
Sailuotong Prevents Hydrogen Peroxide (H2O2)-Induced Injury in EA.hy926 Cells



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
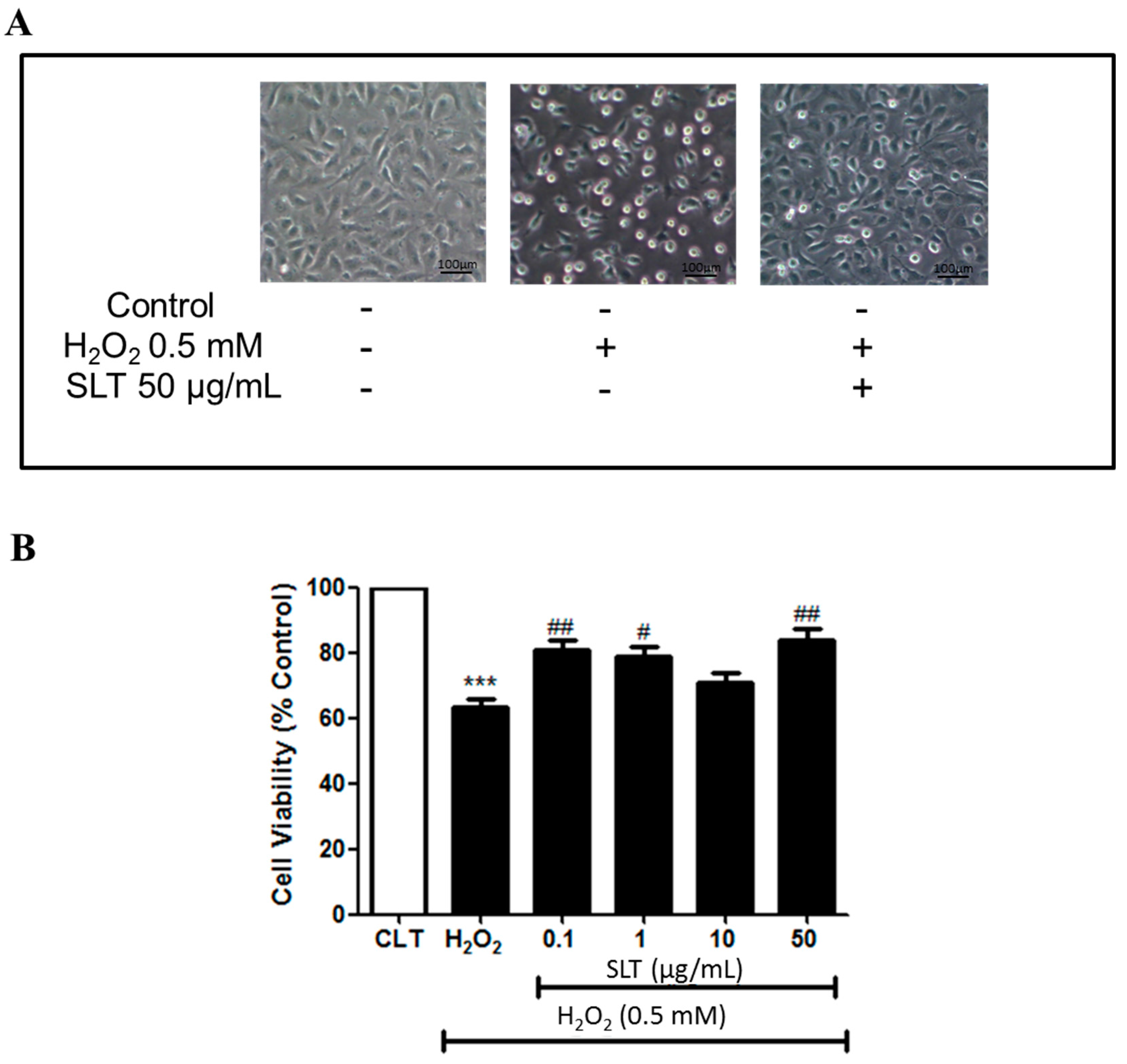
2.1. Effects of SLT on the Viability of EA.hy926 Cells Injured by H2O2
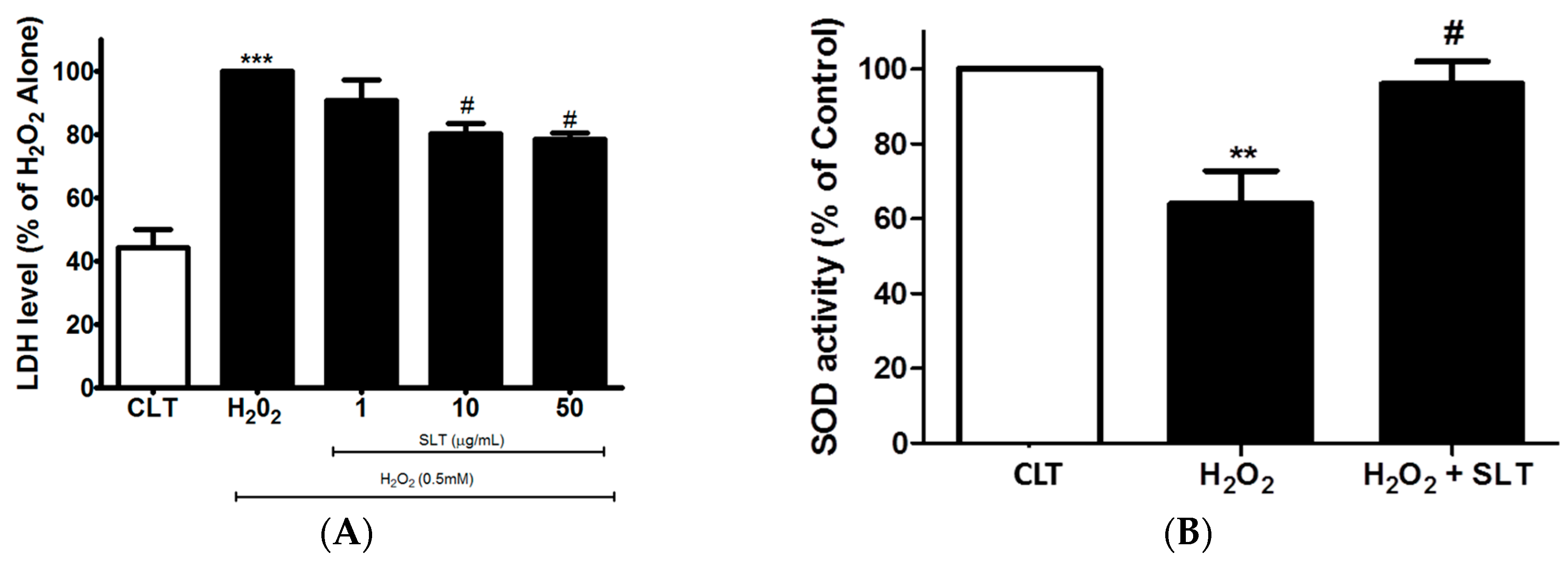
2.2. Effects of SLT on LDH Leakage and SOD Activity in H2O2 Treated EA.hy926 Cells
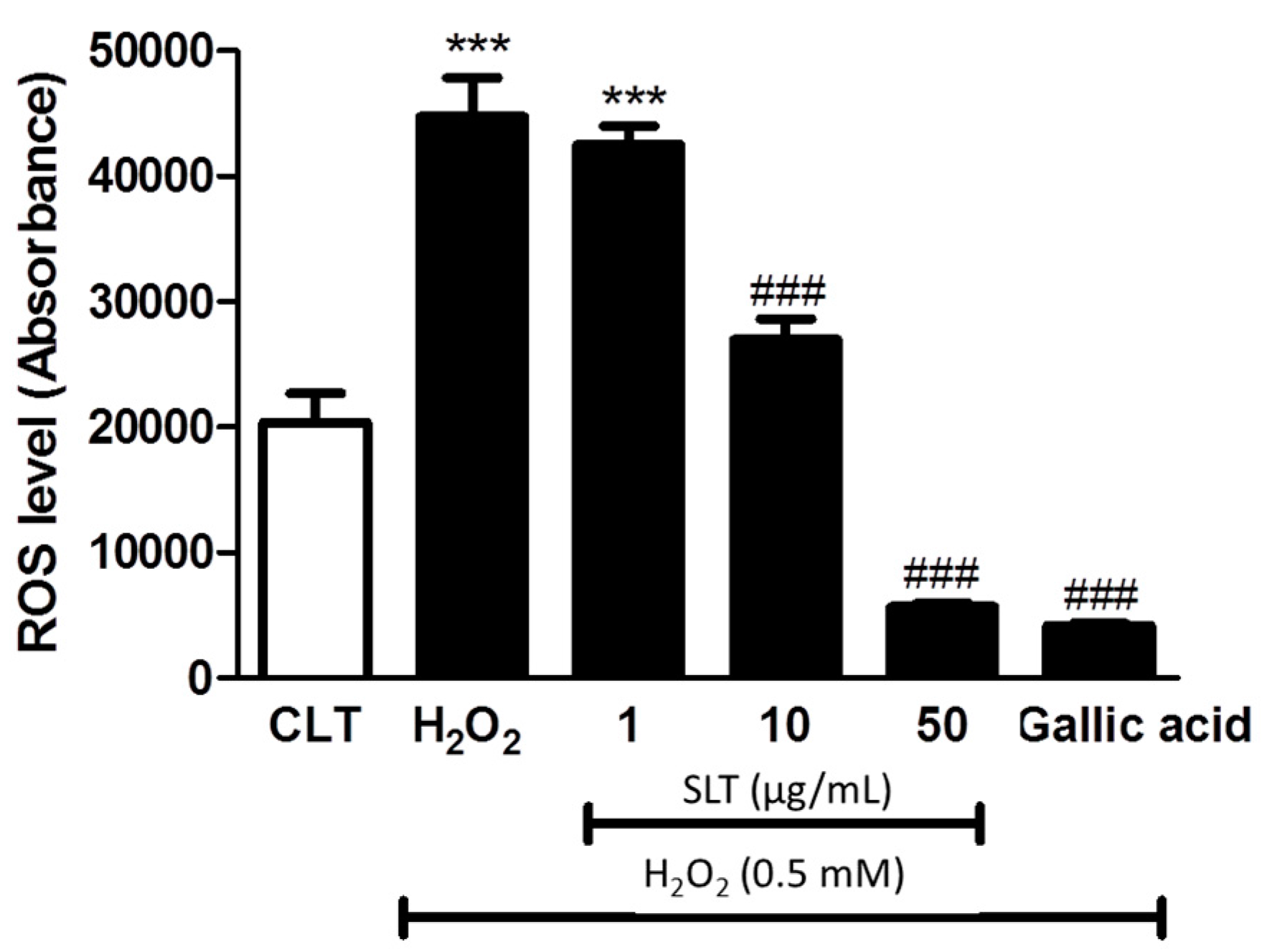
2.3. Effect of SLT on the Intracellular ROS Generation in H2O2 Treated EA.hy926 Cells
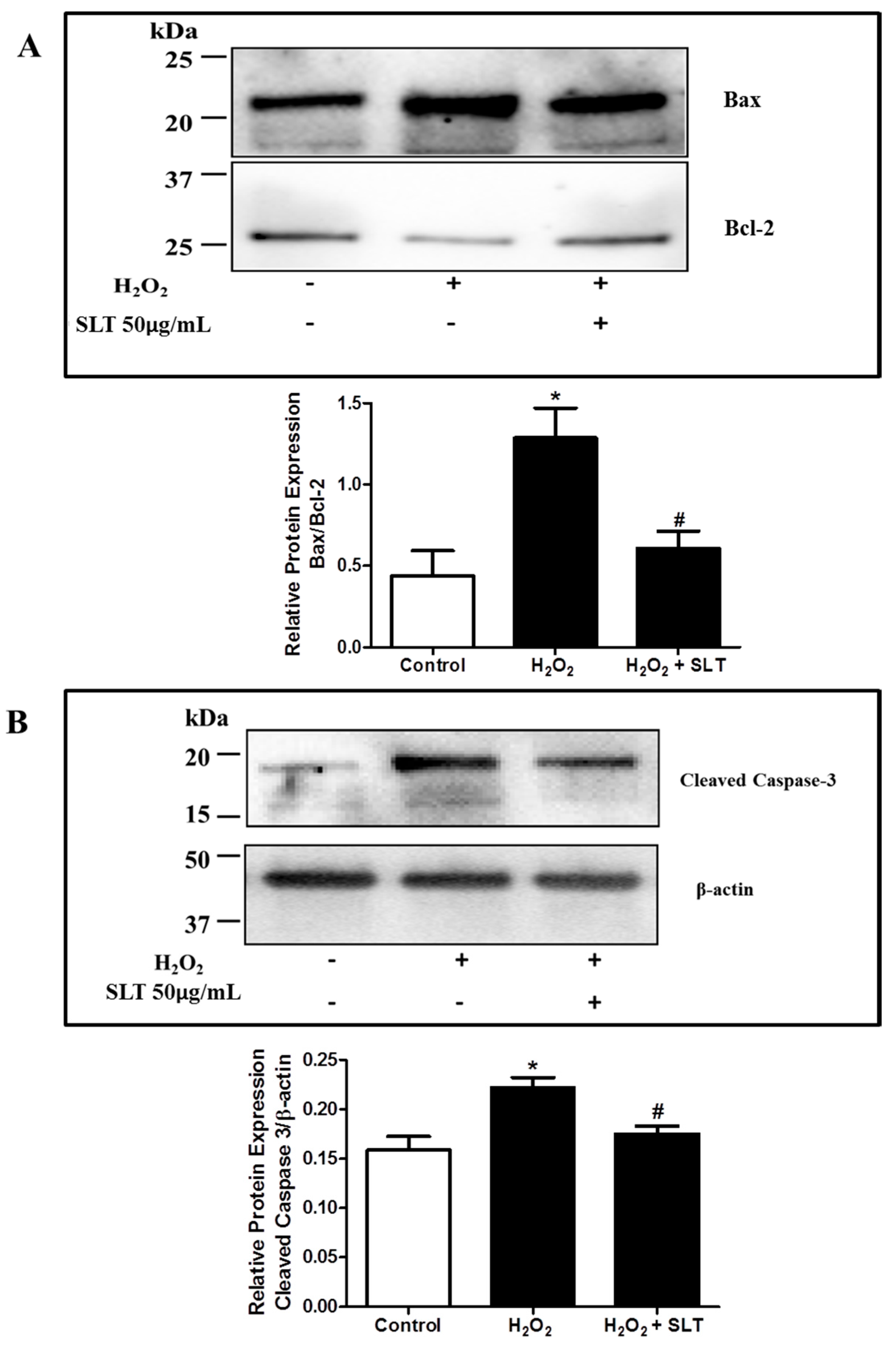
2.4. Effect of SLT on Protein Expression Level of Bax, Bcl-2, and Cleaved Caspase-3 in H2O2 Treated EA.hy926 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. EA.hy 926 Cell Culture
4.3. Measurement of Cell Viability
4.4. Measurement of Intracellular ROS Level
4.5. Measurement of Lactate Dehydrogenase (LDH) Leakage
4.6. Measurement of Intracellular SOD Activity
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Capone, C.; Faraco, G.; Anrather, J.; Zhou, P.; Iadecola, C. Cyclooxygenase 1-derived prostaglandin E2 and EP1 receptors are required for the cerebrovascular dysfunction induced by angiotensin II. Hypertension 2010, 55, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Kwan, Y.W.; Ngai, S.M. Modulatory effect of interleukin-1β on rat isolated basilar artery contraction. Eur. J. Pharmacol. 2006, 531, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; Srivastava, S.; Siow, R.C.; Cash, D.; Modo, M.; Duchen, M.R.; Fraser, P.A.; Williams, S.C.; Mann, G.E. Sulforaphane preconditioning of the Nrf2/HO-1 defense pathway protects the cerebral vasculature against blood-brain barrier disruption and neurological deficits in stroke. Free Radic. Biol. Med. 2013, 65, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Chrissobolis, S.; Drummond, G.R.; Faraci, F.M.; Sobey, C.G. Chronic aldosterone administration causes Nox2-mediated increases in reactive oxygen species production and endothelial dysfunction in the cerebral circulation. J. Hypertens. 2014, 32, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Kaneko, Y.; Tsukamoto, E.; Frankowski, H.; Kouyama, K.; Kita, Y.; Niikura, T.; Aiso, S.; Bredesen, D.E.; Matsuoka, M.; et al. Molecular characterization of neurohybrid cell death induced by Alzheimer’s amyloid-β peptides via p75NTR/PLAIDD. J. Neurochem. 2004, 90, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Poon, C.C.; Seto, S.W.; Au, A.L.; Zhang, Q.; Li, R.W.; Lee, W.Y.; Leung, G.P.; Kong, S.K.; Yeung, J.H.; Ngai, S.M.; et al. Mitochondrial monoamine oxidase-A-mediated hydrogen peroxide generation enhances 5-hydroxytryptamine-induced contraction of rat basilar artery. Br. J. Pharmacol. 2010, 161, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.L.; Hou, Y.C.; Chao, P.D.; Weng, C.S.; Ho, F.M. The antioxidant effects of quercetin metabolites on the prevention of high glucose-induced apoptosis of human umbilical vein endothelial cells. Br. J. Nutr. 2009, 101, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Using zebrafish to assess the impact of drugs on neural development and function. Expert Opin. Drug Discov. 2009, 4, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Chang, D.; Jenkins, A.; Bensoussan, A.; Kiat, H. Angiogenesis in Ischemic Stroke and Angiogenic Effects of Chinese Herbal Medicine. J. Clin. Med. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Rodriguez-Yanez, M.; Sobrino, T.; Leira, R.; Castillo, J. Platelets, inflammation, and atherothrombotic neurovascular disease: The role of endothelial dysfunction. Cerebrovasc. Dis. 2005, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Santulli, G.; del Giudice, C.; Anastasio, A.; Trimarco, B.; Iaccarino, G. Endothelial cells are able to synthesize and release catecholamines both in vitro and in vivo. Hypertension 2012, 60, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G. MicroRNAs and Endothelial (Dys) Function. J. Cell. Physiol. 2016, 231, 1638–1644. [Google Scholar] [CrossRef] [PubMed]
- Quesada, I.M.; Lucero, A.; Amaya, C.; Meijles, D.N.; Cifuentes, M.E.; Pagano, P.J.; Castro, C. Selective inactivation of NADPH oxidase 2 causes regression of vascularization and the size and stability of atherosclerotic plaques. Atherosclerosis 2015, 242, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fotheringham, L.; Wittchen, E.S.; Hartnett, M.E. Rap1 GTPase Inhibits Tumor Necrosis Factor-alpha-Induced Choroidal Endothelial Migration via NADPH Oxidase- and NF-κB-Dependent Activation of Rac1. Am. J. Pathol. 2015, 185, 3316–3325. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.C.; Gao, X.F.; Ruan, C.; Ge, Z.R.; Lu, J.D.; Zhang, J.J.; Zhang, Y.; Wang, L.; Shi, H.M. miR-103 Regulates Oxidative Stress by Targeting the BCL2/Adenovirus E1B 19 kDa Interacting Protein 3 in HUVECs. Oxidative Med. Cell. Longev. 2015, 2015, 489647. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Wong, C.Y.; Wan, C.W.; Kwok, C.Y.; Wu, J.H.; Ng, K.M.; So, C.H.; Au, A.L.; Poon, C.C.; Seto, S.W.; et al. Evaluation of anti-oxidant capacity of root of Scutellaria baicalensis Georgi, in comparison with roots of Polygonum multiflorum Thunb and Panax ginseng CA Meyer. Am. J. Chin. Med. 2010, 38, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, F.; Wei, C.; Song, H.; Wu, L.; Tang, Y.; Jia, J. Rationale and design of a multicenter, phase 2 clinical trial to investigate the efficacy of traditional Chinese medicine SaiLuoTong in vascular dementia. J. Stroke Cerebrovasc. Dis. 2014, 23, 2626–2634. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.Z.; Yeung, A.; Liu, J.X.; Camfield, D.A.; Blasio, F.M.; Pipingas, A.; Scholey, A.B.; Stough, C.; Chang, D.H. The effect of Sailuotong (SLT) on neurocognitive and cardiovascular function in healthy adults: A randomised, double-blind, placebo controlled crossover pilot trial. BMC Complement. Altern. Med. 2016, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.L.; Cong, W.; Wang, J.; Zhang, Y. SLT for Vascular Dementia; China Academy of Chinese Medical Sciences: Beijing, China, 2008. [Google Scholar]
- Xu, L.; Liu, J.X.; Cong, W.H.; Wei, C.E. Effects of Weinaokang capsule on intracephalic cholinergic system and capability of scavenging free radicas in chronic cerebral hypoperfusion rats. Zhongguo Zhong Yao Za Zhi 2008, 33, 531–534. [Google Scholar] [PubMed]
- Liu, J. Development of an Evidence-Based Chinese Herbal Medicine for the Management of Vascular Dementia. Ph.D. Thesis, The University of Western Sydney, Sydney, Australia, 2008. [Google Scholar]
- Zheng, Y.Q.; Liu, J.X.; Wang, J.N.; Xu, L. Effects of crocin on reperfusion-induced oxidative/nitrative injury to cerebral microvessels after global cerebral ischemia. Brain Res. 2007, 1138, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zheng, L.; Lu, S.; Yang, Y. Neuroprotective effects of pretreatment of ginsenoside Rb1 on severe cerebral ischemia-induced injuries in aged mice: Involvement of anti-oxidant signaling. Geriatr. Gerontol. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Nie, B.; Fu, S.; Hu, J.; Yin, L.; Lin, L.; Wang, X.; Lu, P.; Xu, X.M. EGb761 protects hydrogen peroxide-induced death of spinal cord neurons through inhibition of intracellular ROS production and modulation of apoptotic regulating genes. J. Mol. Neurosci. 2009, 38, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.C.; Du, G.H.; Zhang, J.T. Protective effect of ginkgo biloba extract on endothelial cell against damage induced by oxidative stress. J. Cardiovasc. Pharmacol. 2002, 40, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Sung, J.H.; Cho, E.H.; Won, C.K.; Lee, H.J.; Kim, M.O.; Koh, P.O. Gingko biloba Extract (EGb 761) prevents ischemic brain injury by activation of the Akt signaling pathway. Am. J. Chin. Med. 2009, 37, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Q.; Liu, J.X.; Li, X.Z.; Xu, L. Effects and mechanism of Weinaokang on reperfusion-induced vascular injury to cerebral microvessels after global cerebral ischemia. Chin. J. Integr. Med. 2010, 16, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Chang, D.; Ko, W.M.; Kiat, H.; Bensoussan, A.; Lee, S.M.Y.; Hoi, M.P.M.; Liu, J. Angiogenic effects of Sailuotong (SLT) on EA.hy926 cell in vitro and zebrafish in vivo. Int. J. Mol. Sci. 2017. manuscript under preparation. [Google Scholar]
- Chen, C.H.; Liu, T.Z.; Chen, C.H.; Wong, C.H.; Chen, C.H.; Lu, F.J.; Chen, S.C. The efficacy of protective effects of tannic acid, gallic acid, ellagic acid, and propyl gallate against hydrogen peroxide-induced oxidative stress and DNA damages in IMR-90 cells. Mol. Nutr. Food Res. 2007, 51, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Yang, G.Y.; Kiat, H.; Bensoussan, A.; Kwan, Y.W.; Chang, D. Diabetes Mellitus, Cognitive Impairment, and Traditional Chinese Medicine. Int. J. Endocrinol. 2015, 2015, 810439. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, E.A.; Mahmoud, A.M.; Khalifa, A.M.; Ali, S.S. Physiological and pathophysiological reactive oxygen species as probed by EPR spectroscopy: The underutilized research window on muscle ageing. J. Physiol. 2016, 594, 4591–4613. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U. Nitric oxide and oxidative stress in vascular disease. Pflugers Arch. 2010, 459, 923–939. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J. Physiol. Pharmacol. 2009, 60, 111–116. [Google Scholar] [PubMed]
- Hermann, C.; Zeiher, A.M.; Dimmeler, S. Shear stress inhibits H2O2-induced apoptosis of human endothelial cells by modulation of the glutathione redox cycle and nitric oxide synthase. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 3588–3592. [Google Scholar] [CrossRef] [PubMed]
- Aliev, G.; Palacios, H.H.; Walrafen, B.; Lipsitt, A.E.; Obrenovich, M.E.; Morales, L. Brain mitochondria as a primary target in the development of treatment strategies for Alzheimer disease. Int. J. Biochem. Cell Biol. 2009, 41, 1989–2004. [Google Scholar] [CrossRef] [PubMed]
- Breton-Romero, R.; Lamas, S. Hydrogen peroxide signaling in vascular endothelial cells. Redox Biol. 2014, 2, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Asahina, T.; Kashiwagi, A.; Nishio, Y.; Ikebuchi, M.; Harada, N.; Tanaka, Y.; Takagi, Y.; Saeki, Y.; Kikkawa, R.; Shigeta, Y. Impaired activation of glucose oxidation and NADPH supply in human endothelial cells exposed to H2O2 in high-glucose medium. Diabetes 1995, 44, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Belkhiri, A.; Richards, C.; Whaley, M.; McQueen, S.A.; Orr, F.W. Increased expression of activated matrix metalloproteinase-2 by human endothelial cells after sublethal H2O2 exposure. Lab. Investig. 1997, 77, 533–539. [Google Scholar] [PubMed]
- Jia, L.Q.; Yang, G.L.; Ren, L.; Chen, W.N.; Feng, J.Y.; Cao, Y.; Zhang, L.; Li, X.T.; Lei, P. Tanshinone IIA reduces apoptosis induced by hydrogen peroxide in the human endothelium-derived EA.hy926 cells. J. Ethnopharmacol. 2012, 143, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Jiang, F.; Wang, B.; Yu, Y.; Zhang, X.; Yin, Z.; Liu, C. Ophiopogonin D prevents H2O2-induced injury in primary human umbilical vein endothelial cells. J. Ethnopharmacol. 2010, 128, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Giray, B.; Kan, E.; Bali, M.; Hincal, F.; Basaran, N. The effect of vitamin E supplementation on antioxidant enzyme activities and lipid peroxidation levels in hemodialysis patients. Clin. Chim. Acta 2003, 338, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.L.; Kang, Y.; Li, J.K.; Li, Q.S.; Liu, E.L.; Liu, X.X. Protective effects of hyperoside against H2O2-induced apoptosis in human umbilical vein endothelial cells. Mol. Med. Rep. 2016, 14, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Buccellato, L.J.; Tso, M.; Akinci, O.I.; Chandel, N.S.; Budinger, G.R. Reactive oxygen species are required for hyperoxia-induced Bax activation and cell death in alveolar epithelial cells. J. Biol. Chem. 2004, 279, 6753–6760. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Afaq, F.; Mukhtar, H. Involvement of nuclear factor-κB, Bax and Bcl-2 in induction of cell cycle arrest and apoptosis by apigenin in human prostate carcinoma cells. Oncogene 2002, 21, 3727–3738. [Google Scholar] [CrossRef] [PubMed]
- Vogel, M.W. Cell death, Bcl-2, Bax, and the cerebellum. Cerebellum 2002, 1, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kang, J.S.; Park, J.H.; Lee, Y.J.; Choi, J.S.; Kang, Y.H. Polyphenolic flavonoids differ in their antiapoptotic efficacy in hydrogen peroxide-treated human vascular endothelial cells. J. Nutr. 2003, 133, 985–991. [Google Scholar] [PubMed]
- Fuentes-Prior, P.; Salvesen, G.S. The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem. J. 2004, 384, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Acharya, N.K.; Goldwaser, E.L.; Forsberg, M.M.; Godsey, G.A.; Johnson, C.A.; Sarkar, A.; DeMarshall, C.; Kosciuk, M.C.; Dash, J.M.; Hale, C.P.; et al. Sevoflurane and Isoflurane induce structural changes in brain vascular endothelial cells and increase blood-brain barrier permeability: Possible link to postoperative delirium and cognitive decline. Brain Res. 2015, 1620, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.; Grant, M.M.; Aldred, S. Oxidative stress in vascular dementia and Alzheimer’s disease: A common pathology. J. Alzheimers Dis. 2009, 17, 245–257. [Google Scholar] [PubMed]
- Baskys, A.; Hou, A.C. Vascular dementia: Pharmacological treatment approaches and perspectives. Clin. Interv. Aging 2007, 2, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Kalaria, R.N.; Corbett, A.; Ballard, C. Update on Vascular Dementia. J. Geriatr. Psychiatry Neurol. 2016, 29, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Cong, W.H.; Liu, J.X.; Xu, L. Effects of extracts of Ginseng and Ginkgo biloba on hippocampal acetylcholine and monoamines in PDAP-pV717I transgenic mice. Zhongguo Zhong Xi Yi Jie He Za Zhi 2007, 27, 810–813. [Google Scholar] [PubMed]
- Coelho-Santos, V.; Socodato, R.; Portugal, C.; Leitao, R.A.; Rito, M.; Barbosa, M.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Minshall, R.D.; et al. Methylphenidate-triggered ROS generation promotes caveolae-mediated transcytosis via Rac1 signaling and c-Src-dependent caveolin-1 phosphorylation in human brain endothelial cells. Cell. Mol. Life Sci. 2016, 73, 4701–4716. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Lopez, I.; Diaz-Morales, N.; Rovira-Llopis, S.; de Maranon, A.M.; Orden, S.; Alvarez, A.; Banuls, C.; Rocha, M.; Murphy, M.P.; Hernandez-Mijares, A.; et al. The mitochondria-targeted antioxidant MitoQ modulates oxidative stress, inflammation and leukocyte-endothelium interactions in leukocytes isolated from type 2 diabetic patients. Redox Biol. 2016, 10, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Santulli, G.; Reiken, S.R.; Yuan, Q.; Osborne, B.W.; Chen, B.X.; Marks, A.R. Mitochondrial oxidative stress promotes atrial fibrillation. Sci. Rep. 2015, 5, 11427. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar] [PubMed]
- Liu, B.; Ren, K.D.; Peng, J.J.; Li, T.; Luo, X.J.; Fan, C.; Yang, J.F.; Peng, J. Suppression of NADPH oxidase attenuates hypoxia-induced dysfunctions of endothelial progenitor cells. Biochem. Biophys. Res. Commun. 2016. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seto, S.W.; Chang, D.; Ko, W.M.; Zhou, X.; Kiat, H.; Bensoussan, A.; Lee, S.M.Y.; Hoi, M.P.M.; Steiner, G.Z.; Liu, J. Sailuotong Prevents Hydrogen Peroxide (H2O2)-Induced Injury in EA.hy926 Cells. Int. J. Mol. Sci. 2017, 18, 95. https://doi.org/10.3390/ijms18010095
Seto SW, Chang D, Ko WM, Zhou X, Kiat H, Bensoussan A, Lee SMY, Hoi MPM, Steiner GZ, Liu J. Sailuotong Prevents Hydrogen Peroxide (H2O2)-Induced Injury in EA.hy926 Cells. International Journal of Molecular Sciences. 2017; 18(1):95. https://doi.org/10.3390/ijms18010095
Chicago/Turabian StyleSeto, Sai Wang, Dennis Chang, Wai Man Ko, Xian Zhou, Hosen Kiat, Alan Bensoussan, Simon M. Y. Lee, Maggie P. M. Hoi, Genevieve Z. Steiner, and Jianxun Liu. 2017. "Sailuotong Prevents Hydrogen Peroxide (H2O2)-Induced Injury in EA.hy926 Cells" International Journal of Molecular Sciences 18, no. 1: 95. https://doi.org/10.3390/ijms18010095
APA StyleSeto, S. W., Chang, D., Ko, W. M., Zhou, X., Kiat, H., Bensoussan, A., Lee, S. M. Y., Hoi, M. P. M., Steiner, G. Z., & Liu, J. (2017). Sailuotong Prevents Hydrogen Peroxide (H2O2)-Induced Injury in EA.hy926 Cells. International Journal of Molecular Sciences, 18(1), 95. https://doi.org/10.3390/ijms18010095