A Novel Vav3 Homolog Identified in Lamprey, Lampetra japonica, with Roles in Lipopolysaccharide-Mediated Immune Response


Abstract
:1. Introduction
2. Results
2.1. Identification of Lamprey Vav3 Sequence
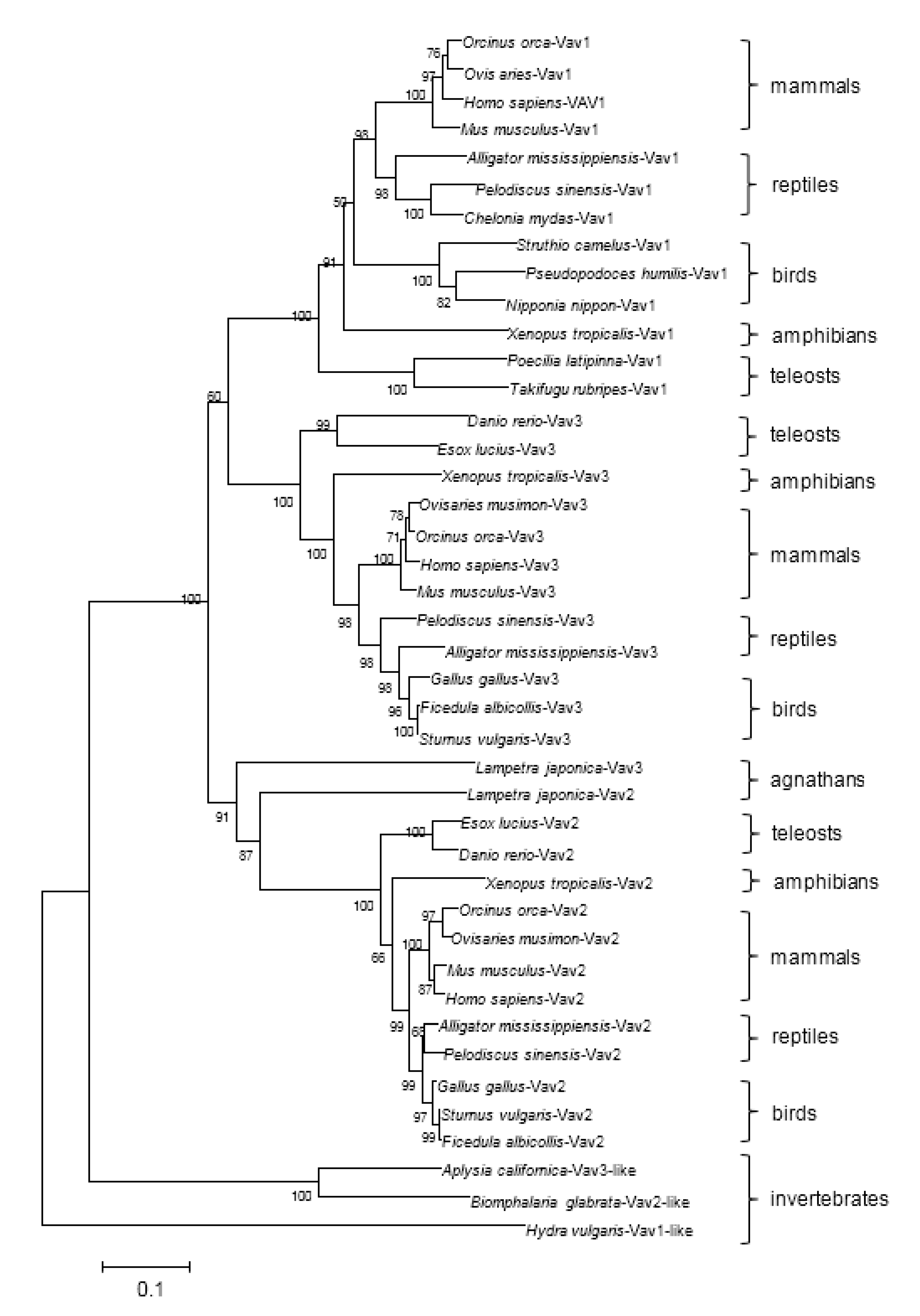
2.2. Phylogenetic Analysis of the Vav Family
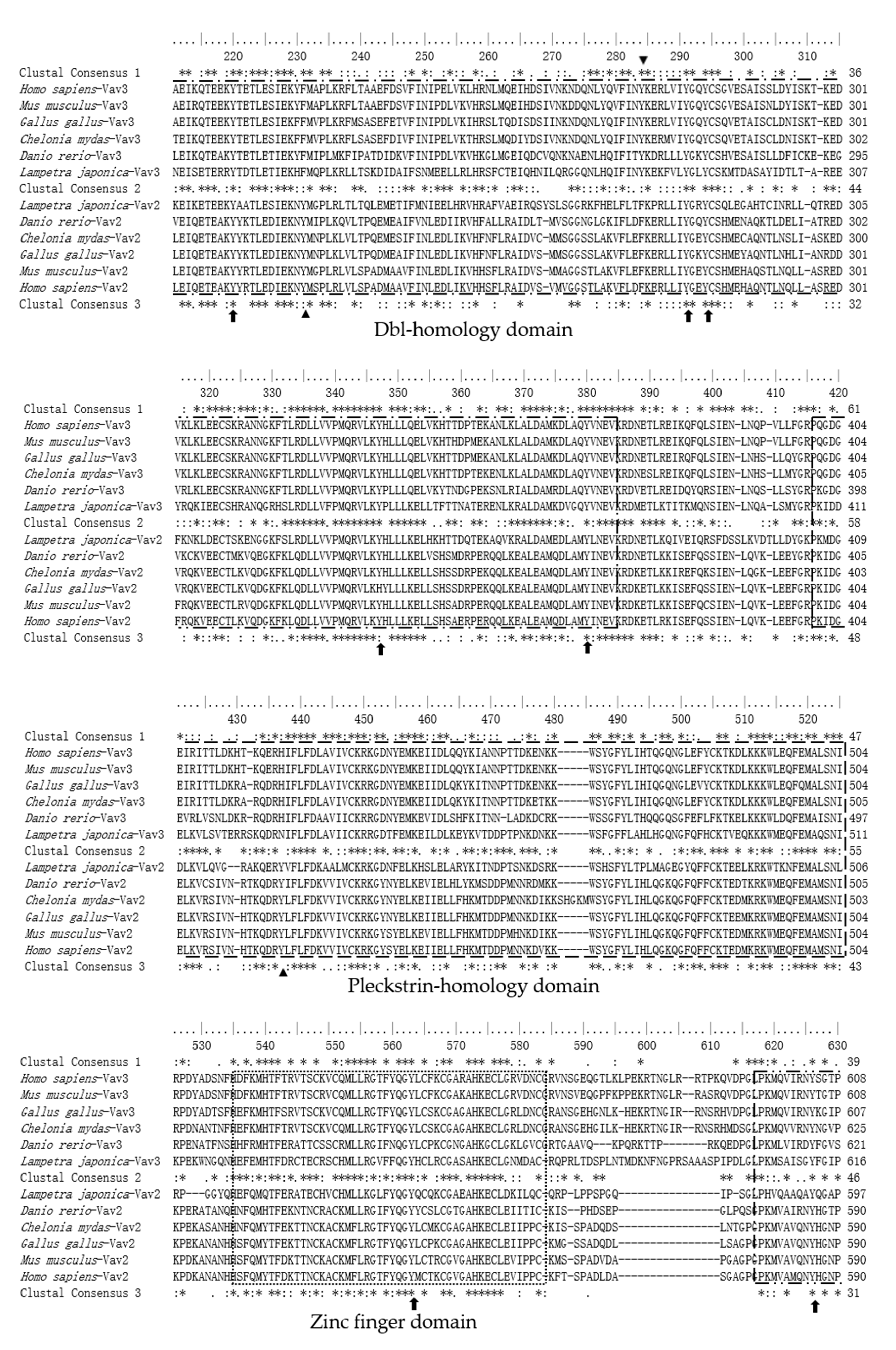
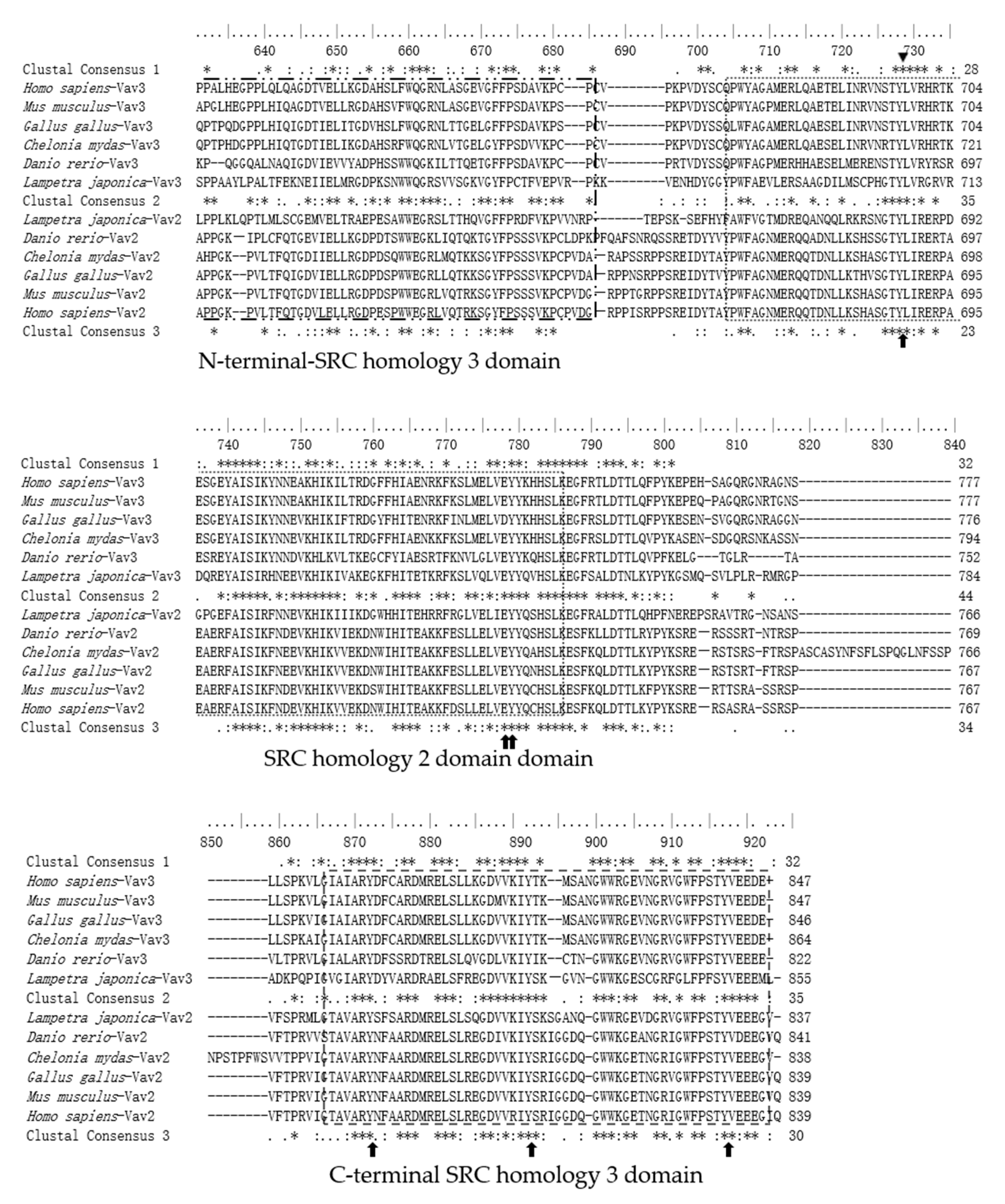
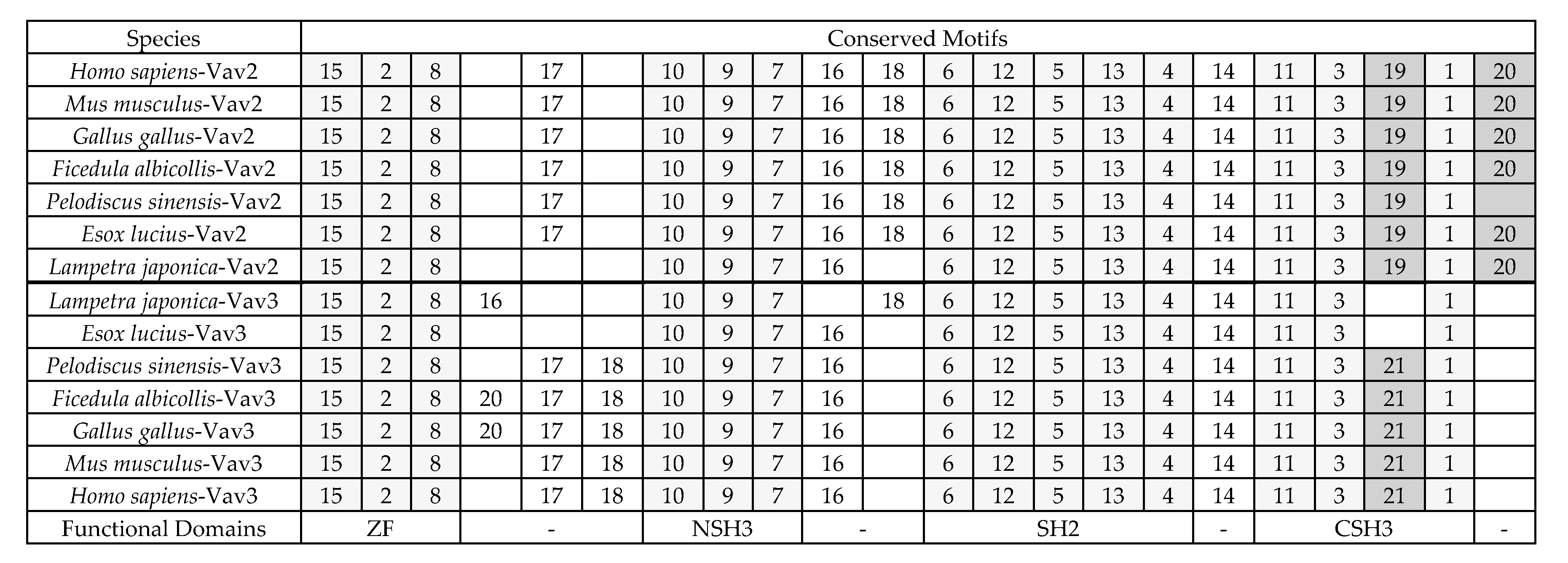
2.3. The Sequence Differences between Vav3 and Vav2 Subfamily
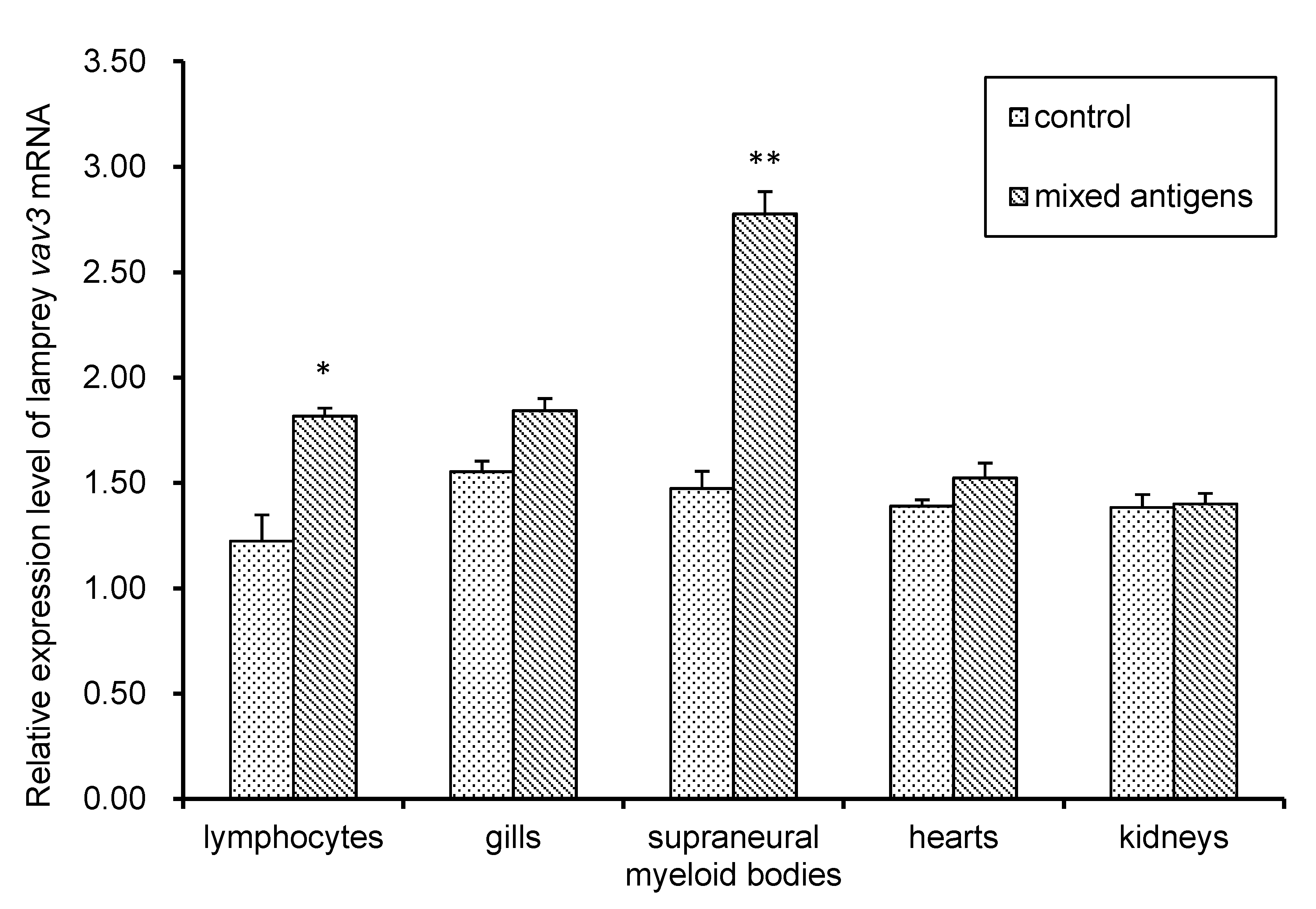
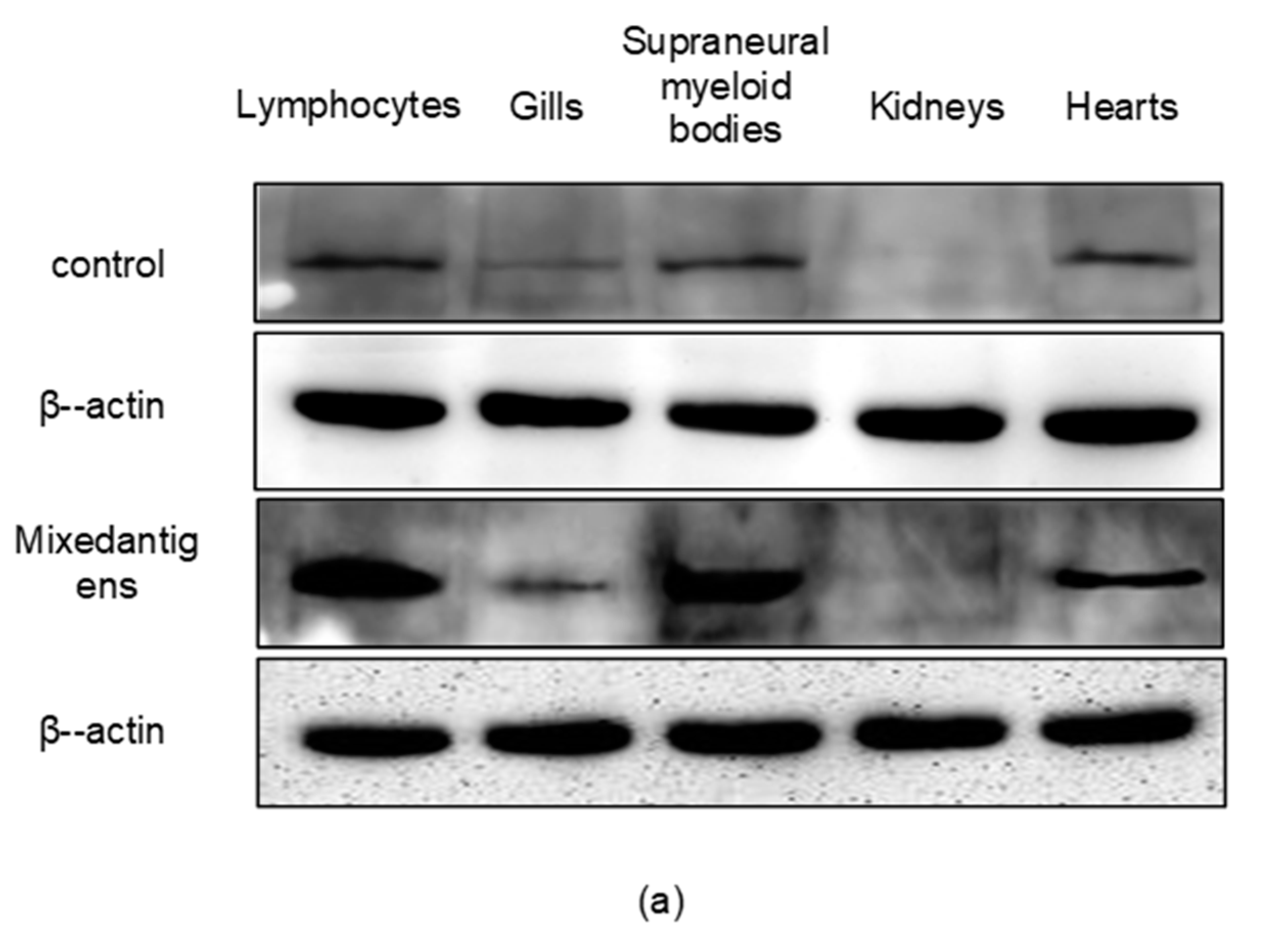
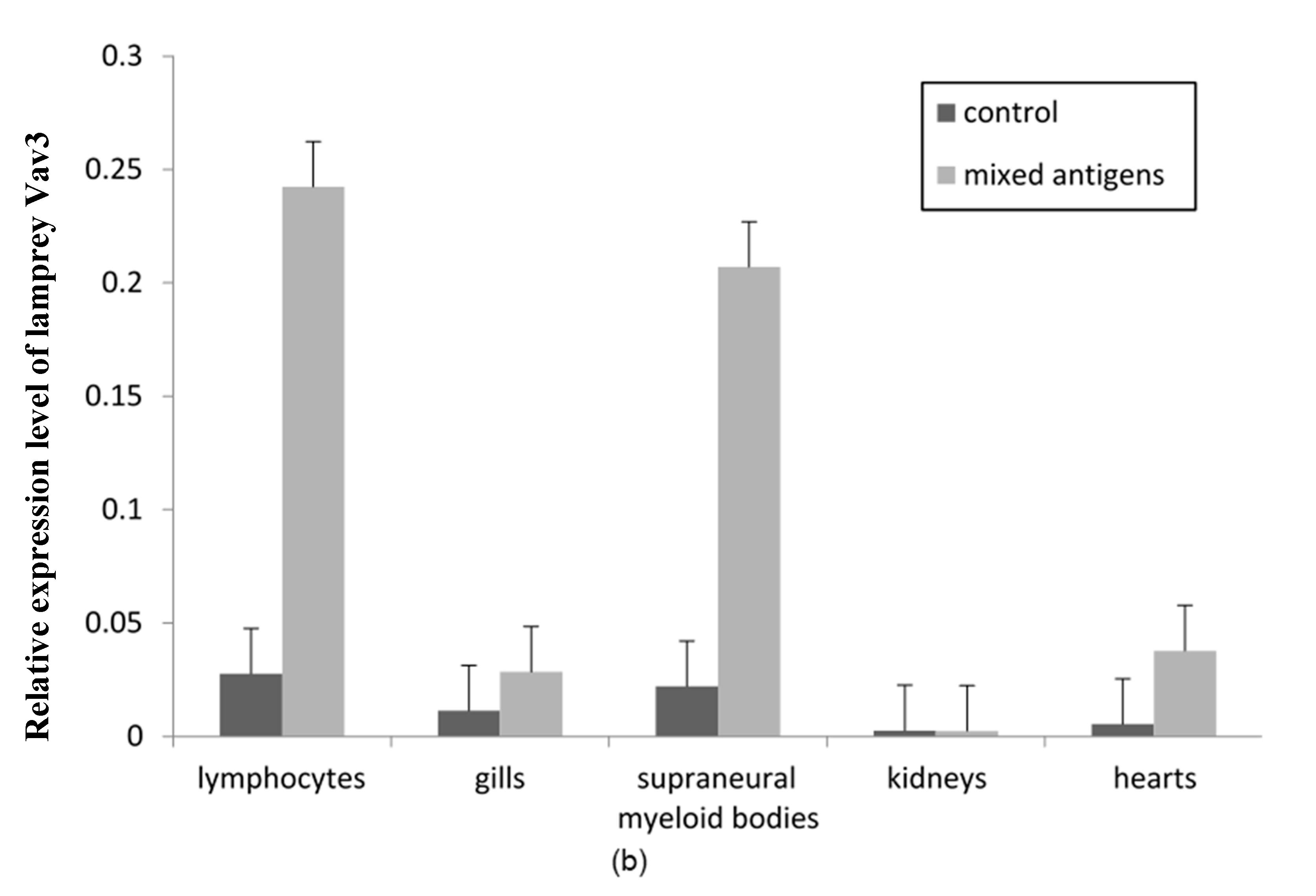
2.4. The Expression Pattern of Lamprey Vav3 mRNA and Protein after Antigen Stimulation
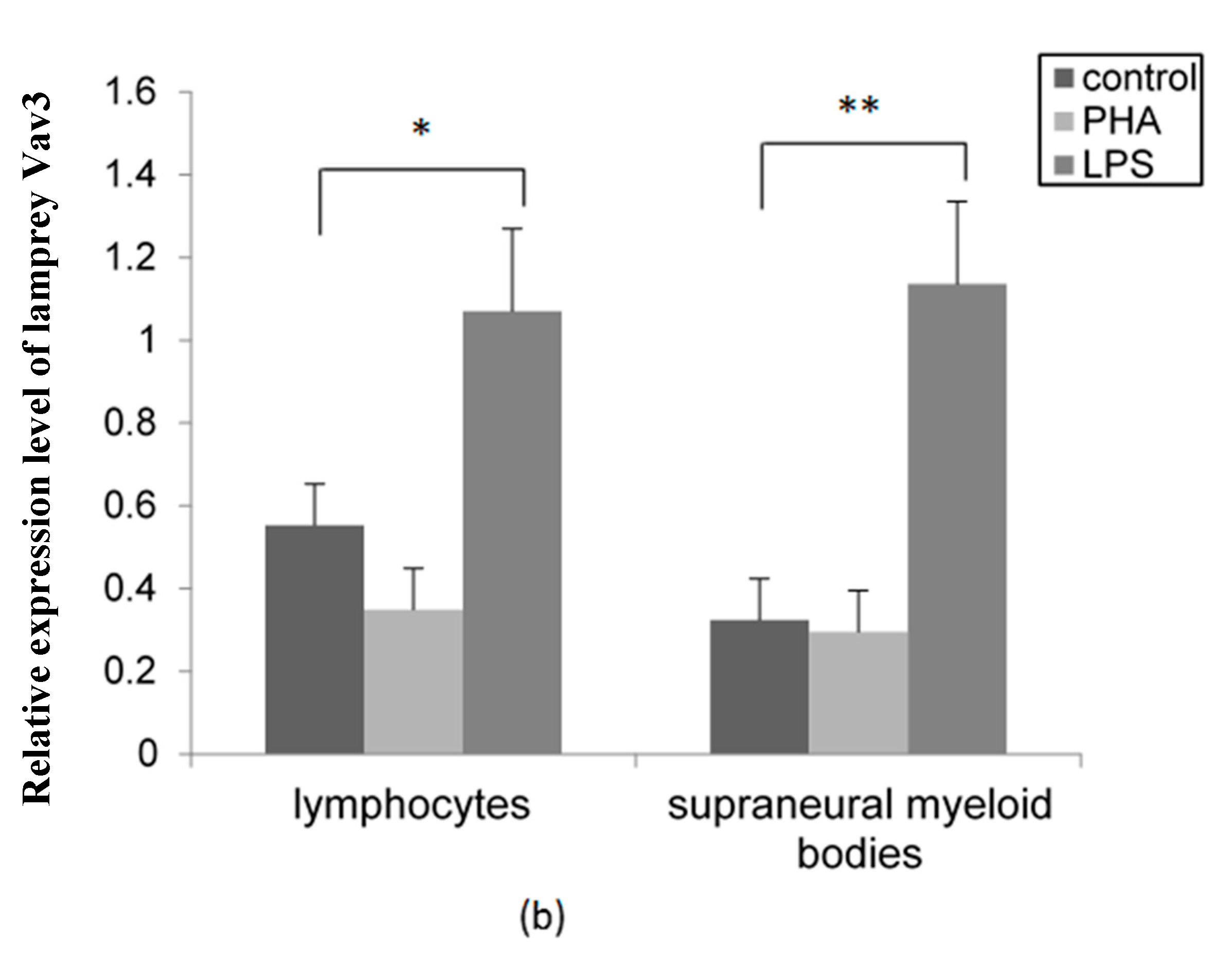
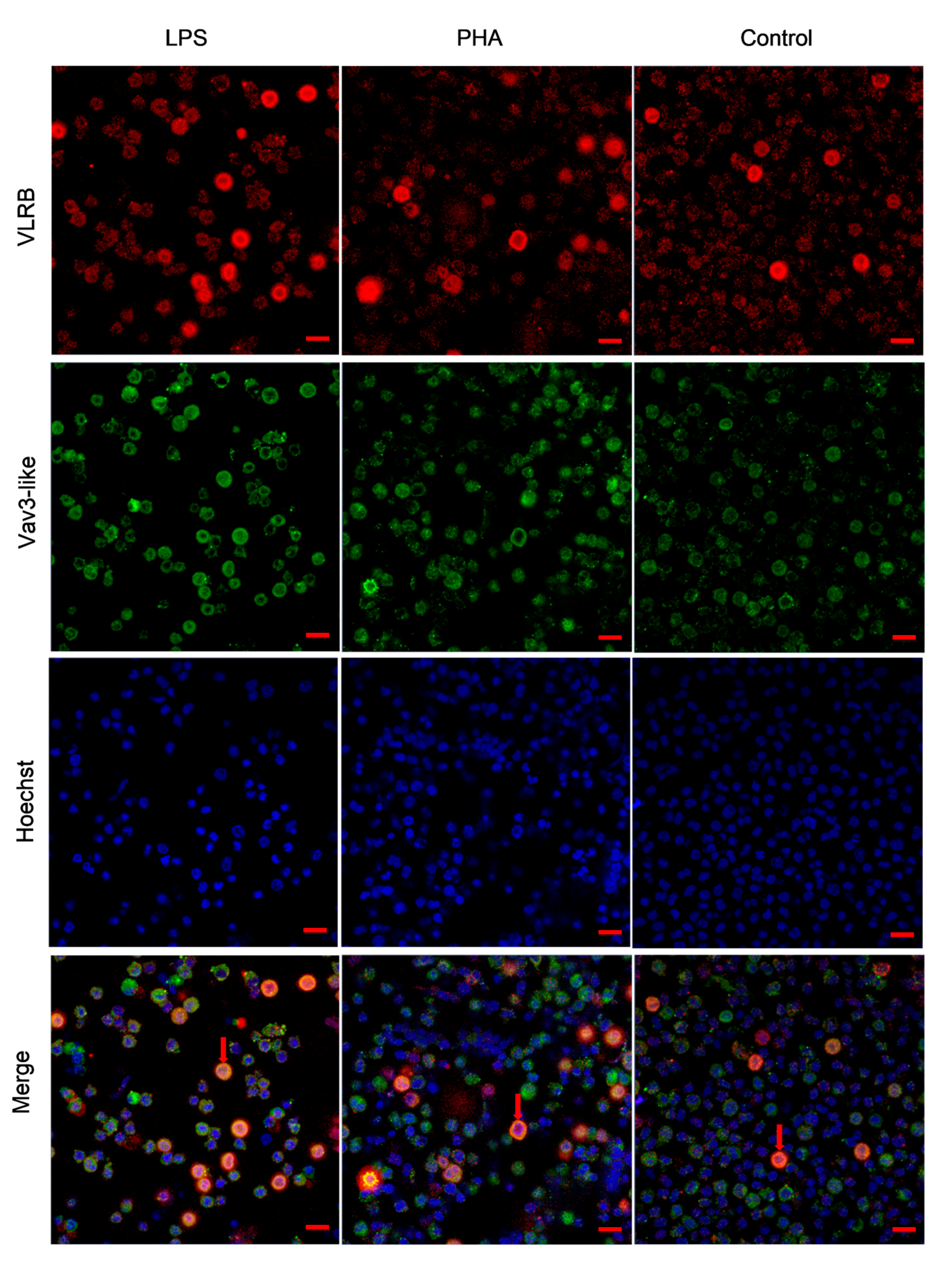
2.5. The Distribution Pattern of Lamprey Vav3 in VLRB+ Lymphocytes after Stimulation with Lipopolysaccharide (LPS) and Phytohemagglutinin (PHA)
3. Discussion
4. Materials and Methods
4.1. Handing of Animals
4.2. Amplification of the Lamprey vav3 cDNA Fragment
4.3. Real-Time Quantitative PCR
4.4. Analyses of Sequence Similarity and Conserved Motifs and Phylogenetic Tree Reconstruction
4.5. The Expression and Purification of Rlamprey Vav3
4.6. Mass Spectrometry of Recombinant Lamprey Vav3 Protein
4.7. Production of Polyclonal Antibody
4.8. Western Blotting Analysis
4.9. Immunofluorescence Assay
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Vav | Vav guanine nucleotide exchange factor |
| TCR | T-cell receptor |
| BCR | B-cell receptor |
| LPS | Lipopolysaccharide |
| PHA | Phytohemagglutinin |
| SH | Src-homology |
| VLR | Variable lymphocyte receptor |
| ORF | Open reading frame |
| NJ | Neighbor-Jointing |
| MEME | Multiple Em for Motif Elicitation |
| Q-PCR | The real-time quantitative PCR |
| pAb | Polyclonal antibody |
| NFAT | The nuclear factor of activated T-cells |
| SMART | Simple Modular Architecture Research Tool |
| ELISA | Enzyme-linked immunosorbent assay |
| PVDF | Polyvinylidene fluoride |
| IPTG | Isopropyl β-d-1-thiogalactopyranoside |
Appendix A. The Expression and Purification of the Recombinant Lamprey Vav3 Protein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences |
|---|---|
| Primers designed for Lja-Vav3 open reading frame (ORF) cloning | |
| Forward-Lja-vav3 | 5′-ATGGAGGAAGAGGGCAGGTT-3′ |
| Reverse-Lja-vav3 | 5′-CGTGGAAGAAGAAATGCTCTGA-3′ |
| Primers designed for real-time PCR | |
| Lja-vav3 (upstream) | 5′-ACCTGCGTCAACAGATTCGG-3′ |
| Lja-vav3 (downstream) | 5′-CACCGATGCCTTTTTTCTGC-3′ |
| gapdh (upstream) | 5′-ACCAACTGCCTGGCTCCT-3′ |
| gapdh(downstream) | 5′-TCTTCTGCGTTGCCGTGT-3′ |
| Primers for subcloning Lja-vav3 ORF into the pET-32a vector | |
| Upstream | 5′-ATGCCTGATATCGGATCCGAATTCATGGAGGAAGAGGGCAGGTTGTGGC-3′ |
| Downstream | 5′-GTGCTCGAGTGCGGCCGCAAGCTTGAGCATTTCTTCTTCCACGTAGGAG-3′ |

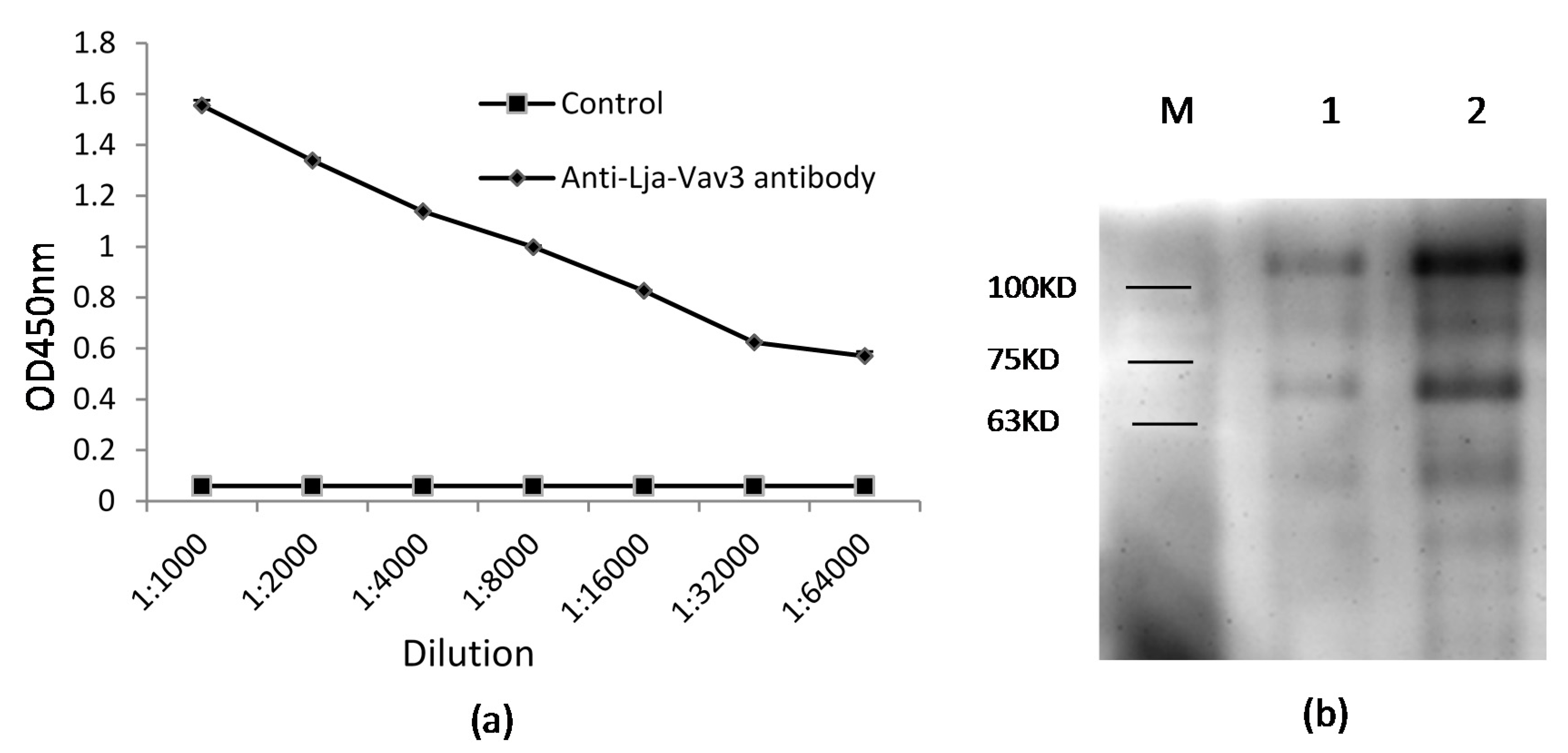
Appendix B. The Production and Verification of Polyclonal Antibody

References
- Tybulewicz, V.L. Vav-family proteins in T-cell signaling. Curr. Opin. Immunol. 2005, 17, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R. Vav proteins, adaptors and cell signaling. Oncogene 2001, 20, 6372–6381. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.; Billaideau, D.D. VAV proteins as signal integrators for multi-subunit immune-recognition receptors. Nat. Rev. Immunol. 2002, 2, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Movilla, N.; Busteo, X.R. Biological and regulatory properties of Vav-3, a new member of the Vav family of oncoproteins. Mol. Cell Biol. 1999, 19, 7870–7885. [Google Scholar] [CrossRef] [PubMed]
- Chrencik, E.; Brooun, A.; Zhang, H.; Mathews, H.; Hura, G.L.; Foster, S.A.; Perry, J.J.; Streiff, M.; Ramage, P.; Widmer, H.; et al. Structural basis of guanine nucleotide exchange mediated by the T-cell essential Vav1. J. Mol. Biol. 2008, 380, 828–843. [Google Scholar] [CrossRef] [PubMed]
- Rapley, J.; Tybulewicz, V.L.; Rittinger, K. Crucial structural role for the PH and C1 domains of the Vav1 exchange factor. EMBO Rep. 2008, 9, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Martins, I.R.; Li, P.; Amarasinghe, G.K.; Umetani, J.; Fernandez-Zapico, M.E.; Billadeau, D.D.; Machius, M.; Tomchick, D.R.; Rosen, M.K. Structural and energetic mechanisms of cooperative autoinhibition and activation of Vav1. Cell 2010, 140, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Nagata, K.; Hachimori, Y.; Horiuchi, M.; Ogura, K.; Mandiyan, V.; Schlessinger, J.; Inagaki, F. Novel recognition mode between Vav and Grb2 SH3 domains. EMBO J. 2001, 20, 2995–3007. [Google Scholar] [CrossRef] [PubMed]
- Ogura, K.; Nagata, K.; Horiuchi, M.; Ebisui, E.; Hasuda, T.; Yuzawa, S.; Nishida, M.; Hatanaka, H.; Inagaki, F. Solution structure of N-terminal SH3 domain of Vav and the recognition site for Grb2 C-terminal SH3 domain. J. Biomol. 2002, 22, 37–46. [Google Scholar] [CrossRef]
- Moores, S.L.; Selfors, L.M.; Fredericks, J.; Breit, T.; Fujikawa, K.; Alt, F.W.; Brugge, J.S.; Swat, W. Vav family proteins couple to diverse cell surface receptors. Mol. Cell Biol. 2000, 20, 6364–6373. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R. Regulatory and signaling properties of the Vav family. Mol. Cell Biol. 2000, 20, 1461–1477. [Google Scholar] [CrossRef] [PubMed]
- Citterio, C.; Menacho, -M.M.; Garcia, -E.R.; Larive, R.M.; Barreiro, O.; Sanchez, -M.F.; Paramio, J.M.; Bustelo, X.R. The Rho exchange factors Vav2 and Vav3 control a lung metastasis-specific transcriptional program in breast cancer cells. Sci. Sig. 2012, 5, ra71. [Google Scholar] [CrossRef] [PubMed]
- Inabe, K.; Ishiai, M.; Scharenberg, A.M.; Freshney, N.; Downward, J.; Kurosaki, T. Vav3 Modulates B cell receptor responses by regulating phosphoinositide 3-kinase activation. J. Exp. Med. 2002, 195, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, K.; Miletic, A.V.; Alt, F.W.; Faccio, R.; Brown, T.; Hoog, J.; Fredericks, J.; Nishi, S.; Mildiner, S.; Moores, S.L.; Brugge, J.; et al. Vav1/2/3-null mice define an essential role for Vav family proteins in lymphocyte development and activation but a differential requirement in MAPK signaling in T and B cells. J. Exp. Med. 2003, 198, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Charvet, C.; Canonigo, A.J.; Billadeau, D.D.; Altman, A. Membrane localization and function of Vav3 in T cells depend on its association with the adapter SLP-76. J. Biol. Chem. 2005, 280, 15289–15299. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Zmuldzinas, A.; Gardner, S.; Barbacid, M.; Bernstein, A.; Guidos, C. Defective T-cell receptor signalling and positive selection of Vav-deficient CD4+ CD8+ thymocytes. Nature 1995, 374, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, S.; Wu, N.; Wu, L.; Wang, C.; Lin, Y. Overexpression of miR-499-5p inhibits non-small cell lung cancer proliferation and metastasis by targeting VAV3. Sci. Rep. 2016, 6, 23100. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, S.I.; Liu, X.A.; Zhou, W.B.; Ma, R.R.; Chen, L. Vav3 oncogene is upregulated and a poor prognostic factor in breast cancer patients. Oncol. Lett. 2015, 9, 2143–2148. [Google Scholar] [CrossRef] [PubMed]
- Graham, A. Jaw development: Chinless wonders. Curr. Biol. 2002, 12, 810–812. [Google Scholar] [CrossRef]
- Elphick, M.R. The evolution and comparative neurobiology of endocannabinoid signaling. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012, 367, 3201–3215. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.P. Evolution and ecology of retinal photoreception in early vertebrates. Brain Behav. Evol. 2010, 75, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, C.T.; Saha, N.R.; Zapata, A. Evolution and development of immunological structures in the lamprey. Curr. Opin. Immunol. 2007, 19, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Pancer, Z.; Amemiya, C.T.; Ehrhardt, G.R.; Ceitlin, J.; Gartland, G.L.; Cooper, M.D. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature 2004, 430, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Hirano, M.; Herrin, B.R.; Li, J.; Yu, C.; Sadlonova, A.; Cooper, M.D. Dual nature of the adaptive immune system in lampreys. Nature 2009, 459, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Hirano, M.; Guo, P.; McCurley, N.; Schorpp, M.; Das, S.; Boehm, T.; Cooper, M.D. Evolutionary implications of a third lymphocyte lineage in lampreys. Nature 2013, 501, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Herrin, B.R.; Alder, M.N.; Roux, K.H.; Sina, C.; Ehrhardt, G.R.; Boydston, J.A.; Turnbough, C.L., Jr.; Cooper, M.D. Structure and specificity of lamprey monoclonal antibodies. Proc. Natl. Acad. Sci. USA 2008, 105, 2040–2045. [Google Scholar] [CrossRef] [PubMed]
- Alder, M.N.; Herrin, B.R.; Sadlonova, A.; Stockard, C.R.; Grizzle, W.E.; Gartland, L.A.; Gartland, G.L.; Boydston, J.A.; Turnbough, C.L., Jr.; Cooper, M.D. Antibody responses of variable lymphocyte receptors in the lamprey. Nat. Immunol. 2008, 9, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Holland, S.J.; Gao, M.; Hirano, M.; Iyer, L.M.; Luo, M.; Schorpp, M.; Cooper, M.D.; Aravind, L.; Mariuzza, R.A.; Boehm, T. Selection of the lamprey VLRC antigen receptor repertoire. Proc. Natl. Acad. Sci. USA 2014, 111, 14834–14839. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.S.; Walters, A.E.; Mee, P.J.; Turner, M.; Reynolds, L.F.; Prisco, A.; Sarner, N.; Zamoyska, R.; Tybulewicz, V.L. The Rho-family GTP exchange factor Vav is a critical transducer of T cell receptor signals to the calcium, ERK, and NF-kappa B pathways. Proc. Natl. Acad. Sci. USA 1999, 96, 3035–3040. [Google Scholar] [CrossRef] [PubMed]
- Doody, G.M.; Billadeau, D.D.; Clayton, E.; Hutchings, A.; Berland, R.; McAdam, S.; Leibson, P.J.; Turner, M. Vav-2 controls NFAT-dependent transcription in B- but not T-lymphocytes. EMBO J. 2000, 19, 6173–6184. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R. Vav family exchange factors: An integrated regulatory and functional view. Small GTPases 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Uinuk, -O.T.; Mayer, W.E.; Sato, A.; Dongak, R.; Cooper, M.D.; Klein, J. Lamprey lymphocyte-like cells express homologs of genes involved in immunologically relevant activities of mammalian lymphocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 14356–14361. [Google Scholar]
- Pancer, Z.; Cooper, M.D. The evolution of adaptive immunity. Annu. Rev. Immunol. 2006, 24, 497–518. [Google Scholar] [CrossRef] [PubMed]
- Bajoghli, B.; Guo, P.; Aghaallaei, N.; Hirano, M.; Strohmeier, C.; McCurley, N.; Bockman, D.E.; Schorpp, M.; Cooper, M.D.; Boehm, T. A thymus candidate in lampreys. Nature 2011, 470, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, J.; Chen, L.; Liu, X.; Su, P.; Han, Y.; Feng, B.; Li, Q. A novel BTK-like protein involved in immune response in Lethenteron japonicum. Immunol. Lett. 2012, 146, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, X.; Yu, T.; Shi, B.; Xiao, R.; Pang, Y.; Li, Q. A novel member of B-cell linker protein identified in lamprey, Lampetra japonica. Acta Biochim. Biophys. Sin. 2014, 46, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, X.; Shi, B.; Xiao, R.; Gou, M.; Wang, H.; Li, Q. Identification and characterisation of the immune response properties of Lampetra japonica BLNK. Sci. Rep. 2016, 6, 25308. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Su, P.; Li, R.; Zhang, Q.; Zhu, T.; Liu, X.; Li, Q. Molecular cloning, expression pattern and molecular evolution of the spleen tyrosine kinase in lamprey, Lampetra japonica. Dev. Genes Evol. 2015, 225, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Su, P.; Liu, C.; Zhang, Q.; Zhu, T.; Pang, Y.; Liu, X.; Li, Q. A novel protein tyrosine kinase Tec identified in lamprey, Lampetra japonica. Acta. Biochim. Biophys. Sin. 2015, 47, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, X.; Wu, Y.; Ma, F.; Wang, J.; Li, Q. Separation and cytological character of peripheral blood lymphocytes in Japanese lamprey. Chin. J. Zool. 2008, 43, 82–87. [Google Scholar]
- George, J.C.; Beamish, F.W. Haemocytology of the supraneural myeloid body in the sea lamprey during several phases of life cycle. Cana. J. Zool. 1974, 52, 1585–1589. [Google Scholar] [CrossRef]
- Zhang, X.; Song, X.; Su, P.; Gou, M.; Wang, H.; Liu, X.; Li, Q. Molecular cloning, expression pattern, and phylogenetic analysis of a tetraspanin CD82-like molecule in lamprey Lampetra japonica. Dev. Genes Evol. 2016, 226, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.T.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chen, L.; Liu, X.; Wang, H.; Su, P.; Han, Y.; Feng, B.; Qiao, X.; Zhao, J.; Ma, N.; Liu, H.; Zheng, Z.; Li, Q. Lamprey variable lymphocyte receptors mediate complement-dependent cytotoxicity. J. Immunol. 2013, 190, 922–930. [Google Scholar] [CrossRef] [PubMed]











| Motifs | Width | Best Possible Match | Motifs | Width | Best Possible Match |
|---|---|---|---|---|---|
| 1 | 20 | QGWWKGEVNGRVGWFPSTYV | 2 | 20 | TNCKACQMLLRGTFYQGYLC |
| 3 | 20 | FCARDMRELSLREGDVVKIY | 4 | 20 | LVEYYQHHSLKEGFRQLDTT |
| 5 | 20 | YAISIKFNNEVKHIKIVTKD | 6 | 20 | DYSAYPWFAGNMERQQADNE |
| 7 | 20 | WWQGRNLQTQKVGYFPSDAV | 8 | 20 | SKCGAGAHKECLEIIDNCKM |
| 9 | 20 | PPLHFQTGDVIELLRGDPHS | 10 | 20 | DPGLPKMQAIQNYHGIPAPP |
| 11 | 20 | RTRSPVFTPRVIGIAIARYD | 12 | 20 | LINHVNGTYLIRHRTAEAEE |
| 13 | 16 | NWFHITENKKFKSLME | 14 | 12 | LQYPYKERENST |
| 15 | 9 | HNFQMHTFD | 16 | 9 | KPCPCDPKP |
| 17 | 9 | HPHDMDTNG | 18 | 7 | IRPPSRE |
| 19 | 7 | SKIGGDQ | 20 | 5 | EEEGV |
| 21 | 5 | TKMSA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Zhang, Y.; Han, Y.; Su, P.; Gou, M.; Pang, Y.; Li, Q.; Liu, X. A Novel Vav3 Homolog Identified in Lamprey, Lampetra japonica, with Roles in Lipopolysaccharide-Mediated Immune Response. Int. J. Mol. Sci. 2017, 18, 2035. https://doi.org/10.3390/ijms18102035
Shen Y, Zhang Y, Han Y, Su P, Gou M, Pang Y, Li Q, Liu X. A Novel Vav3 Homolog Identified in Lamprey, Lampetra japonica, with Roles in Lipopolysaccharide-Mediated Immune Response. International Journal of Molecular Sciences. 2017; 18(10):2035. https://doi.org/10.3390/ijms18102035
Chicago/Turabian StyleShen, Yanqi, Yishan Zhang, Yinglun Han, Peng Su, Meng Gou, Yue Pang, Qingwei Li, and Xin Liu. 2017. "A Novel Vav3 Homolog Identified in Lamprey, Lampetra japonica, with Roles in Lipopolysaccharide-Mediated Immune Response" International Journal of Molecular Sciences 18, no. 10: 2035. https://doi.org/10.3390/ijms18102035
APA StyleShen, Y., Zhang, Y., Han, Y., Su, P., Gou, M., Pang, Y., Li, Q., & Liu, X. (2017). A Novel Vav3 Homolog Identified in Lamprey, Lampetra japonica, with Roles in Lipopolysaccharide-Mediated Immune Response. International Journal of Molecular Sciences, 18(10), 2035. https://doi.org/10.3390/ijms18102035