Aptamers for DNA Damage and Repair

Abstract
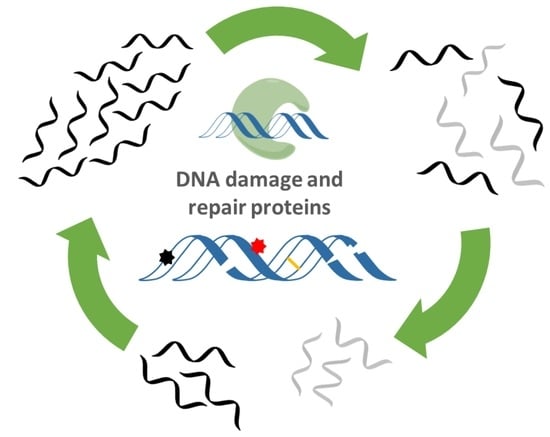
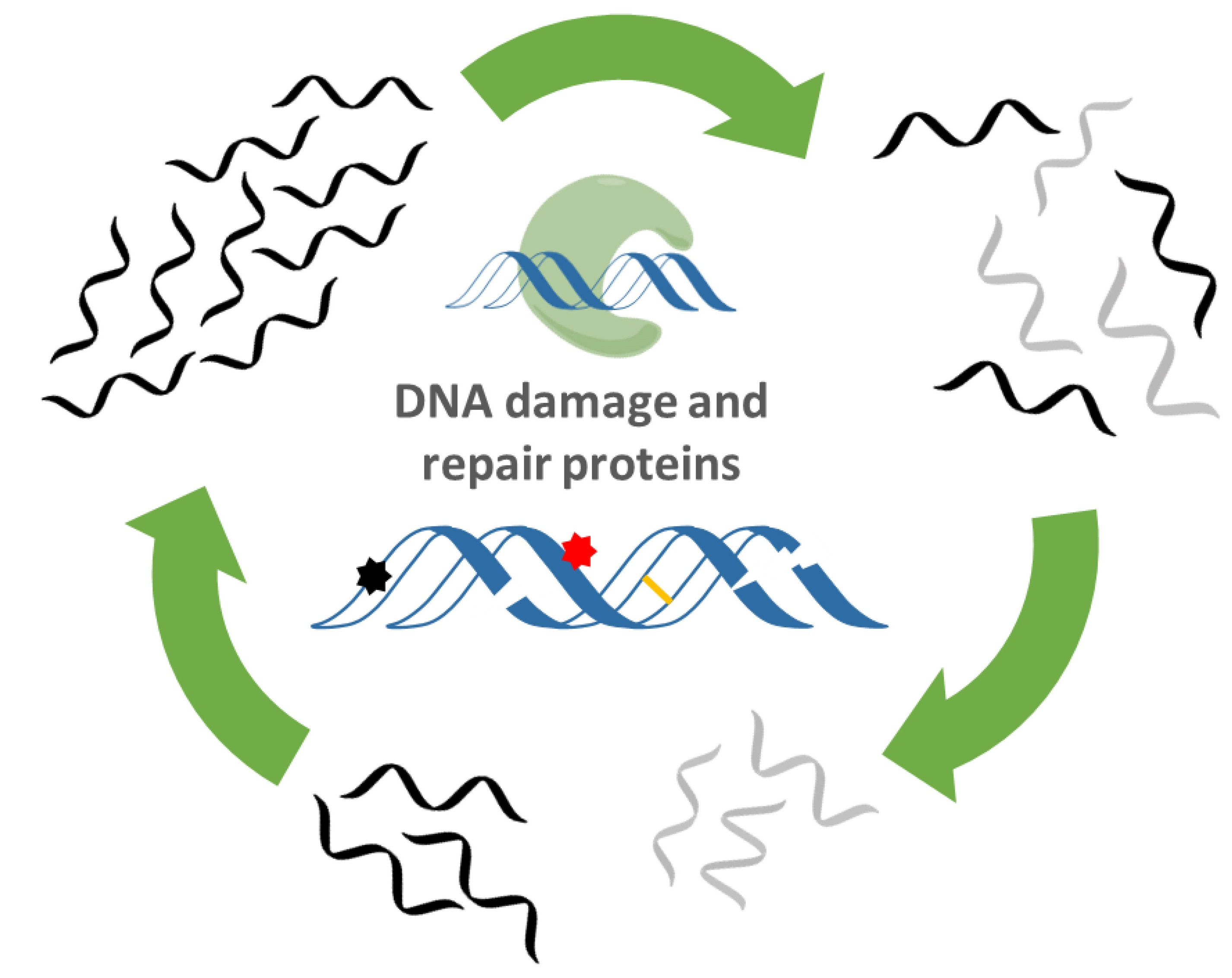
:
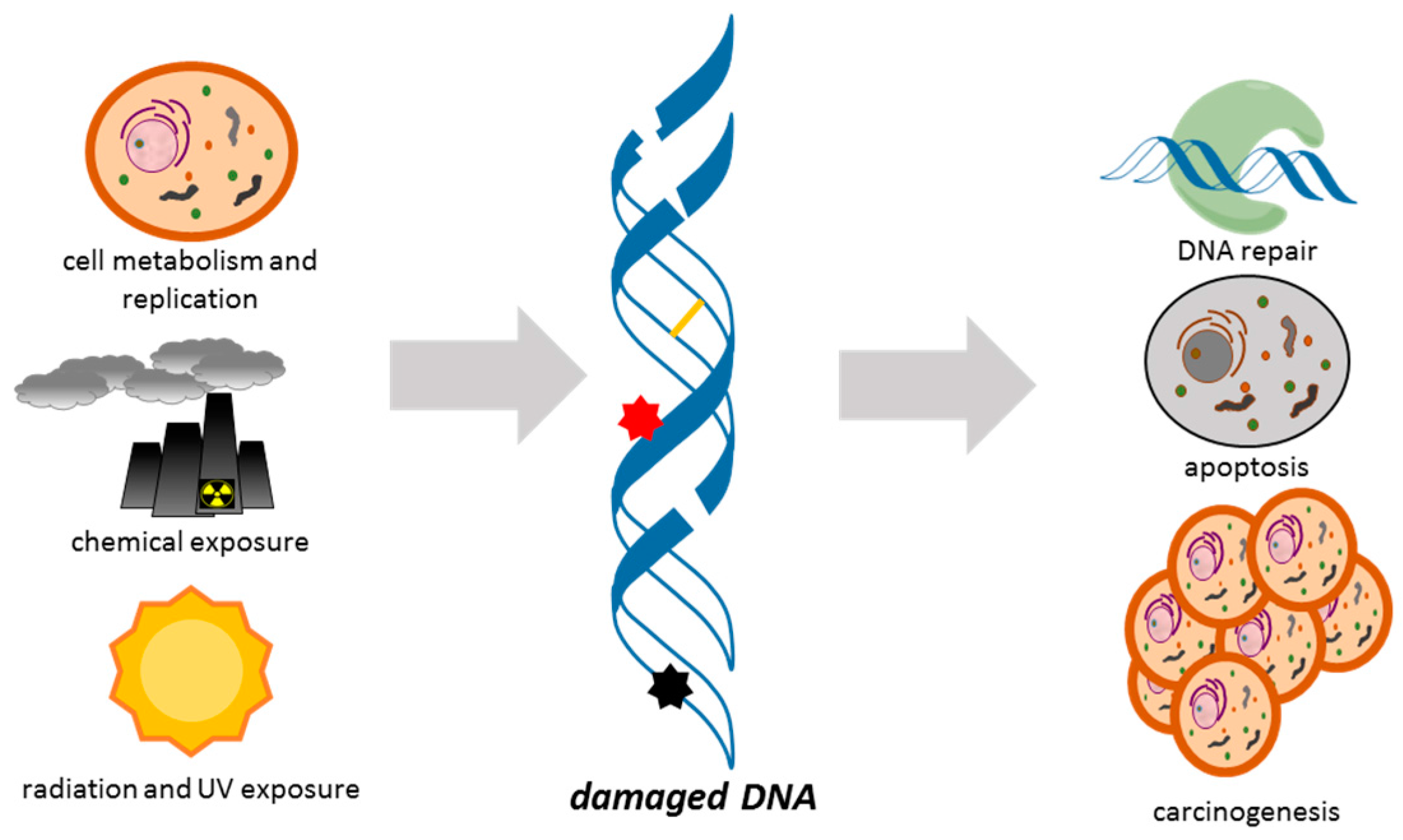
1. Introduction
2. Aptamers for Damaged DNA
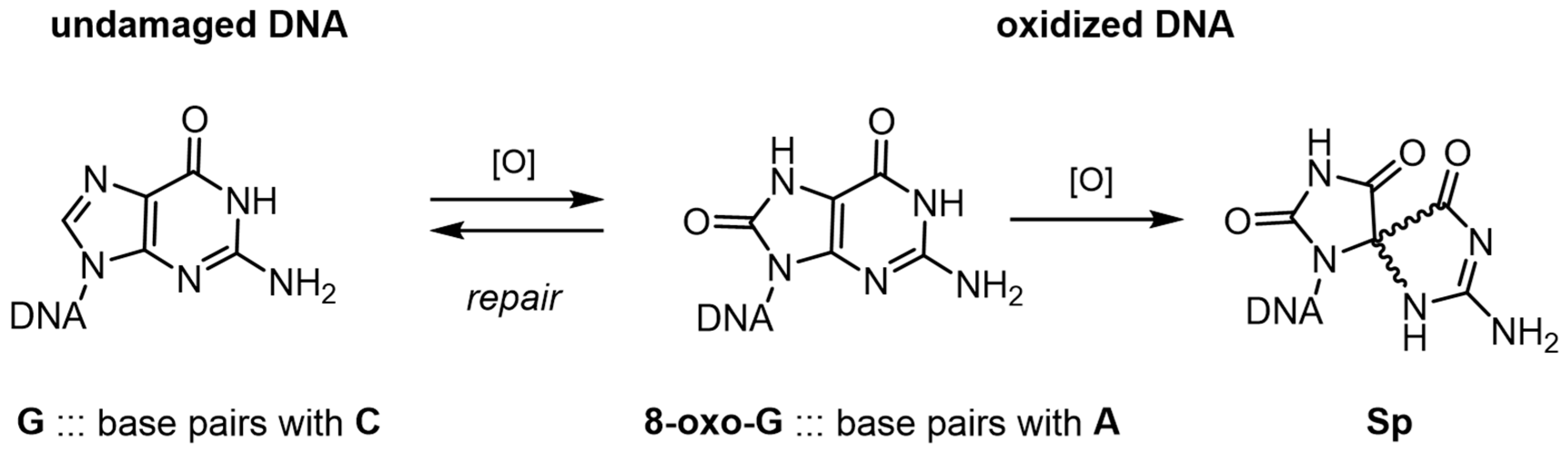
2.1. Guanine Oxidation
2.2. Guanine Alkylation
2.3. Double-Strand Breaks
3. Aptamers for Repair Proteins
3.1. Non-Homologous End Joining
3.2. Base Excision Repair
3.3. Mismatch Repair
3.4. Direct Repair
4. Aptamers That Recognize Mutated Gene Products
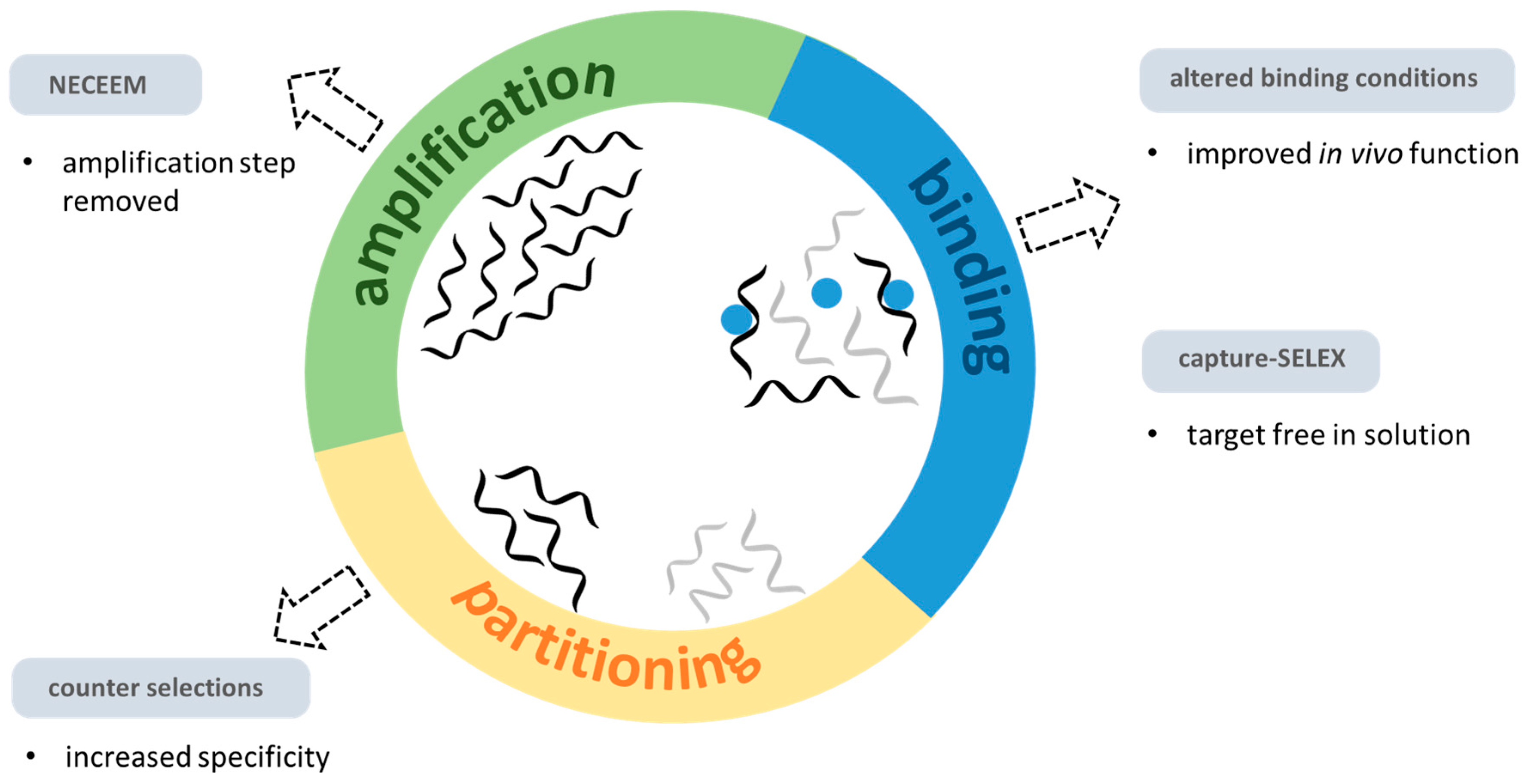
5. Selection Challenges and Considerations
5.1. DNA Adducts
5.2. Strand Breaks
5.3. Proteins
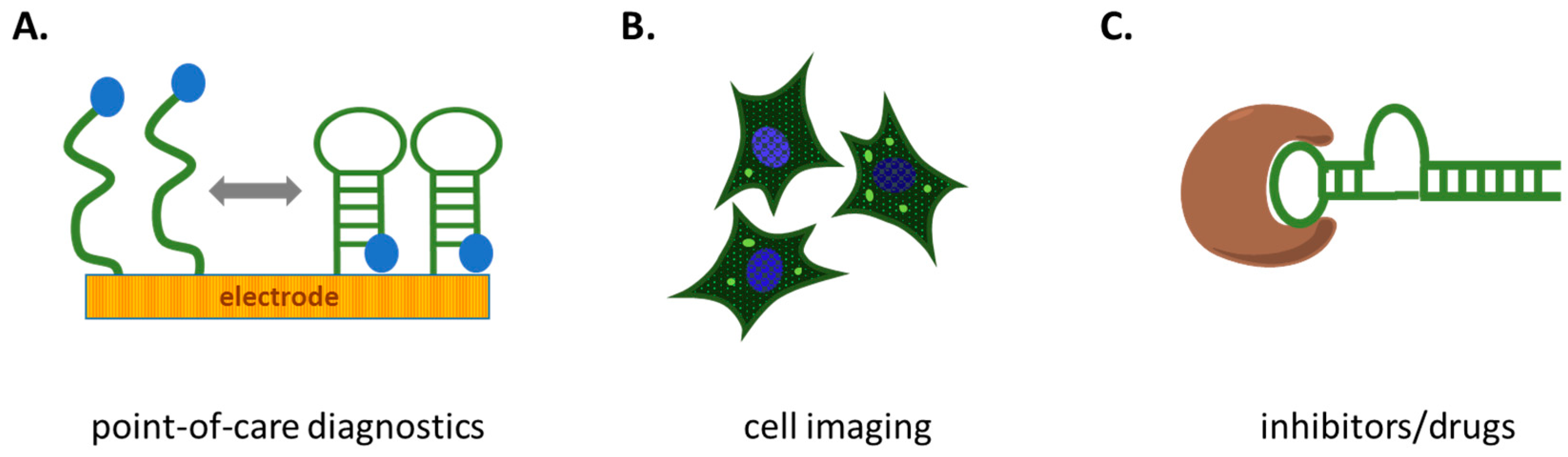
6. Promising Applications
6.1. Diagnostics
6.2. Cellular Imaging
6.3. Therapeutic Targets
6.4. Application Roadblocks
7. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| ABH2 | AlkB homologue 2 |
| AGT | O6-alkylguanine DNA alkyltransferase |
| BER | Base excision repair |
| BIQ | Benzoindoloquinoline |
| bp | Base pair |
| DSB | Double-strand break |
| dsDNA | Double-strand DNA |
| EMSA | Electrophoretic mobility shift assay |
| Fpg | Formamidopyrimidine DNA glycosylase |
| GA | Glycidamide |
| HR | Homologous recombination |
| Kd | Dissociation constant |
| LTR | Long terminal repeat |
| m7-GTP | 7-methylguanosine 5′-triphosphate |
| NECEEM | Non-equilibrium capillary electrophoresis of equilibrium mixtures |
| NER | Nucleotide excision repair |
| NHEJ | Non non-homologous end joining |
| N7-meG | N7-methylguanine |
| 8-oxoG | 8-oxoguanine |
| PNA | Peptide nucleic acid |
| SELEX | Systematic evolution of ligands by EXPonential enrichment |
| Sp | Spiroiminodihydantoin |
References
- Lindahl, T.; Andersson, A. Rate of chain breakage at apurinic sites in double-stranded deoxyribonucleic acid. Biochemistry 1972, 11, 3618–3623. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Nyberg, B. Rate of depurination of native deoxyribonucleic acid. Biochemistry 1972, 11, 3610–3618. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Nyberg, B. Heat-induced deamination of cytosine residues in deoxyribonucleic acid. Biochemistry 1974, 13, 3405–3410. [Google Scholar] [CrossRef] [PubMed]
- Knoch, J.; Kamenisch, Y.; Kubisch, C.; Berneburg, M. Rare hereditary diseases with defects in DNA-repair. Eur. J. Dermatol. 2012, 22, 443–455. [Google Scholar] [PubMed]
- Leone, G.; Pagano, L.; Ben-Yehuda, D.; Voso, M.T. Therapy-related leukemia and myelodysplasia: Susceptibility and incidence. Haematologica 2007, 92, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.E.; Vogel, V.; Ng, A.; Foxhall, L.; Goodwin, P.; Travis, L.B. Second malignant neoplasms: Assessment and strategies for risk reduction. J. Clin. Oncol. 2012, 30, 3734–3745. [Google Scholar] [CrossRef] [PubMed]
- Santella, R.M.; Yang, X.Y.; Hsieh, L.L.; Young, T.L. Immunologic methods for the detection of carcinogen adducts in humans. Prog. Clin. Biol. Res. 1990, 340C, 247–257. [Google Scholar] [PubMed]
- Shia, J.; Ellis, N.A.; Klimstra, D.S. The utility of immunohistochemical detection of DNA mismatch repair gene proteins. Virchows Arch. 2004, 445, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yang, S. Replacing antibodies with aptamers in lateral flow immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.Y.; Citartan, M.; Gopinath, S.C.; Tang, T.H. Aptamers as a replacement for antibodies in enzyme-linked immunosorbent assay. Biosens. Bioelectron. 2015, 64, 392–403. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Derosa, M.C. Challenges and opportunities for small molecule aptamer development. J. Nucleic Acids 2012, 2012, 748913. [Google Scholar] [CrossRef] [PubMed]
- Voskuil, J.L. The challenges with the validation of research antibodies. F1000Research 2017, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kamiya, H. Mutations induced by 8-hydroxyguanine (8-oxo-7,8-dihydroguanine), a representative oxidized base, in mammalian cells. Genes Environ. 2017, 39, 2. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Ding, Y.; Burrows, C.J. Sequencing DNA for the oxidatively modified base 8-oxo-7,8-dihydroguanine. Methods Enzymol. 2017, 591, 187–210. [Google Scholar] [PubMed]
- Alenko, A.; Fleming, A.M.; Burrows, C.J. Reverse transcription past products of guanine oxidation in rna leads to insertion of a and c opposite 8-oxo-7,8-dihydroguanine and a and g opposite 5-guanidinohydantoin and spiroiminodihydantoin diastereomers. Biochemistry 2017, 56, 5053–5064. [Google Scholar] [CrossRef] [PubMed]
- Rink, S.M.; Shen, J.C.; Loeb, L.A. Creation of rna molecules that recognize the oxidative lesion 7,8-dihydro-8-hydroxy-2’-deoxyguanosine (8-oxodg) in DNA. Proc. Natl. Acad. Sci. USA 1998, 95, 11619–11624. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, Y.; Shimizu, N.; Ogino, C.; Fukuda, H.; Kondo, A. Selection of a DNA aptamer that binds 8-ohdg using gmp-agarose. Bioorg. Med. Chem. Lett. 2009, 19, 3619–3622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.; Meng, X.; Dhar, R.; Huang, H. Triple-stranded DNA containing 8-oxo-7,8-dihydro-2′-deoxyguanosine: Implication in the design of selective aptamer sensors for 8-oxo-7,8-dihydroguanine. Anal. Chem. 2013, 85, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Chirania, P.; Ganguly, S.; Huang, H. A DNA aptamer sensor for 8-oxo-7,8-dihydroguanine. Bioorg. Med. Chem. Lett. 2012, 22, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Nikolaus, N.; Strehlitz, B. Capture-selex: Selection of DNA aptamers for aminoglycoside antibiotics. J. Anal. Methods Chem. 2012, 2012, 415697. [Google Scholar] [CrossRef] [PubMed]
- Ghude, P.; Burrows, C.J. Structure-Switching Selex for Selection of Aptamers of Damaged Nucleosides and Nucleobases 8-Oxoguanine and Spiroiminodihydantoin. Ph.D. Thesis, The University of Utah, Salt Lake City, UT, USA, 2015. [Google Scholar]
- Haller, A.A.; Sarnow, P. In vitro selection of a 7-methyl-guanosine binding rna that inhibits translation of capped mrna molecules. Proc. Natl. Acad. Sci. USA 1997, 94, 8521–8526. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Carrocci, T.J.; Hoskins, A.A. Evolution and characterization of a benzylguanine-binding rna aptamer. Chem. Commun. 2016, 52, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Soukup, G.A.; Ellington, A.D.; Maher, L.J., III. Selection of rnas that bind to duplex DNA at neutral ph. J. Mol. Biol. 1996, 259, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Ayel, E.; Escude, C. In vitro selection of oligonucleotides that bind double-stranded DNA in the presence of triplex-stabilizing agents. Nucleic Acids Res. 2010, 38, e31. [Google Scholar] [CrossRef] [PubMed]
- Srisawat, C.; Engelke, D.R. Selection of RNA aptamers that bind hiv-1 ltr DNA duplexes: Strand invaders. Nucleic Acids Res. 2010, 38, 8306–8315. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.; Dynan, W.S. Characterization of the RNA binding properties of ku protein. Biochemistry 1998, 37, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Vuyisich, M.; Beal, P.A. Controlling protein activity with ligand-regulated RNA aptamers. Chem. Biol. 2002, 9, 907–913. [Google Scholar] [CrossRef]
- Gening, L.V.; Klincheva, S.A.; Reshetnjak, A.; Grollman, A.P.; Miller, H. Rna aptamers selected against DNA polymerase beta inhibit the polymerase activities of DNA polymerases beta and kappa. Nucleic Acids Res. 2006, 34, 2579–2586. [Google Scholar] [CrossRef] [PubMed]
- Drabovich, A.P.; Berezovski, M.; Okhonin, V.; Krylov, S.N. Selection of smart aptamers by methods of kinetic capillary electrophoresis. Anal. Chem. 2006, 78, 3171–3178. [Google Scholar] [CrossRef] [PubMed]
- Krylova, S.M.; Karkhanina, A.A.; Musheev, M.U.; Bagg, E.A.; Schofield, C.J.; Krylov, S.N. DNA aptamers for as analytical tools for the quantitative analysis of DNA-dealkylating enzymes. Anal. Biochem. 2011, 414, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Yufa, R.; Krylova, S.M.; Bruce, C.; Bagg, E.A.; Schofield, C.J.; Krylov, S.N. Emulsion pcr significantly improves nonequilibrium capillary electrophoresis of equilibrium mixtures-based aptamer selection: Allowing for efficient and rapid selection of aptamer to unmodified ABH2 protein. Anal. Chem. 2015, 87, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Han, S.R.; Lee, Y.J.; Kim, J.H.; Lee, S.W. Identification of rna aptamer specific to mutant kras protein. Oligonucleotides 2010, 20, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Calvo, J.A.; Samson, L.D. Balancing repair and tolerance of DNA damage caused by alkylating agents. Nat. Rev. Cancer 2012, 12, 104–120. [Google Scholar] [CrossRef] [PubMed]
- Kiga, D.; Futamura, Y.; Sakamoto, K.; Yokoyama, S. An RNA aptamer to the xanthine/guanine base with a distinctive mode of purine recognition. Nucleic Acids Res. 1998, 26, 1755–1760. [Google Scholar] [CrossRef] [PubMed]
- Larguinho, M.; Santos, S.; Almeida, J.; Baptista, P.V. DNA adduct identification using gold-aptamer nanoprobes. IET Nanobiotechnol. 2015, 9, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Wyss, L.A.; Nilforoushan, A.; Eichenseher, F.; Suter, U.; Blatter, N.; Marx, A.; Sturla, S.J. Specific incorporation of an artificial nucleotide opposite a mutagenic DNA adduct by a DNA polymerase. J. Am. Chem. Soc. 2015, 137, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Scharer, O.D. Chemistry and biology of DNA repair. Angew. Chem. Int. Ed. Engl. 2003, 42, 2946–2974. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, T.S. DNA repair pathways and mechanisms. In DNA Repair of Cancer Stem Cells; Mathews, L.A., Cabarcas, S.M., Hurt, E.M., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 19–32. [Google Scholar]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [PubMed]
- Pei, D.S.; Strauss, P.R. Zebrafish as a model system to study DNA damage and repair. Mutat. Res. 2013, 743–744, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Mondragon, E.; Maher, L.J., III. Anti-transcription factor RNA aptamers as potential therapeutics. Nucleic Acid Ther. 2016, 26, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Fattah, F.; Lee, E.H.; Weisensel, N.; Wang, Y.; Lichter, N.; Hendrickson, E.A. Ku regulates the non-homologous end joining pathway choice of DNA double-strand break repair in human somatic cells. PLoS Genet. 2010, 6, e1000855. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, X. Microhomology-mediated end joining: New players join the team. Cell Biosci. 2017, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Durdikova, K.; Chovanec, M. Regulation of non-homologous end joining via post-translational modifications of components of the ligation step. Curr. Genet. 2016, 63, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Beard, W.A.; Prasad, R.; Wilson, S.H. Activities and mechanism of DNA polymerase beta. Methods Enzymol. 2006, 408, 91–107. [Google Scholar] [PubMed]
- Idriss, H.T.; Al-Assar, O.; Wilson, S.H. DNA polymerase beta. Int. J. Biochem. Cell Biol. 2002, 34, 321–324. [Google Scholar] [CrossRef]
- Fishel, R. Mismatch repair. J. Biol. Chem. 2015, 290, 26395–26403. [Google Scholar] [CrossRef] [PubMed]
- Li, G.M. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008, 18, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Musheev, M.U.; Krylov, S.N. Selection of aptamers by systematic evolution of ligands by exponential enrichment: Addressing the polymerase chain reaction issue. Anal. Chim. Acta 2006, 564, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; He, C. DNA repair by reversal of DNA damage. Cold Spring Harb. Perspect. Biol. 2013, 5, a012575. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, T.C.; Prestegarden, L.; Grudic, A.; Hegi, M.E.; Tysnes, B.B.; Bjerkvig, R. The DNA repair protein alkbh2 mediates temozolomide resistance in human glioblastoma cells. Neuro Oncol. 2013, 15, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Chen, B.; Qi, B.; Zhang, W.; Jia, G.; Zhang, L.; Li, C.J.; Dinner, A.R.; Yang, C.G.; He, C. Duplex interrogation by a direct DNA repair protein in search of base damage. Nat. Struct. Mol. Biol. 2012, 19, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Krylova, S.M.; Koshkin, V.; Bagg, E.; Schofield, C.J.; Krylov, S.N. Mechanistic studies on the application of DNA aptamers as inhibitors of 2-oxoglutarate-dependent oxygenases. J. Med. Chem. 2012, 55, 3546–3552. [Google Scholar] [CrossRef] [PubMed]
- Tokheim, C.J.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B.; Karchin, R. Evaluating the evaluation of cancer driver genes. Proc. Natl. Acad. Sci. USA 2016, 113, 14330–14335. [Google Scholar] [CrossRef] [PubMed]
- Adar, S.; Hu, J.; Lieb, J.D.; Sancar, A. Genome-wide kinetics of DNA excision repair in relation to chromatin state and mutagenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E2124–E2133. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.T.; Asthana, S.; Gao, S.P.; Lee, B.H.; Chapman, J.S.; Kandoth, C.; Gao, J.; Socci, N.D.; Solit, D.B.; Olshen, A.B.; et al. Identifying recurrent mutations in cancer reveals widespread lineage diversity and mutational specificity. Nat. Biotechnol. 2016, 34, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, B.A.; Sha, M.; Wathen, S.T.; Rando, R.R. RNA aptamers that specifically bind to a k ras-derived farnesylated peptide. Bioorg. Med. Chem. 1997, 5, 1115–1122. [Google Scholar] [CrossRef]
- Tanaka, Y.; Akagi, K.; Nakamura, Y.; Kozu, T. RNA aptamers targeting the carboxyl terminus of kras oncoprotein generated by an improved selex with isothermal rna amplification. Oligonucleotides 2007, 17, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, G.P.; Denissenko, M.F.; Olivier, M.; Tretyakova, N.; Hecht, S.S.; Hainaut, P. Tobacco smoke carcinogens, DNA damage and p53 mutations in smoking-associated cancers. Oncogene 2002, 21, 7435–7451. [Google Scholar] [CrossRef] [PubMed]
- Hang, B. Formation and repair of tobacco carcinogen-derived bulky DNA adducts. J. Nucleic Acids 2010, 2010, 709521. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Rashid, F.; Shah, A.; Awan, H.M.; Wu, M.; Liu, A.; Wang, J.; Zhu, T.; Luo, Z.; Shan, G. The isolation of an rna aptamer targeting to p53 protein with single amino acid mutation. Proc. Natl. Acad. Sci. USA 2015, 112, 10002–10007. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage t4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of rna molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Joyce, G.F. Selection in vitro of an RNA enzyme that specifically cleaves single-stranded DNA. Nature 1990, 344, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Jijakli, K.; Khraiwesh, B.; Fu, W.; Luo, L.; Alzahmi, A.; Koussa, J.; Chaiboonchoe, A.; Kirmizialtin, S.; Yen, L.; Salehi-Ashtiani, K. The in vitro selection world. Methods 2016, 106, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zu, Y. A highlight of recent advances in aptamer technology and its application. Molecules 2015, 20, 11959–11980. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; McConnell, E.M.; Cruz-Toledo, J.; Bernard, E.D.; Pach, A.; Mastronardi, E.; Zhang, X.; Beking, M.; Francis, T.; Giamberardino, A.; et al. Analysis of in vitro aptamer selection parameters. J. Mol. Evol. 2015, 81, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, M. Selex experiments: New prospects, applications and data analysis in inferring regulatory pathways. Biomol. Eng. 2007, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Huang, R.; Deng, Y.; He, N. Progress in selection and biomedical applications of aptamers. J. Biomed. Nanotechnol. 2014, 10, 3043–3062. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Esposito, C.L. Aptamer cell-based selection: Overview and advances. Biomedicines 2017, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Ruscito, A.; McConnell, E.M.; Koudrian, A.; Velu, R.; Mattic, C.; Hung, V.; McKeague, M.; DeRosa, M.C. In vitro selection and characterization of DNA aptamers to a small molecule target. Curr. Protoc. Chem. Biol. 2017, 9, 1–36. [Google Scholar]
- Yang, K.A.; Pei, R.; Stojanovic, M.N. In vitro selection and amplification protocols for isolation of aptameric sensors for small molecules. Methods 2016, 106, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Nutiu, R.; Li, Y. Structure-switching signaling aptamers. J. Am. Chem. Soc. 2003, 125, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.E.; Egholm, M.; Berg, R.H.; Buchardt, O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Peffer, N.J.; Hanvey, J.C.; Bisi, J.E.; Thomson, S.A.; Hassman, C.F.; Noble, S.A.; Babiss, L.E. Strand-invasion of duplex DNA by peptide nucleic acid oligomers. Proc. Natl. Acad. Sci. USA 1993, 90, 10648–10652. [Google Scholar] [CrossRef] [PubMed]
- Scheibe, C.; Wedepohl, S.; Riese, S.B.; Dernedde, J.; Seitz, O. Carbohydrate-pna and aptamer-pna conjugates for the spatial screening of lectins and lectin assemblies. Chembiochem 2013, 14, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Brudno, Y.; Birnbaum, M.E.; Kleiner, R.E.; Liu, D.R. An in vitro translation, selection and amplification system for peptide nucleic acids. Nat. Chem. Biol. 2010, 6, 148–155. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Derosa, M.C. Aptamers and selex: Tools for the development of transformative molecular recognition technology. Aptamers Synth. Antib. 2014, 1, 12–16. [Google Scholar]
- McKeague, M.; Velu, R.; De Girolamo, A.; Valenzano, S.; Pascale, M.; Smith, M.; DeRosa, M.C. Comparison of in-solution biorecognition properties of aptamers against ochratoxin A. Toxins 2016, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.M.; Holahan, M.R.; DeRosa, M.C. Aptamers as promising molecular recognition elements for diagnostics and therapeutics in the central nervous system. Nucleic Acid Ther. 2014, 24, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Kruspe, S.; Giangrande, P.H. Aptamer-sirna chimeras: Discovery, progress, and future prospects. Biomedicines 2017, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Suess, B. Identification of rna aptamers with riboswitching properties. Methods 2016, 97, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Klaene, J.J.; Sharma, V.K.; Glick, J.; Vouros, P. The analysis of DNA adducts: The transition from (32)p-postlabeling to mass spectrometry. Cancer Lett. 2013, 334, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.H.; Arlt, V.M. Genotoxicity: Damage to DNA and its consequences. EXS 2009, 99, 87–110. [Google Scholar] [PubMed]
- Guo, J.; Yun, B.H.; Upadhyaya, P.; Yao, L.; Krishnamachari, S.; Rosenquist, T.A.; Grollman, A.P.; Turesky, R.J. Multiclass carcinogenic DNA adduct quantification in formalin-fixed paraffin-embedded tissues by ultraperformance liquid chromatography-tandem mass spectrometry. Anal. Chem. 2016, 88, 4780–4787. [Google Scholar] [CrossRef] [PubMed]
- Wyss, L.A.; Nilforoushan, A.; Williams, D.M.; Marx, A.; Sturla, S.J. The use of an artificial nucleotide for polymerase-based recognition of carcinogenic o6-alkylguanine DNA adducts. Nucleic Acids Res. 2016, 44, 6564–6573. [Google Scholar] [CrossRef] [PubMed]
- Riedl, J.; Ding, Y.; Fleming, A.M.; Burrows, C.J. Identification of DNA lesions using a third base pair for amplification and nanopore sequencing. Nat. Commun. 2015, 6, 8807. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fleming, A.M.; Burrows, C.J. Sequencing the mouse genome for the oxidatively modified base 8-oxo-7,8-dihydroguanine by og-seq. J. Am. Chem. Soc. 2017, 139, 2569–2572. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Foster, A.; Miguel, Y.; Giamberardino, A.; Verdin, C.; Chan, J.Y.S.; DeRosa, M.C. Development of a DNA aptamer for direct and selective homocysteine detection in human serum. RSC Adv. 2013, 3, 24415–24422. [Google Scholar] [CrossRef]
- Schoukroun-Barnes, L.R.; Glaser, E.P.; White, R.J. Heterogeneous electrochemical aptamer-based sensor surfaces for controlled sensor response. Langmuir 2015, 31, 6563–6569. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.T.; Bewersdorf, J.; Knight, K.L. Immunofluorescence imaging of DNA damage response proteins: Optimizing protocols for super-resolution microscopy. Methods 2009, 48, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Luise, C.; Nuciforo, P. Immunohistochemistry protocol for γH2AX detection (formalin-fixed paraffin-embedded sections). Protoc. Exch. 2006. [Google Scholar] [CrossRef]
- Paige, J.S.; Wu, K.Y.; Jaffrey, S.R. RNA mimics of green fluorescent protein. Science 2011, 333, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Nilaratanakul, V.; Hauer, D.A.; Griffin, D.E. Development and characterization of sindbis virus with encoded fluorescent rna aptamer spinach2 for imaging of replication and immune-mediated changes in intracellular viral rna. J. Gen. Virol. 2017, 98, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Doyle, F.; Wurz, Z.E.; Tenenbaum, S.A.; Hammond, R.K.; Caplan, J.L.; Meyers, B.C. Fastmir: An RNA-based sensor for in vitro quantification and live-cell localization of small rnas. Nucleic Acids Res. 2017, 45, e130. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Wong, R.S.; Smolke, C.D. Opportunities in the design and application of rna for gene expression control. Nucleic Acids Res. 2016, 44, 2987–2999. [Google Scholar] [CrossRef] [PubMed]
- Alsaafin, A.; McKeague, M. Functional nucleic acids as in vivo metabolite and ion biosensors. Biosens. Bioelectron. 2017, 94, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rossi, J. Aptamers as targeted therapeutics: Current potential and challenges. Nat. Rev. Drug Discov. 2017, 16, 440. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.W.; Shima, D.T.; Calias, P.; Cunningham, E.T., Jr.; Guyer, D.R.; Adamis, A.P. Pegaptanib, a targeted anti-vegf aptamer for ocular vascular disease. Nat. Rev. Drug Discov. 2006, 5, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Poolsup, S.; Kim, C.Y. Therapeutic applications of synthetic nucleic acid aptamers. Curr. Opin. Biotechnol. 2017, 48, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Soll, J.M.; Sobol, R.W.; Mosammaparast, N. Regulation of DNA alkylation damage repair: Lessons and therapeutic opportunities. Trends Biochem. Sci. 2017, 42, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Kelley, M.R.; Logsdon, D.; Fishel, M.L. Targeting DNA repair pathways for cancer treatment: What’s new? Futur. Oncol. 2014, 10, 1215–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, A.D.; Fahrer, J.; Johnson, G.E.; Kaina, B. Theoretical considerations for thresholds in chemical carcinogenesis. Mutat. Res. Rev. Mutat. Res. 2015, 765, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hu, J.; Adebali, O.; Adar, S.; Yang, Y.; Chiou, Y.Y.; Sancar, A. Human genome-wide repair map of DNA damage caused by the cigarette smoke carcinogen benzo[a]pyrene. Proc. Natl. Acad. Sci. USA 2017, 114, 6752–6757. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Selby, C.P.; Adar, S.; Adebali, O.; Sancar, A. Molecular mechanisms and genomic maps of DNA excision repair in escherichia coli and humans. J. Biol. Chem. 2017, 292, 15588–15597. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Wang, Y.H.; Smolke, C.D. In vitro screening and in silico modeling of rna-based gene expression control. ACS Chem. Biol. 2015, 10, 2463–2467. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Class | Target | Nucleic Acid | Kd 1 | Reference |
|---|---|---|---|---|
| DNA Adducts | 8-oxodG | RNA | 270 nM | [16] |
| 8-oxodG | DNA | 100 nM | [17] | |
| 8-oxoG | DNA 2 | 5.5 µM | [19] | |
| 8-oxoG | DNA | 3 nM | [21] | |
| 8-oxodG | DNA | 25 µM | [21] | |
| (−),-(R)-dSp | DNA | 28 nM | [21] | |
| (+),-(S)-dS | DNA | 76 nM | [21] | |
| (−),-(R)-Sp | DNA | 12 nM | [21] | |
| m7-GTP | RNA | 500 nM | [22] | |
| benzylguanine | RNA | 200 nM | [23] | |
| Strand Breaks | homopurine/pyrimidine duplex | RNA | 1 µM | [24] |
| 20 bp duplex | DNA 3 | 43.9 nM | [25] | |
| 3′LTR | RNA | 300 nM | [26] | |
| Ku protein | RNA | 2 nM | [27] | |
| Repair Proteins | Fpg (DNA glycosylase) | RNA | 2.5 nM | [28] |
| Polβ/polκ | RNA | 290 nM | [29] | |
| MutS | DNA | 3.6 nM | [30] | |
| AlkB | DNA | 20 nM | [31] | |
| AlkB homologue 2 | DNA | 85 nM | [32] | |
| Mutated Gene | KRASV12 | RNA | 4.04 nM | [33] |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKeague, M. Aptamers for DNA Damage and Repair. Int. J. Mol. Sci. 2017, 18, 2212. https://doi.org/10.3390/ijms18102212
McKeague M. Aptamers for DNA Damage and Repair. International Journal of Molecular Sciences. 2017; 18(10):2212. https://doi.org/10.3390/ijms18102212
Chicago/Turabian StyleMcKeague, Maureen. 2017. "Aptamers for DNA Damage and Repair" International Journal of Molecular Sciences 18, no. 10: 2212. https://doi.org/10.3390/ijms18102212
APA StyleMcKeague, M. (2017). Aptamers for DNA Damage and Repair. International Journal of Molecular Sciences, 18(10), 2212. https://doi.org/10.3390/ijms18102212