Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood

Abstract
:1. Introduction
2. Results
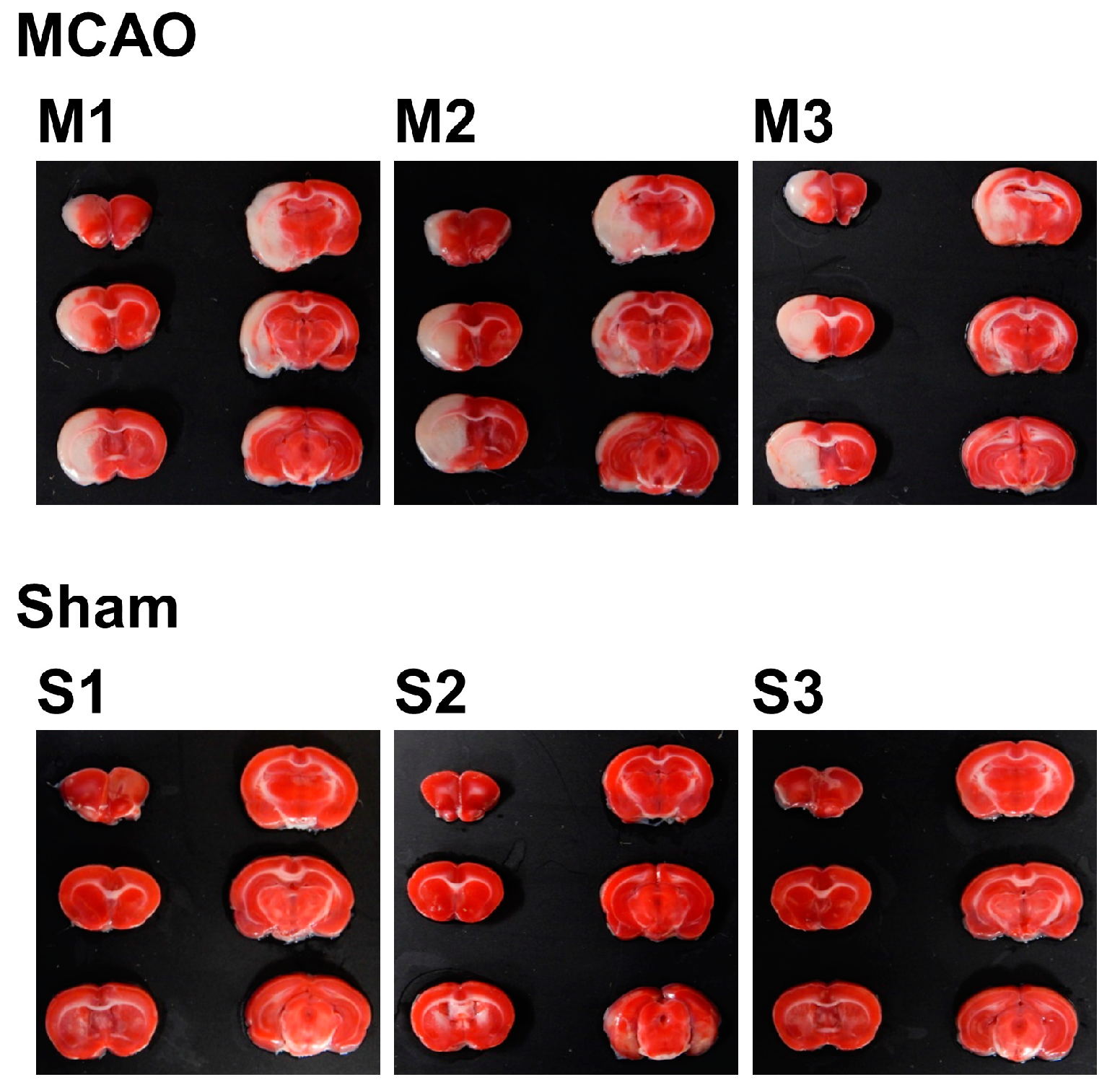
2.1. Internal Condition of the Brain after Stroke
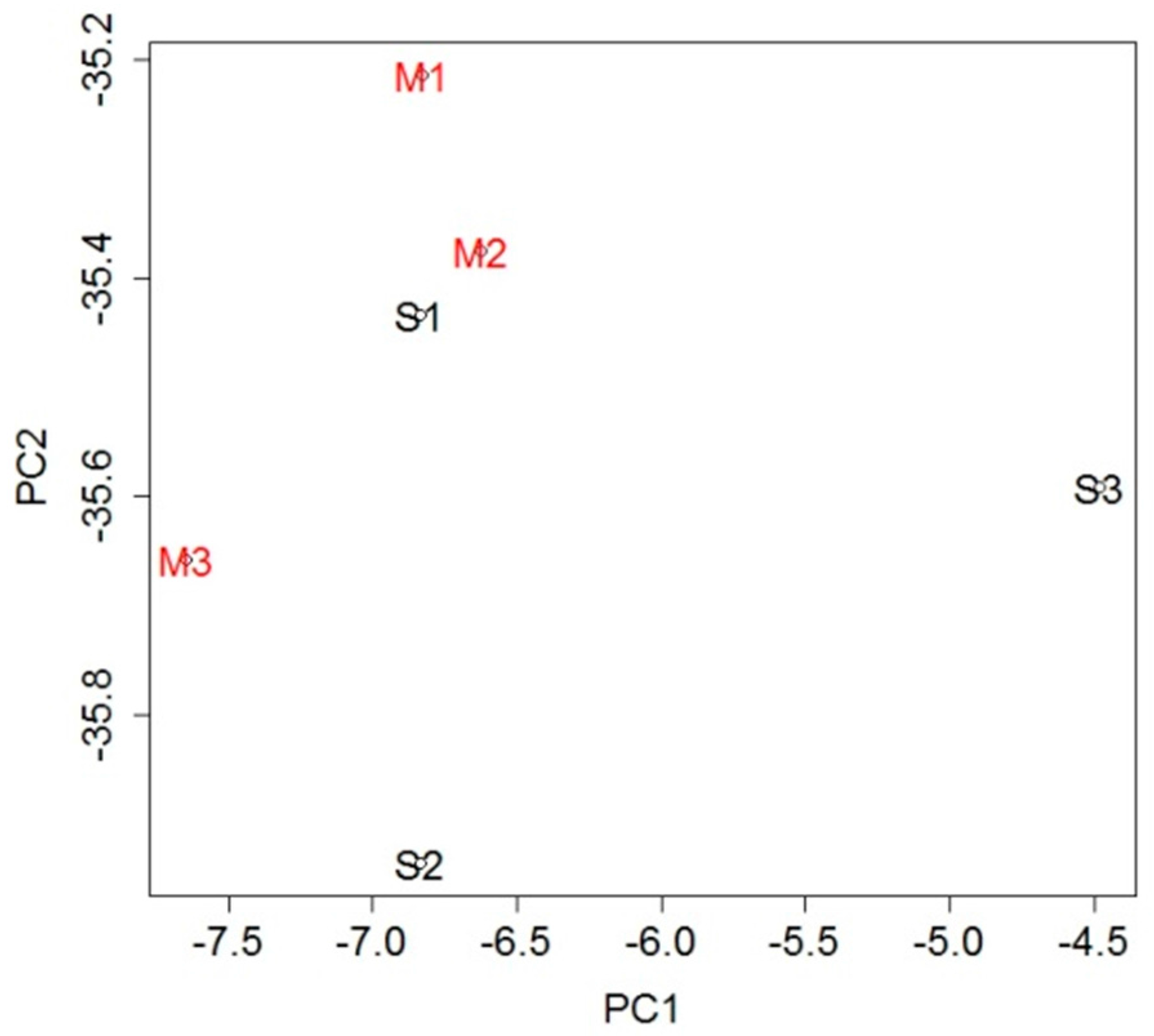
2.2. mRNA Expression in Blood
2.3. miRNA Expression
3. Discussion
3.1. Immunodepression after Stroke
3.2. mRNA–miRNA Correlation
3.3. miRNA as Novel Biomarkers
4. Materials and Methods
4.1. Animals
4.2. MCAO for Blood Samples
4.3. Measurement of Infarct Volume
- Nonedema volume = (contralateral hemisphere) × 2;
- Edema volume = (ipsilateral hemisphere) − (contralateral hemisphere);
- Infarct volume = (contralateral hemisphere) − (noninfarcted part of ipsilateral hemisphere);
- Edema index = (edema volume)/(contralateral hemisphere);
- Infarct index = (infarct volume)/(nonedema volume).
4.4. RNA Isolation
4.5. DNA Microarray Analysis
4.6. miRNA Microarray Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feigin, V.L.; Forouzanfar, M.H.; Krishnamurthi, R.; Mensah, G.A.; Connor, M.; Bennett, D.A.; Moran, A.E.; Sacco, R.L.; Anderson, L.; Truelsen, T. Global and regional burden of stroke during 1990–2010: Findings from the Global Burden of Disease Study 2010. Lancet 2014, 383, 245–255. [Google Scholar] [CrossRef]
- Chamorro, Á.; Meisel, A.; Planas, A.M.; Urra, X.; van de Beek, D.; Veltkamp, R. The immunology of acute stroke. Nat. Rev. Neurol. 2012, 8, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Anrather, J. The immunology of stroke: From mechanisms to translation. Nat. Med. 2011, 17, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Saenger, A.K.; Christenson, R.H. Stroke biomarkers: Progress and challenges for diagnosis, prognosis, differentiation, and treatment. Clin. Chem. 2010, 56, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Liu, J.; Wang, Y.; Wang, L.; Weng, S.; Tang, Y.; Zheng, C.; Cheng, Q.; Chen, S.; Yang, G.-Y. MicroRNA-210 as a novel blood biomarker in acute cerebral ischemia. Front. Biosci. 2011, 3, 1265–1272. [Google Scholar] [CrossRef]
- Liu, D.Z.; Jickling, G.C.; Ander, B.P.; Hull, H.; Zhan, X.; Cox, C.; Shroff, N.; Dykstra-Aiello, C.; Stamova, B.; Sharp, F.R. Elevating microRNA-122 in blood improves outcomes after temporary middle cerebral artery occlusion in rats. J. Cereb. Blood Flow Metab. 2016, 36, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p and miR-143–3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, Y.; Huang, J.; Qu, M.; Zhang, Y.; Geng, J.; Zhang, Z.; Liu, J.; Yang, G.Y. Increased circulating exosomal miRNA-223 is associated with acute ischemic stroke. Front. Neurol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, J.; Yoshida, Y.; Nakazawa, T.; Ooneda, G. Experimental studies of ischemic brain edema 1. A new experimental model of cerebral embolism in rats in which recirculation can be introduced in the ischemic area. Jpn. J. Stroke 1986, 8, 1–8. [Google Scholar] [CrossRef]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.T. Rodent models of focal stroke: Size, mechanism, and purpose. NeuroRx 2005, 2, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Prass, K.; Meisel, C.; Höflich, C.; Braun, J.; Halle, E.; Wolf, T.; Ruscher, K.; Victorov, I.V.; Priller, J.; Dirnagl, U. Stroke-induced immunodeficiency promotes spontaneous bacterial infections and is mediated by sympathetic activation reversal by poststroke T helper cell type 1–like immunostimulation. J. Exp. Med. 2003, 198, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Katzan, I.L.; Cebul, R.D.; Husak, S.H.; Dawson, N.V.; Baker, D.W. The effect of pneumonia on mortality among patients hospitalized for acute stroke. Neurology 2003, 60, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Hilker, R.; Poetter, C.; Findeisen, N.; Sobesky, J.; Jacobs, A.; Neveling, M.; Heiss, W.-D. Nosocomial pneumonia after acute stroke. Stroke 2003, 34, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Prass, K.; Braun, J.S.; Dirnagl, U.; Meisel, C.; Meisel, A. Stroke propagates bacterial aspiration to pneumonia in a model of cerebral ischemia. Stroke 2006, 37, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U.; Klehmet, J.; Braun, J.S.; Harms, H.; Meisel, C.; Ziemssen, T.; Prass, K.; Meisel, A. Stroke-induced immunodepression. Stroke 2007, 38, 770–773. [Google Scholar] [CrossRef] [PubMed]
- Weiss, N.; Miller, F.; Cazaubon, S.; Couraud, P.-O. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim. Biophys. Acta Biomembr. 2009, 1788, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, I.S.; Thomas, M.L. CD45: An emerging role as a protein tyrosine phosphatase required for lymphocyte activation and development. Annu. Rev. Immunol. 1994, 12, 85–116. [Google Scholar] [CrossRef] [PubMed]
- Haijun, Z.; Yonghui, Y.; Jiake, C.; Hongjie, D. Detection of the MicroRNA expression profile in skeletal muscles of burn trauma at the early stage in rats. Ulus Travma Acil Cerrahi Derg 2015, 21, 241–247. [Google Scholar] [PubMed]
- Zhao, R.; Qian, L.; Jiang, L. Identification of retinopathy of prematurity related miRNAs in hyperoxia-induced neonatal rats by deep sequencing. Int. J. Mol. Sci. 2014, 16, 840–856. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.M.; Dong, X.; Tu, Y.; Liu, P. A microarray study of chronic unpredictable mild stress rat blood serum with electro-acupuncture intervention. Neurosci. Lett. 2016, 627, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Yi, J.-S.; Lee, H.-J.; Lee, I.-W.; Park, K.-C.; Yang, J.-H. Dysregulated expression profiles of microRNAs of experimentally induced cerebral aneurysms in rats. J. Korean Neurosurg. Soc. 2013, 53, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Robinson, A.M.; Zucchi, F.C.R.; Robbins, J.C.; Babenko, O.; Kovalchuk, O.; Kovalchuk, I.; Olson, D.M.; Metz, G.A.S. Ancestral exposure to stress epigenetically programs preterm birth risk and adverse maternal and newborn outcomes. BMC Med. 2014, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Rau, T. Detection of Traumatic Brain Injury. U.S. Patent No 9,605,315, 28 March 2017. [Google Scholar]
- Herman, A.; Gruden, K.; Blejec, A.; Podpečan, V.; Motaln, H.; Rožman, P.; Hren, M.; Zupančič, K.; Veber, M.; Verbovšek, U. Analysis of glioblastoma patients’ plasma revealed the presence of microRNAs with a prognostic impact on survival and those of viral origin. PLoS ONE 2015, 10, e0125791. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, M.; He, H.; Chen, J.; Zeng, H.; Li, J.; Duan, R. MicroRNA/mRNA profiling and regulatory network of intracranial aneurysm. BMC Med. Genom. 2013, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Camps, C.; Saini, H.K.; Mole, D.R.; Choudhry, H.; Reczko, M.; Guerra-Assunção, J.A.; Tian, Y.-M.; Buffa, F.M.; Harris, A.L.; Hatzigeorgiou, A.G. Integrated analysis of microRNA and mRNA expression and association with HIF binding reveals the complexity of microRNA expression regulation under hypoxia. Mol. Cancer 2014, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Bai, S.; Su, L.; Li, C.; Wu, J.; Xia, Z.; Xu, D. A subset of circulating microRNAs is expressed differently in patients with myocardial infarction. Mol. Med. Rep. 2015, 12, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-J.; Lin, Y.; Jin, Y.; Zheng, C.-Q. Investigation of key microRNAs associated with hepatocellular carcinoma using small RNA-seq data. Mol. Biol. Rep. 2014, 41, 4341–4349. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shen, Z.-L.; Wang, L.; Lv, C.-Y.; Huang, X.-E.; Zhou, R.-P. Hsa-miR-181a-5p expression and effects on cell proliferation in gastric cancer. Asian Pac. J. Cancer Prev. 2013, 14, 3871–3875. [Google Scholar] [CrossRef] [PubMed]
- Van der Goten, J.; Vanhove, W.; Lemaire, K.; Van Lommel, L.; Machiels, K.; Wollants, W.-J.; De Preter, V.; De Hertogh, G.; Ferrante, M.; Van Assche, G. Integrated miRNA and mRNA expression profiling in inflamed colon of patients with ulcerative colitis. PLoS ONE 2014, 9, e116117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.; Li, Q.; Liu, S.; Wang, F.; Xiong, Z.; Chen, J.; Chen, H.; Yang, Y.; Tan, X.; Luo, Q. Integrated microRNA and mRNA transcriptome sequencing reveals the potential roles of miRNAs in stage I endometrioid endometrial carcinoma. PLoS ONE 2014, 9, e110163. [Google Scholar] [CrossRef] [PubMed]
- Aghanoori, M.-R.; Mirzaei, B.; Tavallaei, M. MiRNA molecular profiles in human medical conditions: Connecting lung cancer and lung development phenomena. Asian Pac. J. Cancer Prev. 2014, 15, 9557–9565. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.M.; Orosco, R.K.; Shen, J.P.; Egloff, A.M.; Carter, H.; Hofree, M.; Choueiri, M.; Coffey, C.S.; Lippman, S.M.; Hayes, D.N. Multi-tiered genomic analysis of head and neck cancer ties TP53 mutation to 3p loss. Nat. Genet. 2014, 46, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-Z.; Tian, Y.; Ander, B.P.; Xu, H.; Stamova, B.S.; Zhan, X.; Turner, R.J.; Jickling, G.; Sharp, F.R. Brain and Blood microRNA Expression Profiling of Ischemic Stroke, Intracerebral Hemorrhage, and Kainate Seizures. J. Cereb. Blood Flow Metab. 2010, 30, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Inaba, T.; Nito, C.; Kamiya, N.; Katayama, Y. Therapeutic impact of eicosapentaenoic acid on ischemic brain damage following transient focal cerebral ischemia in rats. Brain Res. 2013, 1519, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Memezawa, H.; Smith, M.L.; Siesjö, B.K. Penumbral tissues salvaged by reperfusion following middle cerebral artery occlusion in rats. Stroke 1992, 23, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Bederson, J.B.; Pitts, L.H.; Germano, S.M.; Nishimura, M.C.; Davis, R.L.; Bartkowski, H.M. Evaluation of 2,3,5-triphenyltetrazolium chloride as a stain for detection and quantification of experimental cerebral infarction in rats. Stroke 1986, 17, 1304–1308. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.A.; Morton, M.T.; Tsao-Wu, G.; Savalos, R.A.; Davidson, C.; Sharp, F.R. A semiautomated method for measuring brain infarct volume. J. Cereb. Blood Flow Metab. 1990, 10, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Graham, S.H.; Zhu, R.L.; Simon, R.P. Stress proteins and tolerance to focal cerebral ischemia. J. Cereb. Blood Flow Metab. 1996, 16, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.N.; He, Y.Y.; Wu, G.; Khan, M.; Hsu, C.Y. Effect of brain edema on infarct volume in a focal cerebral ischemia model in rats. Stroke 1993, 24, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2014, 43, D146–D152. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Total Volume (mm3) | Nonedema Volume (mm3) | Edema Volume (mm3) | Infarct Volume (mm3) | Edema Index (%) | Infarct Index (%) | |
|---|---|---|---|---|---|---|
| MCAO | ||||||
| M1 | 1326.83 | 1177.82 | 149.00 | 215.50 | 25.30 | 18.30 |
| M2 | 1394.82 | 1227.18 | 167.63 | 255.50 | 27.32 | 20.82 |
| M3 | 1503.44 | 1281.54 | 221.90 | 226.85 | 34.63 | 17.70 |
| Mean | 1408.36 | 1228.85 | 179.51 | 232.62 | 29.08 | 18.94 |
| Sham | ||||||
| S1 | 1319.66 | 1254.14 | 65.51 | 0.00 | 10.45 | 0.00 |
| S2 | 1324.36 | 1289.78 | 34.58 | 0.00 | 5.36 | 0.00 |
| S3 | 1275.43 | 1243.52 | 31.91 | 0.00 | 5.13 | 0.00 |
| Mean | 1306.48 | 1262.48 | 44.00 | 0.00 | 6.98 | 0.00 |
| Ingenuity Canonical Pathways | −log(p-Value) | Z-Score | Molecules |
|---|---|---|---|
| PI3K signaling in B lymphocytes | 2.00 | 2.00 | Cr2, Chp1, Prkcb, Ptprc, Akt2 |
| CD28 signaling in T helper cells | 4.41 | −2.00 | Arpc2, Cd3g, Rt1-Ba, Rt1-Db1, Chp1, Ptprc, Akt2, Rt1-Da |
| Role of NFAT in regulation of the immune response | 3.20 | −2.00 | Cd3g, Rt1-Ba, Rt1-Db1, Gna12, Chp1, Gng12, Akt2, Rt1-Da |
| eNOS signaling | 2.18 | −2.00 | Hspa1a, Hspa8, Prkcb, Hsp90ab1, Hsp90aa1, Akt2 |
| Sphingosine-1-phosphate signaling | 2.17 | −2.00 | Rhoh, Gna12, Acer2, Akt2, S1pr1 |
| fMLP signaling in neutrophils | 2.07 | −2.00 | Arpc2, Chp1, Gng12, Prkcb, Cybb |
| CXCR4 signaling | 2.04 | −2.00 | Rhph, Gna12, Myl12b, Gng12, Prkcb, Akt2 |
| G beta gamma signaling | 1.64 | −2.00 | Gna12, Gng12, Prkcb, Akt2 |
| Fcγ receptor-mediated phagocytosis in macrophages and monocytes | 1.57 | −2.00 | Arpc2, Vamp3, Prkcb, Akt2 |
| Gαq signaling | 1.51 | −2.00 | Rhoh, Chp1, Gng12, Prkcb, Akt2 |
| Signaling by rho family GTPases | 1.32 | −2.00 | Arpc2, Rhoh, Gna12, Myl12b, Gng12, Cybb |
| Integrin signaling | 2.01 | −2.24 | Arpc2, Rhoh, Itgal, Pfn1, Myl12b, Ppp1cb, Akt2 |
| IL-8 signaling | 2.88 | −2.65 | Rhoh, Irak1, Cr2, Gna12, Gng12, Prkcb, Cybb, Akt2 |
| Gene Symbol | Gene Name | Probe Set ID | FDR (M < S) |
|---|---|---|---|
| Cd36 | CD36 molecule (thrombospondin receptor) | 1367689_a_at | 0.0212 |
| 1386901_at | 0.0357 | ||
| Cd3g | CD3 molecule, gamma | 1384787_at | 0.0466 |
| Cd47 | Cd47 molecule | 1369559_a_at | 0.0192 |
| Cd59 | CD59 molecule, complement regulatory protein | 1367929_at | 0.0186 |
| Cd74 | Cd74 molecule, major histocompatibility complex, class II invariant chain | 1367679_at | 0.0003 |
| Signal | ||||||
|---|---|---|---|---|---|---|
| miRNA | MCAO | Sham | Fold Change | p-Value | Target Gene | Target mRNA |
| miR-107-5p | 1.08 | 0.19 | 1.85 | 0.022 | 69 | H2afz (M < S) |
| miR-383-5p | 4.26 | 3.39 | 1.83 | 0.024 | 19 | |
| miR-24-1-5p | 2.58 | 1.91 | 1.59 | 0.031 | 21 | |
| mir-191b | 1.06 | 0.41 | 1.57 | 0.004 | ||
| miR-196b-5p | 1.51 | 0.90 | 1.53 | 0.050 | 22 | Ptprc (M < S) |
| miR-3552 | 0.98 | 0.37 | 1.52 | 0.002 | 74 | Srsf2 (M < S) |
| mir-194-1 | 0.20 | 0.99 | −1.72 | 0.017 | ||
| Rat miRNA | Sequences | Human miRNA | Sequences | Mismatch |
|---|---|---|---|---|
| miR-107-5p | 24, 1–23 | hsa-miR-103a-2-5p | 23, 1–23 | 1 |
| rno-miR-383-5p | 21, 2–21 | hsa-miR-383-5p | 22, 1–20 | 1 |
| rno-miR-24-1-5p | 22, 2–22 | hsa-miR-24-1-5p | 22, 1–21 | 0 |
| 22, 2–18 | hsa-miR-24-2-5p | 22, 1–17 | 1 | |
| rno-mir-191b | 120, 15–104 | hsa-mir-191 | 92, 2–91 | 30 |
| 120, 22–84 | hsa-mir-4653 | 83, 15–77 | 25 | |
| rno-miR-196b-5p | 22, 1–22 | hsa-miR-196b-5p | 22, 1–22 | 0 |
| 22, 1–22 | hsa-miR-196a-5p | 22, 1–22 | 1 | |
| rno-miR-3552 | 21 | |||
| rno-mir-194-1 | 83, 1–83 | hsa-mir-194-1 | 85, 1–84 | 12 |
| 83, 11–67 | hsa-mir-194-2 | 85, 11–68 | 10 | |
| 83, 60–78 | hsa-mir-548az | 95, 39–57 | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takuma, A.; Abe, A.; Saito, Y.; Nito, C.; Ueda, M.; Ishimaru, Y.; Harada, H.; Abe, K.; Kimura, K.; Asakura, T. Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood. Int. J. Mol. Sci. 2017, 18, 2335. https://doi.org/10.3390/ijms18112335
Takuma A, Abe A, Saito Y, Nito C, Ueda M, Ishimaru Y, Harada H, Abe K, Kimura K, Asakura T. Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood. International Journal of Molecular Sciences. 2017; 18(11):2335. https://doi.org/10.3390/ijms18112335
Chicago/Turabian StyleTakuma, Ayako, Arata Abe, Yoshikazu Saito, Chikako Nito, Masayuki Ueda, Yoshiro Ishimaru, Hideki Harada, Keiko Abe, Kazumi Kimura, and Tomiko Asakura. 2017. "Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood" International Journal of Molecular Sciences 18, no. 11: 2335. https://doi.org/10.3390/ijms18112335
APA StyleTakuma, A., Abe, A., Saito, Y., Nito, C., Ueda, M., Ishimaru, Y., Harada, H., Abe, K., Kimura, K., & Asakura, T. (2017). Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood. International Journal of Molecular Sciences, 18(11), 2335. https://doi.org/10.3390/ijms18112335