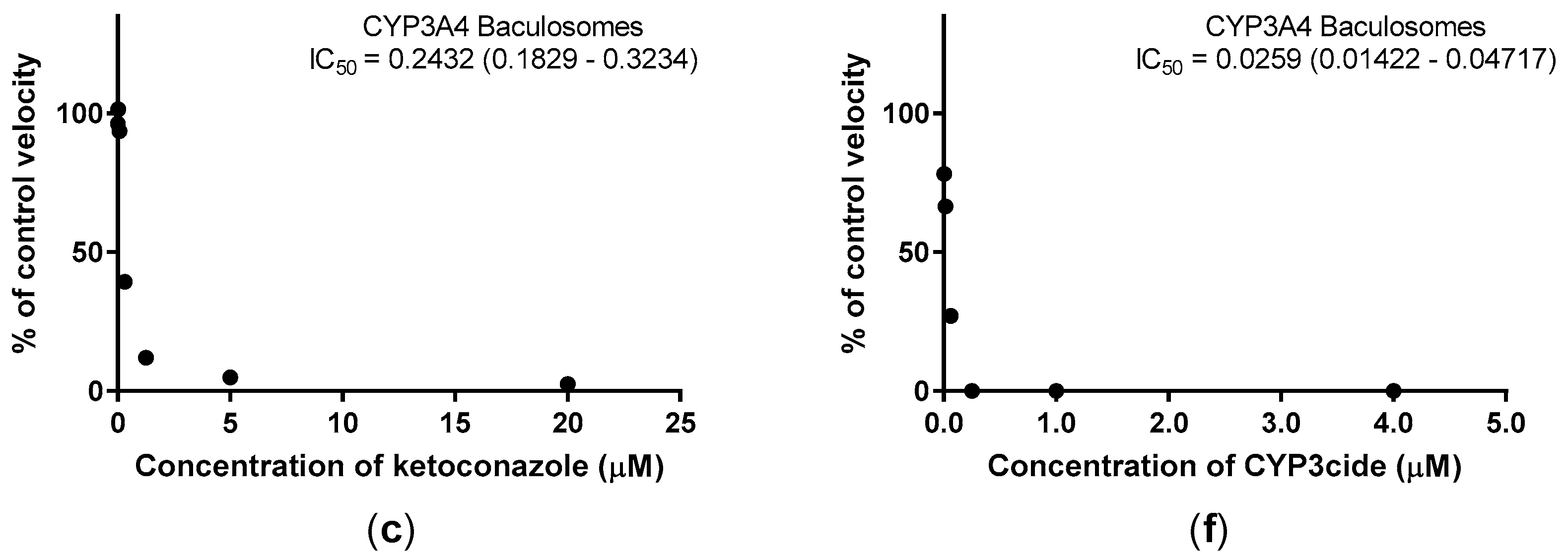
2.2. Inhibition Studies with Adult Zebrafish Liver Microsomes
2.3. Benzyloxy-Methyl-Resorufin Assay in Cytochrome P450 (CYP) Baculosomes® and in Recombinant Zebrafish CYPs
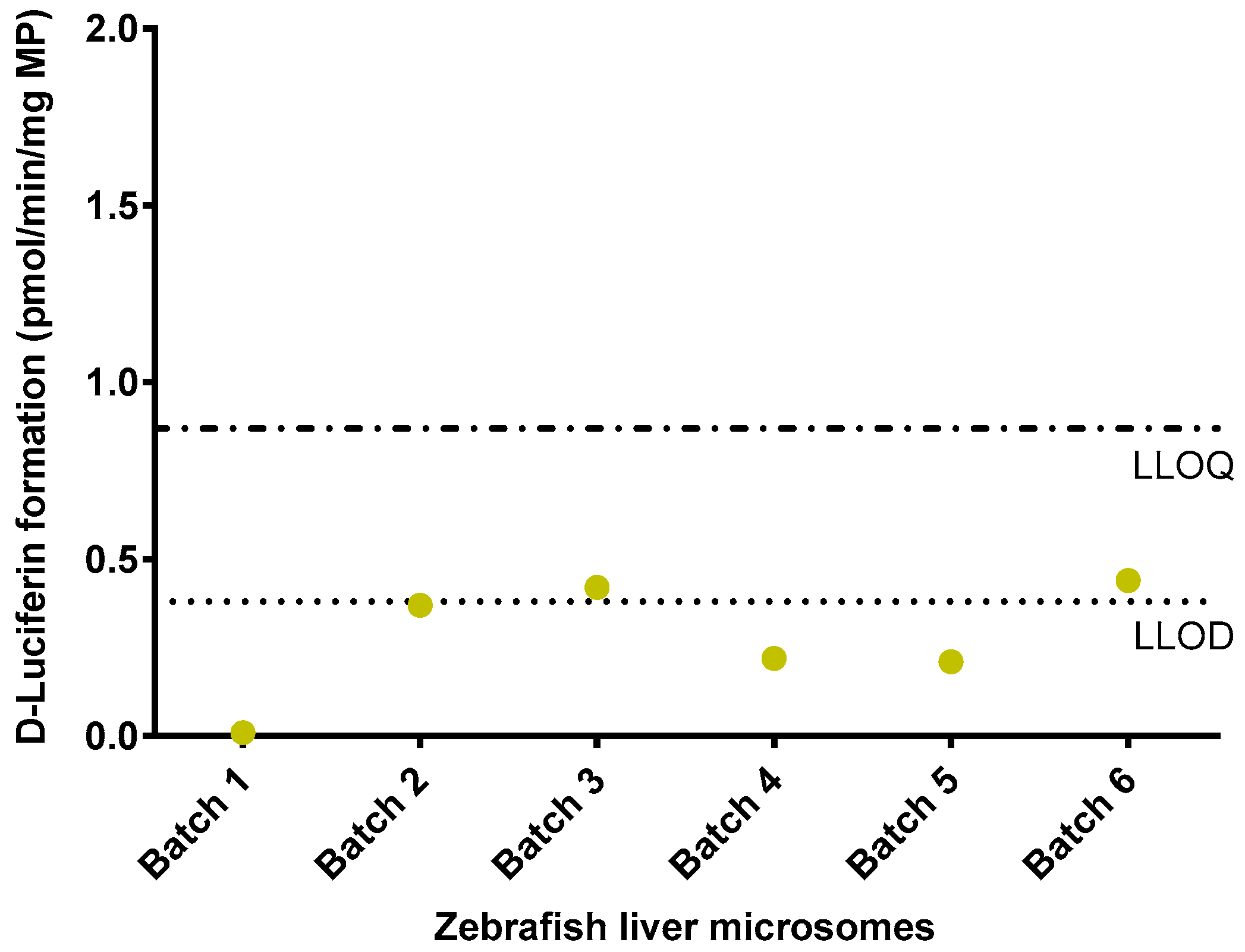
2.4. Luciferin-IPA Assay with Adult Zebrafish Liver Microsomes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fish Maintenance and Breeding
5.2. Tissue Sampling
5.2.1. Adult Zebrafish
5.2.2. Zebrafish Embryos
5.3. Isolation of Microsomes
5.3.1. Adult Zebrafish
5.3.2. Zebrafish Embryos
5.4. Benzyloxy-Methyl-Resorufin Assay in Adult Zebrafish Liver Microsomes
5.5. Benzyloxy-Methyl-Resorufin Assay in Microsomes from Whole Zebrafish Embryo Homogenates
5.6. Inhibition Studies with Adult Zebrafish Liver Microsomes
5.6.1. Ketoconazole
5.6.2. CYP3cide
5.7. Benzyloxy-Methyl-Resorufin Assay in CYP Baculosomes®
5.8. Benzyloxy-Methyl-Resorufin Assay in Recombinant Zebrafish CYPs
5.9. Luciferin-IPA Assay with Adult Zebrafish Liver Microsomes
5.10. Mathematical and Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barros, T.P.; Alderton, W.K.; Reynolds, H.M.; Roach, A.G.; Berghmans, S. Zebrafish: An emerging technology for in vivo pharmacological assessment to identify potential safety liabilities in early drug discovery. Br. J. Pharmacol. 2008, 154, 1400–1413. [Google Scholar] [CrossRef] [PubMed]
- Kari, G.; Rodeck, U.; Dicker, A.P. Zebrafish: An emerging model system for human disease and drug discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Lammer, E.; Carr, G.J.; Wendler, K.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T. Is the fish embryo toxicity test (FET) with the zebrafish (Danio rerio) a potential alternative for the fish acute toxicity test? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 196–209. [Google Scholar] [CrossRef] [PubMed]
- McGrath, P.; Li, C.Q. Zebrafish: A predictive model for assessing drug-induced toxicity. Drug Discov. Today 2008, 13, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Brachydanio Rerio), 5th ed.; M. Westerfield: Eugene, OR, USA, 1993; pp. 1.1, 2.1–2.8. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=314.108 (accessed on 19 September 2016).
- European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Strahle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Hines, R.N.; McCarver, D.G. The ontogeny of human drug-metabolizing enzymes: Phase I oxidative enzymes. J. Pharmacol. Exp. Ther. 2002, 300, 355–360. [Google Scholar] [CrossRef] [PubMed]
- de Wildt, S.N.; Kearns, G.L.; Leeder, J.S.; van den Anker, J.N. Glucuronidation in humans. Pharmacogenetic and developmental aspects. Clin. Pharmacokinet. 1999, 36, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Weigt, S.; Huebler, N.; Strecker, R.; Braunbeck, T.; Broschard, T.H. Zebrafish (Danio rerio) embryos as a model for testing proteratogens. Toxicology 2011, 281, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.W.; Kam, P.C. The physiological and pharmacological roles of cytochrome P450 isoenzymes. Anaesthesia 1999, 54, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450s and other enzymes in drug metabolism and toxicity. AAPS J. 2006, 8, E101–E111. [Google Scholar] [CrossRef] [PubMed]
- Martignoni, M.; Groothuis, G.M.M.; de Kanter, R. Species differences between mouse, rat, dog, monkey and human CYP–mediated drug metabolism, inhibition and induction. Expert Opin. Drug Metab. Toxicol. 2006, 2, 875–894. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.V.; McArthur, A.G.; Kubota, A.; Zanette, J.; Parente, T.; Jonsson, M.E.; Nelson, D.R.; Stegeman, J.J. Identification and developmental expression of the full complement of Cytochrome P450 genes in Zebrafish. BMC Genom. 2010, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Hegelund, T.; Cox, R.L.; Stegeman, J.J.; Liljenberg, M.; Olsson, U.; Sundberg, P.; Celander, M.C. Phylogenetic analysis of the cytochrome P450 3 (CYP3) gene family. J. Mol. Evol. 2003, 57, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Cai, Z. Molecular evolution and functional divergence of the cytochrome P450 3 (CYP3) Family in Actinopterygii (ray-finned fish). PLoS ONE 2010, 5, e14276. [Google Scholar] [CrossRef] [PubMed]
- Braunig, J.; Schiwy, S.; Broedel, O.; Muller, Y.; Frohme, M.; Hollert, H.; Keiter, S.H. Time–dependent expression and activity of cytochrome P450 1s in early life–stages of the zebrafish (Danio rerio). Environ. Sci. Pollut. Res. Int. 2015, 22, 16319–16328. [Google Scholar] [CrossRef] [PubMed]
- Chng, H.T.; Ho, H.K.; Yap, C.W.; Lam, S.H.; Chan, E.C. An investigation of the bioactivation potential and metabolism profile of Zebrafish versus human. J. Biomol. Screen. 2012, 17, 974–986. [Google Scholar] [CrossRef] [PubMed]
- Corley-Smith, G.E.; Su, H.T.; Wang-Buhler, J.L.; Tseng, H.P.; Hu, C.H.; Hoang, T.; Chung, W.G.; Buhler, D.R. CYP3C1, the first member of a new cytochrome P450 subfamily found in zebrafish (Danio rerio). Biochem. Biophys. Res. Commun. 2006, 340, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Creusot, N.; Brion, F.; Piccini, B.; Budzinski, H.; Porcher, J.M.; Ait-Aissa, S. BFCOD activity in fish cell lines and zebrafish embryos and its modulation by chemical ligands of human aryl hydrocarbon and nuclear receptors. Environ. Sci. Pollut. Res. Int. 2015, 22, 16393–16404. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Liu, K.; Wang, S.; Hou, H.; Yuan, Y.; Wang, X. Toxicity induced by emodin on zebrafish embryos. Drug Chem. Toxicol. 2012, 35, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.S.; Panter, G.H.; Hutchinson, T.H.; Chipman, J.K. Oxidative and conjugative xenobiotic metabolism in zebrafish larvae in vivo. Zebrafish 2010, 7, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, J.; Fang, Q.; Li, Y. Transcription alterations of microRNAs, cytochrome P4501A1 and 3A65, and AhR and PXR in the liver of zebrafish exposed to crude microcystins. Toxicon 2013, 73, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Nie, F.H.; Lin, H.Y.; Ma, Y.; Ju, X.H.; Chen, J.J.; Gooneratne, R. Developmental toxicity, EROD, and CYP1A mRNA expression in zebrafish embryos exposed to dioxin–like PCB126. Environ. Toxicol. 2016, 31, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Mattingly, C.J.; Toscano, W.A. Posttranscriptional silencing of cytochrome P4501A1 (CYP1A1) during zebrafish (Danio rerio) development. Dev. Dyn. 2001, 222, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.C.; Schmidt, A.D.; Hollert, H.; Braunbeck, T. Spatio–temporal development of CYP1 activity in early life–stages of zebrafish (Danio rerio). Aquat. Toxicol. 2010, 100, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.; Verbueken, E.; Pype, C.; Casteleyn, C.; Van Ginneken, C.; Maes, L.; Cos, P.; Van Cruchten, S. In vitro CYP1A activity in the zebrafish: Temporal but low metabolite levels during organogenesis and lack of gender differences in the adult stage. Reprod. Toxicol. 2016, 64, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Schiwy, S.; Braunig, J.; Alert, H.; Hollert, H.; Keiter, S.H. A novel contact assay for testing aryl hydrocarbon receptor (AhR)-mediated toxicity of chemicals and whole sediments in zebrafish (Danio rerio) embryos. Environ. Sci. Pollut. Res. Int. 2015, 22, 16305–16318. [Google Scholar] [CrossRef] [PubMed]
- Scornaienchi, M.L.; Thornton, C.; Willett, K.L.; Wilson, J.Y. Cytochrome P450-mediated 17β-estradiol metabolism in zebrafish (Danio rerio). J. Endocrinol. 2010, 206, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Scornaienchi, M.L.; Thornton, C.; Willett, K.L.; Wilson, J.Y. Functional differences in the cytochrome P450 1 family enzymes from zebrafish (Danio rerio) using heterologously expressed proteins. Arch. Biochem. Biophys. 2010, 502, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Shaya, L.; Dejong, C.; Wilson, J.Y. Expression patterns of cytochrome P450 3B and 3C genes in model fish species. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.P.; Hseu, T.H.; Buhler, D.R.; Wang, W.D.; Hu, C.H. Constitutive and xenobiotics-induced expression of a novel CYP3A gene from zebrafish larva. Toxicol. Appl. Pharmacol. 2005, 205, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Doshi, U.; Li, A.P. Luciferin IPA-based higher throughput human hepatocyte screening assays for CYP3A4 inhibition and induction. J. Biomol. Screen. 2011, 16, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Meisenheimer, P.L.; Uyeda, H.T.; Ma, D.; Sobol, M.; McDougall, M.G.; Corona, C.; Simpson, D.; Klaubert, D.H.; Cali, J.J. Proluciferin acetals as bioluminogenic substrates for cytochrome P450 activity and probes for CYP3A inhibition. Drug Metab. Disp. 2011, 39, 2403–2410. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, D.J.; Zhao, Y.; Venkatakrishnan, K.; Duan, S.X.; Harmatz, J.S.; Parent, S.J.; Court, M.H.; von Moltke, L.L. Mechanism of cytochrome P450–3A inhibition by ketoconazole. J. Pharm. Pharmacol. 2011, 63, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Walsky, R.L.; Obach, R.S.; Hyland, R.; Kang, P.; Zhou, S.; West, M.; Geoghegan, K.F.; Helal, C.J.; Walker, G.S.; Goosen, T.C.; et al. Selective mechanism-based inactivation of CYP3A4 by CYP3cide (PF-04981517) and its utility as an in vitro tool for delineating the relative roles of CYP3A4 versus CYP3A5 in the metabolism of drugs. Drug Metab. Dispos. 2012, 40, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, N.J.; Lechon, M.J.G.; Houston, J.B.; Hallifax, D.; Brown, H.S.; Maurel, P.; Kenna, J.G.; Gustavsson, L.; Lohmann, C.; Skonberg, C.; et al. Primary hepatocytes: Current understanding of the regulation of metabolic enzymes and transporter proteins, and pharmaceutical practice for the use of hepatocytes in metabolism, enzyme induction, transporter, clearance, and hepatotoxicity studies. Drug Metab. Rev. 2007, 39, 159–234. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Zgoda, V.G.; Arison, B.; Correia, M.A. Structure-function relationships of rat liver CYP3A9 to its human liver orthologs: Site–directed active site mutagenesis to a progesterone dihydroxylase. Arch. Biochem. Biophys. 2003, 409, 113–126. [Google Scholar] [CrossRef]
- Eagling, V.A.; Tjia, J.F.; Back, D.J. Differential selectivity of cytochrome P450 inhibitors against probe substrates in human and rat liver microsomes. Br. J. Clin. Pharmacol. 1998, 45, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Hegelund, T.; Ottosson, K.; Radinger, M.; Tomberg, P.; Celander, M.C. Effects of the antifungal imidazole ketoconazole on CYP1A and CYP3A in rainbow trout and killifish. Environ. Toxicol. Chem. 2004, 23, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.H.; Remley, M.J.; Raunig, D.; Vaz, A.D. In vitro drug interactions of cytochrome p450: An evaluation of fluorogenic to conventional substrates. Drug Metab. Dispos. 2003, 31, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Zlokarnik, G.; Volak, L.P.; Andrew, M.M.; Tran, C.; Drysdale, T.; Shah, U.; Cleveland, T.; Makings, L.R. Validation of Fluorogenic Vivid™ Probe Substrates for Analysis of CYP450-Drug Interactions. In Proceedings of the 10th North American ISSX Meeting, Indianapolis, IN, USA, 24–28 October 2000.
- Field, H.A.; Ober, E.A.; Roeser, T.; Stainier, D.Y. Formation of the digestive system in zebrafish. I. Liver morphogenesis. Dev. Biol. 2003, 253, 279–290. [Google Scholar] [CrossRef]
- Ober, E.A.; Field, H.A.; Stainier, D.Y. From endoderm formation to liver and pancreas development in zebrafish. Mech. Dev. 2003, 120, 5–18. [Google Scholar] [CrossRef]
- Ng, A.N.; de Jong-Curtain, T.A.; Mawdsley, D.J.; White, S.J.; Shin, J.; Appel, B.; Dong, P.D.; Stainier, D.Y.; Heath, J.K. Formation of the digestive system in zebrafish: III. Intestinal epithelium morphogenesis. Dev. Biol. 2005, 286, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Alderton, W.; Berghmans, S.; Butler, P.; Chassaing, H.; Fleming, A.; Golder, Z.; Richards, F.; Gardner, I. Accumulation and metabolism of drugs and CYP probe substrates in zebrafish larvae. Xenobiotica 2010, 40, 547–557. [Google Scholar] [CrossRef] [PubMed]
- European Union. Commission Recommendation of 18 June 2007 on guidelines for the accomodation and care of animals used for experimental and other scientific purposes (2007/526/EC). Off. J. Eur. Union 2007, 197, 89. [Google Scholar]
- Hill, J.R. In vitro Drug Metabolism Using Liver Microsomes. In Current Protocols in Pharmacology; Enna, S.J., Williams, M., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2003; Volume 2, pp. 7.8.1–7.8.11. [Google Scholar]
- Sengul, U. Comparing determination methods of detection and quantification limits for aflatoxin analysis in hazelnut. J. Food Drug Anal. 2016, 24, 56–62. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CYP Isoform | Content in Liver (% of Total CYP) | % of Drugs Metabolized by CYP |
|---|---|---|
| CYP3A4/5 | ±30 | ±50 |
| CYP2D6 | ±4 | ±30 |
| CYP2B6 | 2–10 | ±25 |
| CYP2C8, -2C9, -2C19 | ±20 | ±16 |
| CYP1A2 | ±13 | ±4 |
| Recombinant CYPs | Resorufin Formation |
|---|---|
| CYP Baculosomes® 1 | pmol/min/pmol CYP |
| CYP1A2 BAC® | 0.519 ± 0.100 |
| CYP2B6 BAC® | 0.497 ± 0.133 |
| CYP3A4 BAC® | 0.133 ± 0.070 |
| CYP2C9 BAC® | 0.026 ± 0.002 |
| CYP2C19 BAC® | <LLOQ |
| CYP2D6 BAC® | <LLOQ |
| Recombinant Zebrafish CYPs 1 | pmol/min/µg Total Protein |
| CYP1A | 1.152 ± 0.068 |
| CYP1B | 0.105 ± 0.008 |
| CYP1C2 | 0.078 ± 0.011 |
| CYP1C1 | 0.004 ± 0.001 |
| CYP1D | <LLOQ |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verbueken, E.; Alsop, D.; Saad, M.A.; Pype, C.; Van Peer, E.M.; Casteleyn, C.R.; Van Ginneken, C.J.; Wilson, J.; Van Cruchten, S.J. In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development. Int. J. Mol. Sci. 2017, 18, 217. https://doi.org/10.3390/ijms18010217
Verbueken E, Alsop D, Saad MA, Pype C, Van Peer EM, Casteleyn CR, Van Ginneken CJ, Wilson J, Van Cruchten SJ. In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development. International Journal of Molecular Sciences. 2017; 18(1):217. https://doi.org/10.3390/ijms18010217
Chicago/Turabian StyleVerbueken, Evy, Derek Alsop, Moayad A. Saad, Casper Pype, Els M. Van Peer, Christophe R. Casteleyn, Chris J. Van Ginneken, Joanna Wilson, and Steven J. Van Cruchten. 2017. "In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development" International Journal of Molecular Sciences 18, no. 1: 217. https://doi.org/10.3390/ijms18010217
APA StyleVerbueken, E., Alsop, D., Saad, M. A., Pype, C., Van Peer, E. M., Casteleyn, C. R., Van Ginneken, C. J., Wilson, J., & Van Cruchten, S. J. (2017). In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development. International Journal of Molecular Sciences, 18(1), 217. https://doi.org/10.3390/ijms18010217