
Preferential Enhancement of Sensory and Motor Axon Regeneration by Combining Extracellular Matrix Components with Neurotrophic Factors

 , and
, and
Abstract
:1. Introduction
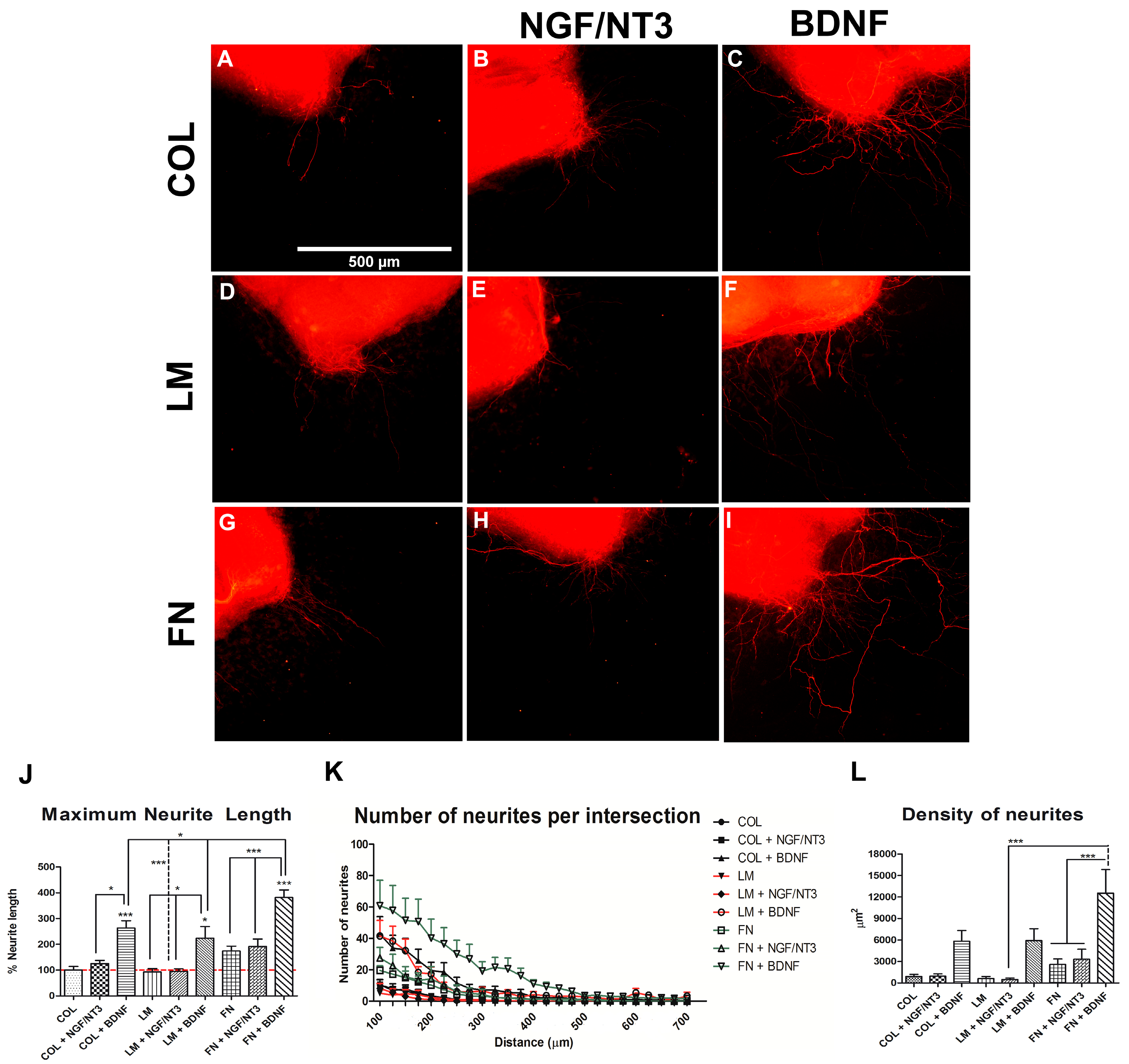
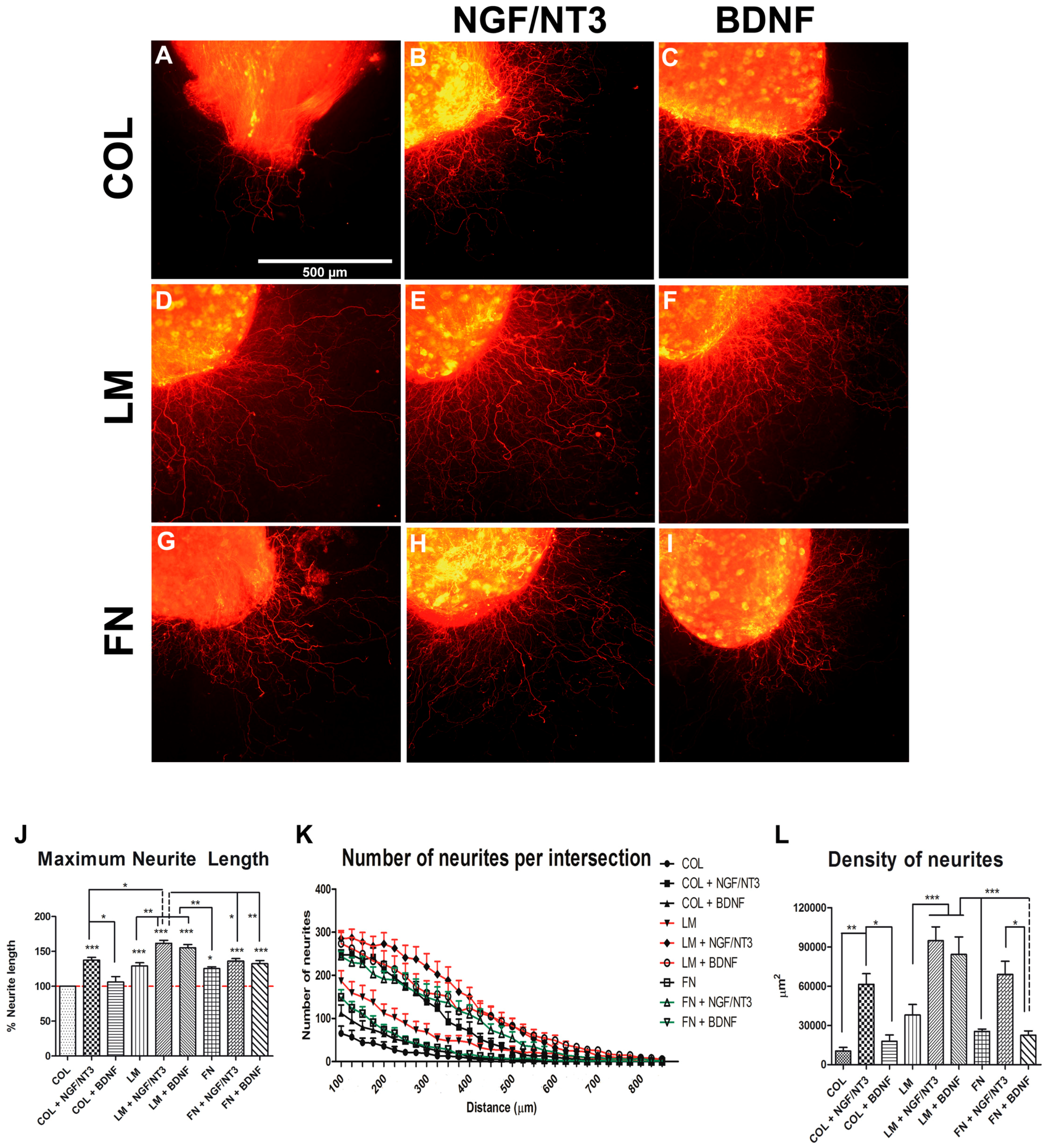
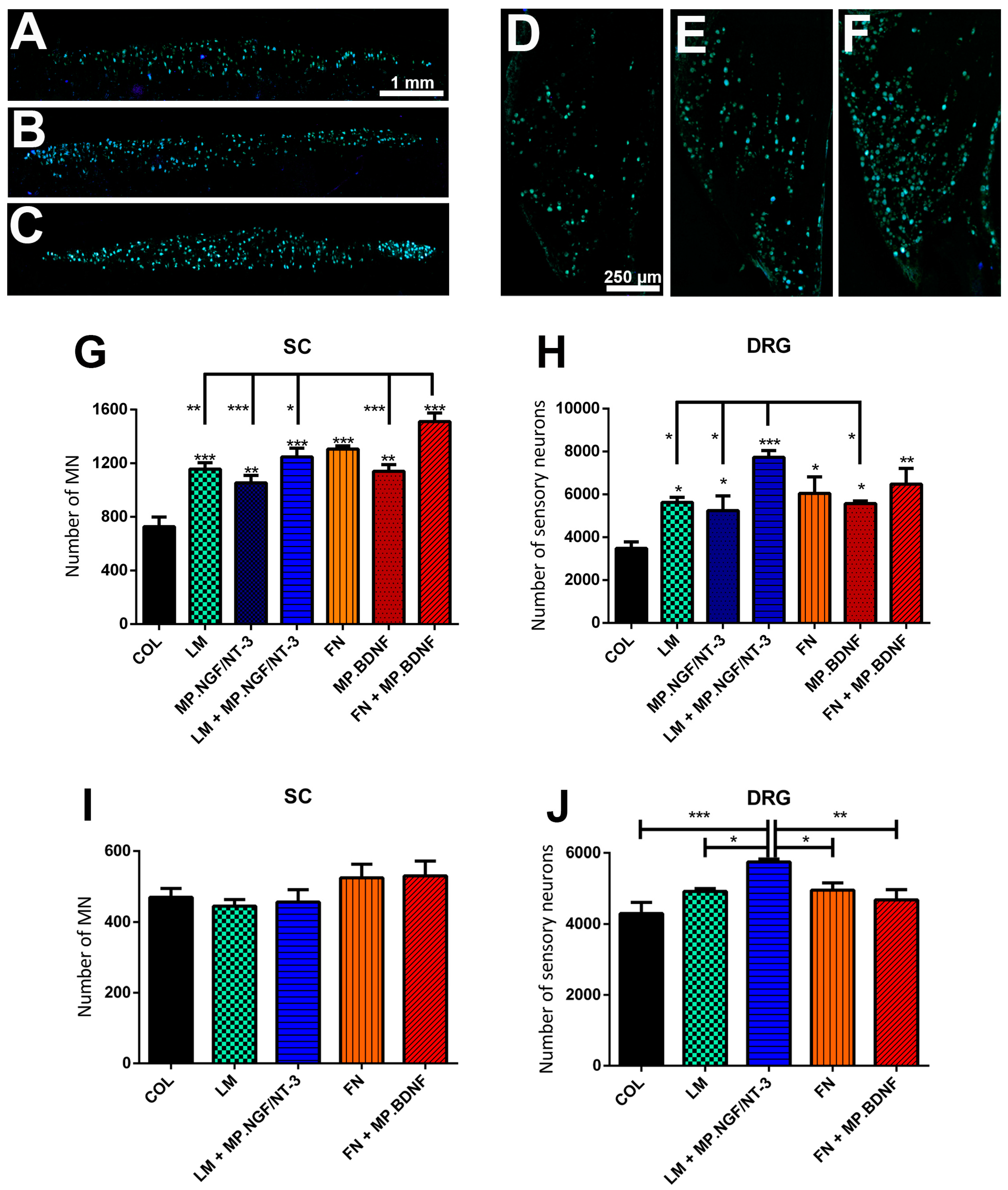
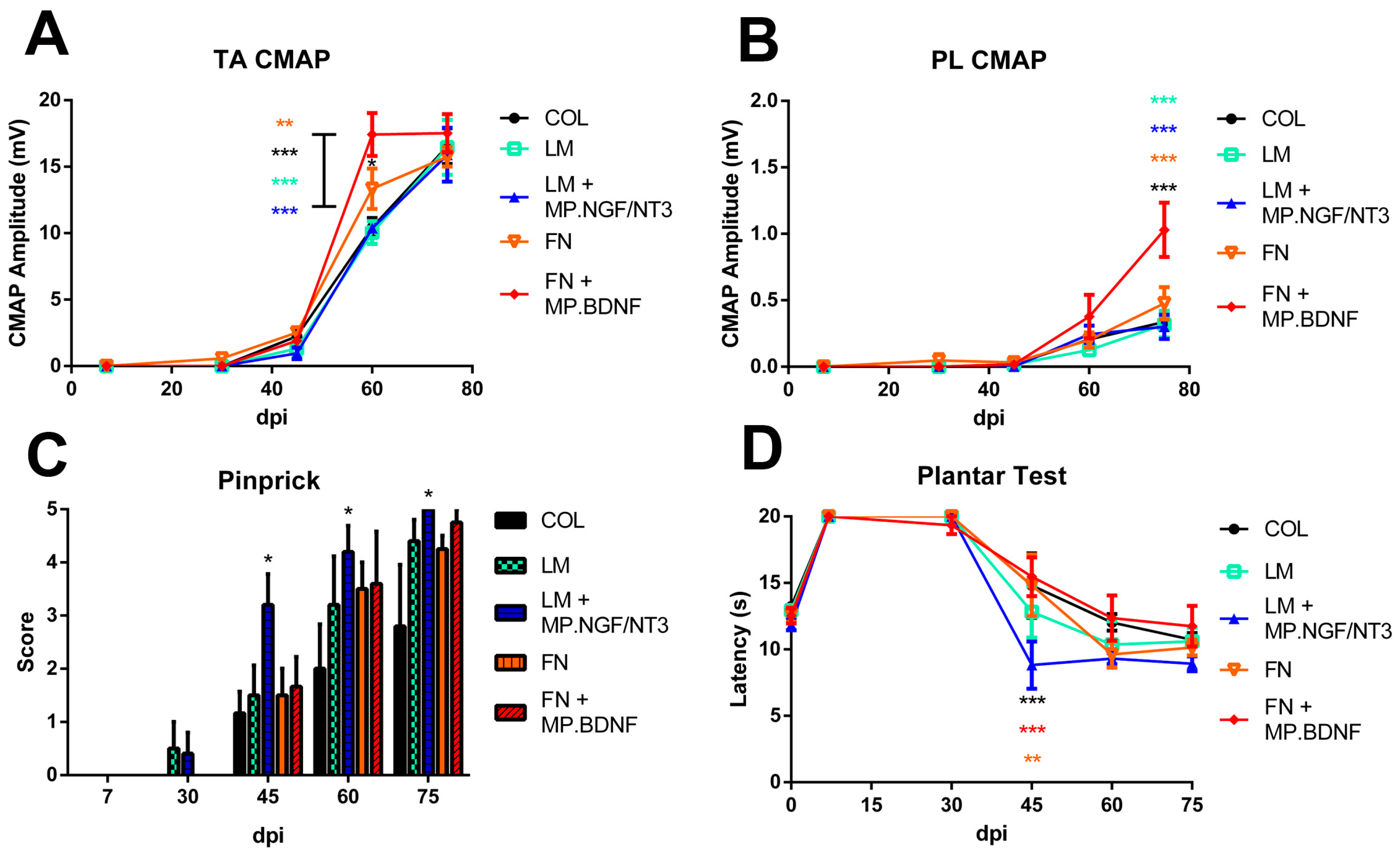
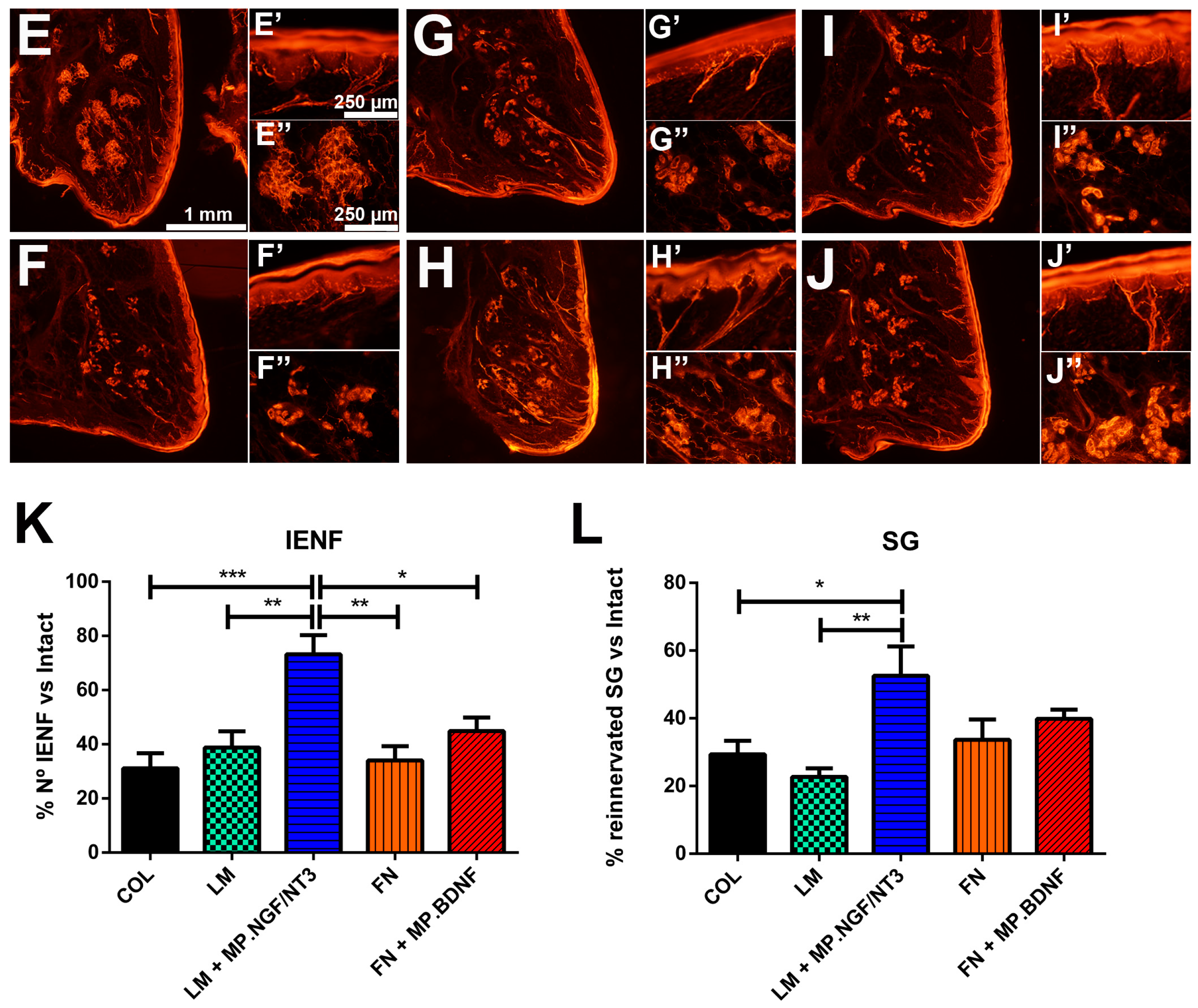
2. Results
2.1. In Vitro Effects of Combining Neurotrophic Factors (NTFs) and Extracellular Matrix (ECM) Substrates on Neurite Outgrowth
2.2. In Vivo Effects of Combination of NTF and ECM Substrates on Nerve Regeneration
2.3. Combination of FN and BDNF Promotes Motor Functional Recovery at Long Term
2.4. Combination of LM and NGF/NT3 Promotes Sensory Functional Recovery at Long Term
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. In Vitro Study on Organotypic Cultures
4.3. In Vivo Study of Peripheral Nerve Regeneration
4.4. Retrograde Labeling and Neuronal Counting
4.5. Assessment of Muscle Reinnervation
4.6. Assessment of Skin Nociceptive Reinnervation
4.7. Evaluation of Skin and Sweat Gland Reinnervation
4.8. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hall, S. The response to injury in the peripheral nervous system. J. Bone Jt. Surg. Br. 2005, 87, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; Miao, T.; Wu, D.; Zhang, Y.; Yeh, J.; Bo, X. Responses of the nerve cell body to axotomy. Neurosurgery 2009, 65, A74–A79. [Google Scholar] [CrossRef] [PubMed]
- Navarro, X.; Vivó, M.; Valero-Cabré, A. Neural plasticity after peripheral nerve injury and regeneration. Prog. Neurobiol. 2007, 82, 163–201. [Google Scholar] [CrossRef] [PubMed]
- Aldskogius, H.; Molander, C. Specificity in regenerative outgrowth and target reinnervation by mammalian peripheral axons. Restor. Neurol. Neurosci. 1990, 1, 275–280. [Google Scholar] [PubMed]
- Lundborg, G. Nerve injury and repair—A challenge to the plastic brain. Peripher. Nerv. Syst. 2003, 8, 209–226. [Google Scholar] [CrossRef]
- Valero-Cabré, A.; Navarro, X. Functional impact of axonal misdirection after peripheral nerve injuries followed by graft or tube repair. J. Neurotrauma 2002, 19, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Valero-Cabré, A.; Tsironis, K.; Skouras, E.; Navarro, X.; Neiss, W.F. Peripheral and spinal motor reorganization after nerve injury and repair. J. Neurotrauma 2004, 21, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Clements, I.P.; Mukhatyar, V.J.; Srinivasan, A.; Bentley, J.T.; Andreasen, D.S.; Bellamkonda, R.V. Regenerative scaffold electrodes for peripheral nerve interfacing. IEEE Trans. Neural Syst. Rehabil. Eng. 2013, 21, 554–566. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, P.; Garde, K.; Chouhan, A.; Bengali, E.; Romero-Ortega, M. Modality-specific axonal regeneration: Toward selective regenerative neural interfaces. Front. Neuroeng. 2011, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Allodi, I.; Udina, E.; Navarro, X. Specificity of peripheral nerve regeneration: Interactions at the axon level. Prog. Neurobiol. 2012, 98, 16–37. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, N.J. Integrins and the extracellular matrix: Key mediators of development and regeneration of the sensory nervous system. Dev. Neurobiol. 2011, 71, 1054–1072. [Google Scholar] [CrossRef] [PubMed]
- Lemons, M.L.; Condic, M.L. Integrin signaling is integral to regeneration. Exp. Neurol. 2008, 209, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Plantman, S.; Patarroyo, M.; Fried, K.; Domogatskaya, A.; Tryggvason, K.; Hammarberg, H.; Cullheim, S. Integrin-laminin interactions controlling neurite outgrowth from adult DRG neurons in vitro. Mol. Cell. Neurosci. 2008, 39, 50–62. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, F.; Alé, A.; Santos, D.; Barwig, C.; Freier, T.; Navarro, X.; Udina, E. Substratum preferences of motor and sensory neurons in postnatal and adult rats. Eur. J. Neurosci. 2016, 43, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Allodi, I.; Guzmán-Lenis, M.-S.; Hernàndez, J.; Navarro, X.; Udina, E. In vitro comparison of motor and sensory neuron outgrowth in a 3D collagen matrix. J. Neurosci. Methods 2011, 198, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Brushart, T.M.; Aspalter, M.; Griffin, J.W.; Redett, R.; Hameed, H.; Zhou, C.; Wright, M.; Vyas, A.; Höke, A. Schwann cell phenotype is regulated by axon modality and central-peripheral location, and persists in vitro. Exp. Neurol. 2013, 247, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Höke, A.; Redett, R.; Hameed, H.; Jari, R.; Zhou, C.; Li, Z.B.; Griffin, J.W.; Brushart, T.M. Schwann cells express motor and sensory phenotypes that regulate axon regeneration. J. Neurosci. 2006, 26, 9646–9655. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.A.; Madison, R.D. Motor neurons can preferentially reinnervate cutaneous pathways. Exp. Neurol. 2004, 190, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Martini, R.; Schachner, M.; Brushart, T.M. The L2/HNK-1 carbohydrate is preferentially expressed by previously motor axon-associated Schwann cells in reinnervated peripheral nerves. J. Neurosci. 1994, 14, 7180–7191. [Google Scholar] [PubMed]
- Madl, C.M.; Heilshorn, S.C. Matrix interactions modulate neurotrophin-mediated neurite outgrowth and pathfinding. Neural Regen. Res. 2015, 10, 514–517. [Google Scholar] [PubMed]
- Santos, D.; Giudetti, G.; Micera, S.; Navarro, X.; del Valle, J. Focal release of neurotrophic factors by biodegradable microspheres enhance motor and sensory axonal regeneration in vitro and in vivo. Brain Res. 2016, 1636, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Valero-Cabré, A.; Tsironis, K.; Skouras, E.; Perego, G.; Navarro, X.; Neiss, W.F. Superior muscle reinnervation after autologous nerve graft or poly-l-lactide-ε-caprolactone (PLC) tube implantation in comparison to silicone tube repair. J. Neurosci. Res. 2001, 63, 214–223. [Google Scholar] [CrossRef]
- Gordon, T. The role of neurotrophic factors in nerve regeneration. Neurosurg. Focus 2009, 26, E3. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.; Yu, V.M.; Lowe, J.B.; Brenner, M.J.; Hunter, D.A.; Mackinnon, S.E.; Sakiyama-Elbert, S.E. Controlled release of nerve growth factor enhances sciatic nerve regeneration. Exp. Neurol. 2003, 184, 295–303. [Google Scholar] [CrossRef]
- Kemp, S.W.P.; Webb, A.A.; Dhaliwal, S.; Syed, S.; Walsh, S.K.; Midha, R. Dose and duration of nerve growth factor (NGF) administration determine the extent of behavioral recovery following peripheral nerve injury in the rat. Exp. Neurol. 2011, 229, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Vögelin, E.; Baker, J.M.; Gates, J.; Dixit, V.; Constantinescu, M.A.; Jones, N.F. Effects of local continuous release of brain derived neurotrophic factor (BDNF) on peripheral nerve regeneration in a rat model. Exp. Neurol. 2006, 199, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Sterne, G.D.; Brown, R.A.; Green, C.J.; Terenghi, G. Neurotrophin-3 delivered locally via fibronectin mats enhances peripheral nerve regeneration. Eur. J. Neurosci. 1997, 9, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Ding, N.; Xing, Z.; Zhang, W.; Liu, H.; Li, Z. Insulin-like growth factor-1 regulates neurite outgrowth and neuronal migration from organotypic cultured dorsal root ganglion. Int. J. Neurosci. 2011, 121, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Levi-Montalcini, R. A nerve growth-stimulating factor isolated from snake venom. Proc. Natl. Acad. Sci. USA 1956, 42, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Al-Ali, H.; Beckerman, S.; Bixby, J.L.; Lemmon, V.P. In vitro models of axon regeneration. Exp. Neurol. 2017, 287, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Allodi, I.; Casals-Díaz, L.; Santos-Nogueira, E.; González-Pérez, F.; Navarro, X.; Udina, E. FGF-2 low molecular weight selectively promotes neuritogenesis of motor neurons in vitro. Mol. Neurobiol. 2013, 47, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Torres-Espín, A.; Allodi, I.; Santos, D.; González-Pérez, F.; Udina, E.; del Valle, J.; Navarro, X. Analysis of axonal growth in organotypic neural cultures. Protoc. Exch. 2016. [Google Scholar] [CrossRef]
- Hari, A.; Djohar, B.; Skutella, T.; Montazeri, S. Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. Int. J. Dev. Neurosci. 2004, 22, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Willem, M.; Jones, L.L.; Kreutzberg, G.W.; Mayer, U. Impaired axonal regeneration in α7 integrin-deficient mice. J. Neurosci. 2000, 20, 1822–1830. [Google Scholar] [PubMed]
- Karchewski, L.A.; Kim, F.A.; Johnston, J.; McKnight, R.M.; Verge, V.M.K. Anatomical evidence supporting the potential for modulation by multiple neurotrophins in the majority of adult lumbar sensory neurons. J. Comp. Neurol. 1999, 413, 327–341. [Google Scholar] [CrossRef]
- Gardiner, N.J.; Fernyhough, P.; Tomlinson, D.R.; Mayer, U.; von der Mark, H.; Streuli, C.H. α7 Integrin mediates neurite outgrowth of distinct populations of adult sensory neurons. Mol. Cell. Neurosci. 2005, 28, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Tucker, B.A.; Mearow, K.M. Peripheral sensory axon growth: From receptor binding to cellular signaling. Can. J. Neurol. Sci. 2008, 35, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Condic, M.L. Adult neuronal regeneration induced by transgenic integrin expression. J. Neurosci. 2001, 21, 4782–4788. [Google Scholar] [PubMed]
- Grabham, P.W.; Foley, M.; Umeojiako, A.; Goldberg, D.J. Nerve growth factor stimulates coupling of β1 integrin to distinct transport mechanisms in the filopodia of growth cones. J. Cell Sci. 2000, 113, 3003–3012. [Google Scholar] [PubMed]
- Filbin, M.T. Recapitulate development to promote axonal regeneration: Good or bad approach? Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.A. The manufacturing techniques of various drug loaded biodegradable poly (lactide-co-glycolide) (PLGA) devices. Biomaterials 2000, 21, 2475–2490. [Google Scholar] [CrossRef]
- Tannemaat, M.R.; Eggers, R.; Hendriks, W.T.; de Ruiter, G.C.W.; van Heerikhuize, J.J.; Pool, C.W.; Malessy, M.J.A.; Boer, G.J.; Verhaagen, J. Differential effects of lentiviral vector-mediated overexpression of nerve growth factor and glial cell line-derived neurotrophic factor on regenerating sensory and motor axons in the transected peripheral nerve. Eur. J. Neurosci. 2008, 28, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, L.; Delcroix, J.-D.; Fernyhough, P.; Tomlinson, D.R. Focally administered nerve growth factor suppresses molecular regenerative responses of axotomized peripheral afferents in rats. Neuroscience 1999, 91, 265–271. [Google Scholar] [CrossRef]
- González-Pérez, F.; Udina, E.; Navarro, X. Extracellular matrix components in peripheral nerve regeneration. Int. Rev. Neurobiol. 2013, 108, 257–275. [Google Scholar] [PubMed]
- Jesuraj, N.J.; Nguyen, P.K.; Wood, M.D.; Moore, A.M.; Borschel, G.H.; Mackinnon, S.E.; Sakiyama-Elbert, S.E. Differential gene expression in motor and sensory Schwann cells in the rat femoral nerve. J. Neurosci. Res. 2012, 90, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.; Gordon, T. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury. Mol. Neurobiol. 2003, 27, 277–324. [Google Scholar] [CrossRef]
- Navarro, X. Functional evaluation of peripheral nerve regeneration and target reinnervation in animal models: A critical overview. Eur. J. Neurosci. 2016, 43, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.; Tyreman, N.; Raji, M.A. The basis for diminished functional recovery after delayed peripheral nerve repair. J. Neurosci. 2011, 31, 5325–5334. [Google Scholar] [CrossRef] [PubMed]
- Levi-Montalcini, R. The nerve growth factor: Thirty-five years later. Biosci. Rep. 1987, 7, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Torres-Espín, A.; Santos, D.; González-Pérez, F.; del Valle, J.; Navarro, X. Neurite-J: An Image-J plug-in for axonal growth analysis in organotypic cultures. J. Neurosci. Methods 2014, 236, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Giudetti, G.; del Valle Macia, J.; Acebes, X.N.; Micera, S. NGF-loaded PLGA microparticles for advanced multifunctional regenerative electrodes. In Proceedings of the 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, EMBC 2014, Chicago, IL, USA, 26–30 August 2014; pp. 1993–1995.
- Verdú, E.; Labrador, R.O.; Rodríguez, F.J.; Ceballos, D.; Forés, J.; Navarro, X. Alignment of collagen and laminin-containing gels improve nerve regeneration within silicone tubes. Restor. Neurol. Neurosci. 2002, 20, 169–179. [Google Scholar] [PubMed]
- Santos, D.; González-Pérez, F.; Navarro, X.; del Valle, J. Dose-dependent differential effect of neurotrophic factors on in vitro and in vivo regeneration of motor and sensory neurons. Neural Plast. 2016, 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J. Stereology of arbitrary particles: A review of unbiased number and size estimators and the presentation of some new ones, in memory of William R. Thompson. J. Microsc. 1986, 143, 3–45. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Wieringa, P.; Moroni, L.; Navarro, X.; del Valle, J. PEOT/PBT guides enhance nerve regeneration in long gap defects. Adv. Healthc. Mater. 2016. [Google Scholar] [CrossRef] [PubMed]
- Navarro, X.; Verdú, E.; Butí, M. Comparison of regenerative and reinnervating capabilities of different functional types of nerve fibers. Exp. Neurol. 1994, 129, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Garcia, X.; del Valle, J.; Escribano, E.; Domenech, J.; Queralt, J. Analgesic and antiallodynic effects of antidepressants after infiltration into the rat. Pharmacology 2010, 86, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Navarro, X.; Verdú, E.; Wendelscafer-Crabb, G.; Kennedy, W.R. Innervation of cutaneous structures in the mouse hind paw: A confocal microscopy immunohistochemical study. J. Neurosci. Res. 1995, 41, 111–120. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Abbreviation | N | Description |
|---|---|---|---|
| In Vitro Condition | |||
| Collagen | COL | 8 | Collagen type I (3 mg/mL) gel |
| Collagen + NGF/NT3 | COL + NGF/NT3 | 7 | Collagen type I (3 mg/mL) gel supplemented with NGF and NT3 (25 + 25 ng/mL) |
| Collagen + BDNF | COL + BDNF | 6 | Collagen type I (3 mg/mL) gel supplemented with BDNF (50 ng/mL) |
| Laminin | LM | 8 | Collagen type I (3 mg/mL) gel containing 20% laminin type I |
| Laminin + NGF/NT3 | LM + NGF/NT3 | 7 | Collagen type I (3 mg/mL) gel containing 20% laminin type I and NGF + NT3 (25 + 25 ng/mL) |
| Laminin + BDNF | LM + BDNF | 7 | Collagen type I (3 mg/mL) gel containing 20% laminin type I and BDNF (50 ng/mL) |
| Fibronectin | FN | 6 | Collagen type I (3 mg/mL) gel containing 20% fibronectin |
| Fibronectin + NGF/NT-3 | FN + NGF/NT3 | 6 | Collagen type I (3 mg/mL) gel containing 20% fibronectin and NGF + NT3 (25 + 25 ng/mL) |
| Fibronectin + BDNF | FN + BDNF | 7 | Collagen type I (3 mg/mL) gel containing 20% fibronectin and BDNF (50 ng/mL) |
| In Vivo Condition | |||
| Collagen | COL | 6 | Collagen type I (3 mg/mL) gel |
| Laminin | LM | 6 | Collagen type I (3 mg/mL) gel containing 20% laminin type I |
| Collagen + NGF/NT3 * | MP.NGF/NT3 | 6 | Collagen type I (3 mg/mL) gel containing NGF + NT3 (1 + 1 µg/mL) encapsulated in PLGA microspheres |
| Laminin + NGF/NT3 | LM + MP.NGF/NT3 | 6 | Collagen type I (3 mg/mL) gel containing 20% laminin type I and NGF + NT3 (1 + 1 µg/mL) encapsulated in PLGA microspheres |
| Fibronectin | FN | 6 | Collagen type I (3 mg/mL) gel containing 20% fibronectin |
| Collagen + BDNF * | MP.BDNF | 6 | Collagen type I (3 mg/mL) gel containing 2 µg/mL of BDNF encapsulated in PLGA microspheres |
| Fibronectin + BDNF | FN + MP.BDNF | 6 | Collagen type I (3 mg/mL) gel containing 20% fibronectin and 2 µg/ml of BDNF encapsulated in PLGA microspheres |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, D.; González-Pérez, F.; Giudetti, G.; Micera, S.; Udina, E.; Del Valle, J.; Navarro, X. Preferential Enhancement of Sensory and Motor Axon Regeneration by Combining Extracellular Matrix Components with Neurotrophic Factors. Int. J. Mol. Sci. 2017, 18, 65. https://doi.org/10.3390/ijms18010065
Santos D, González-Pérez F, Giudetti G, Micera S, Udina E, Del Valle J, Navarro X. Preferential Enhancement of Sensory and Motor Axon Regeneration by Combining Extracellular Matrix Components with Neurotrophic Factors. International Journal of Molecular Sciences. 2017; 18(1):65. https://doi.org/10.3390/ijms18010065
Chicago/Turabian StyleSantos, Daniel, Francisco González-Pérez, Guido Giudetti, Silvestro Micera, Esther Udina, Jaume Del Valle, and Xavier Navarro. 2017. "Preferential Enhancement of Sensory and Motor Axon Regeneration by Combining Extracellular Matrix Components with Neurotrophic Factors" International Journal of Molecular Sciences 18, no. 1: 65. https://doi.org/10.3390/ijms18010065
APA StyleSantos, D., González-Pérez, F., Giudetti, G., Micera, S., Udina, E., Del Valle, J., & Navarro, X. (2017). Preferential Enhancement of Sensory and Motor Axon Regeneration by Combining Extracellular Matrix Components with Neurotrophic Factors. International Journal of Molecular Sciences, 18(1), 65. https://doi.org/10.3390/ijms18010065