
Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice

Abstract
:
1. Introduction
2. Results
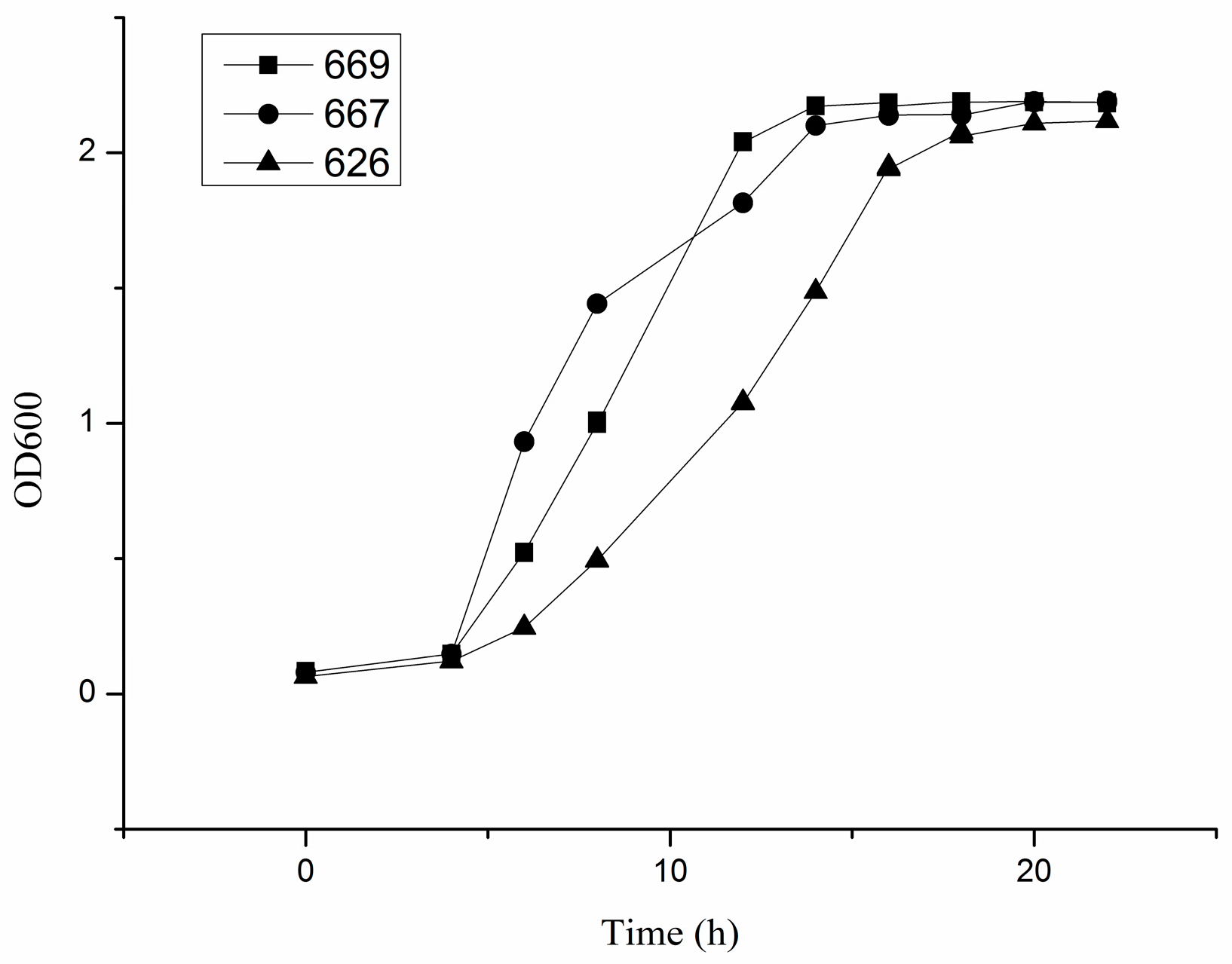
2.1. Growth Characteristics of B. adolescentis In Vitro
2.2. Tolerance Capacity of B. adolescentis to Simulated Gastric and Small Intestine Juices
2.3. Adhesion of Different B. adolescentis to HT-29 Cells
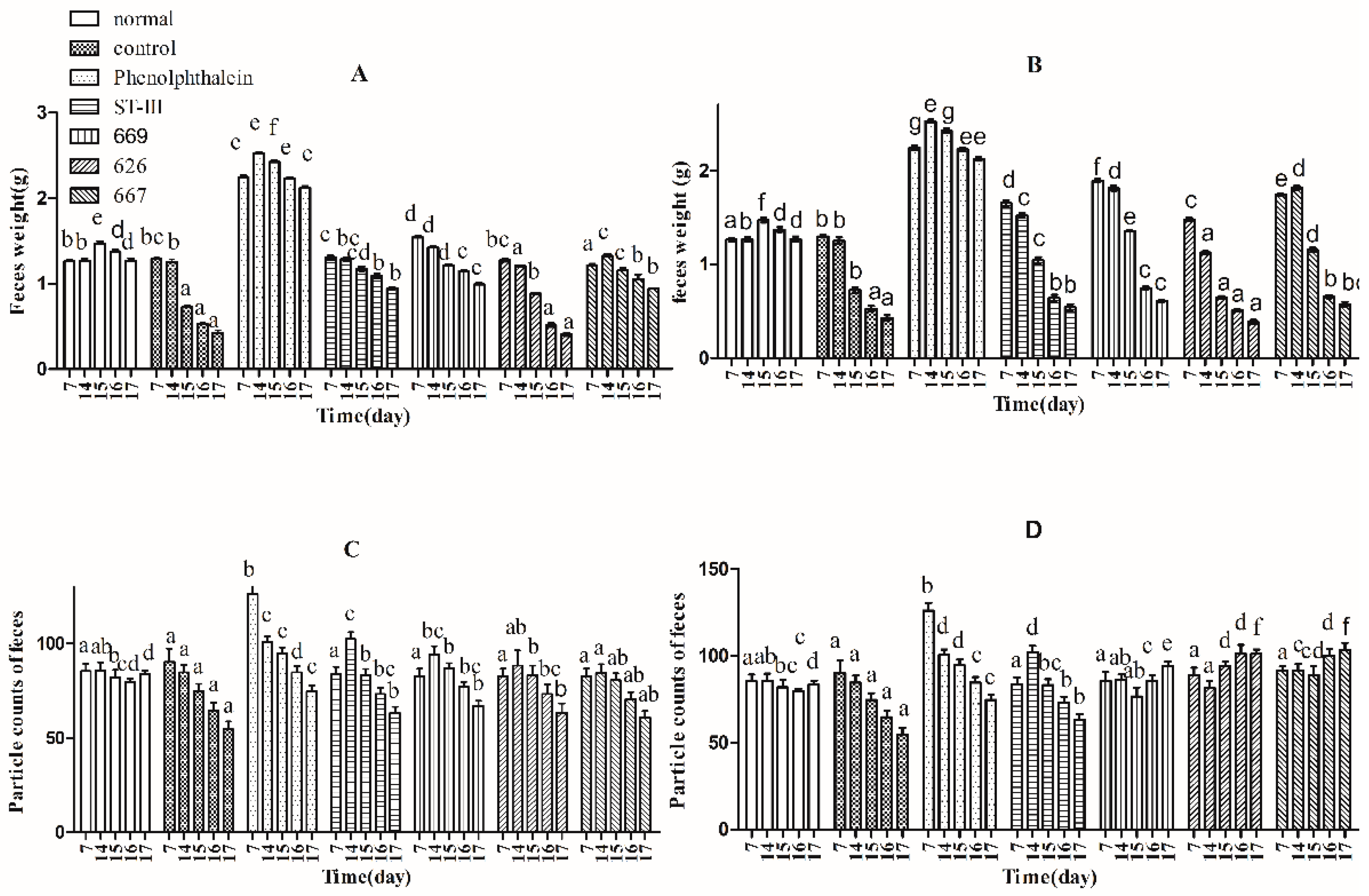
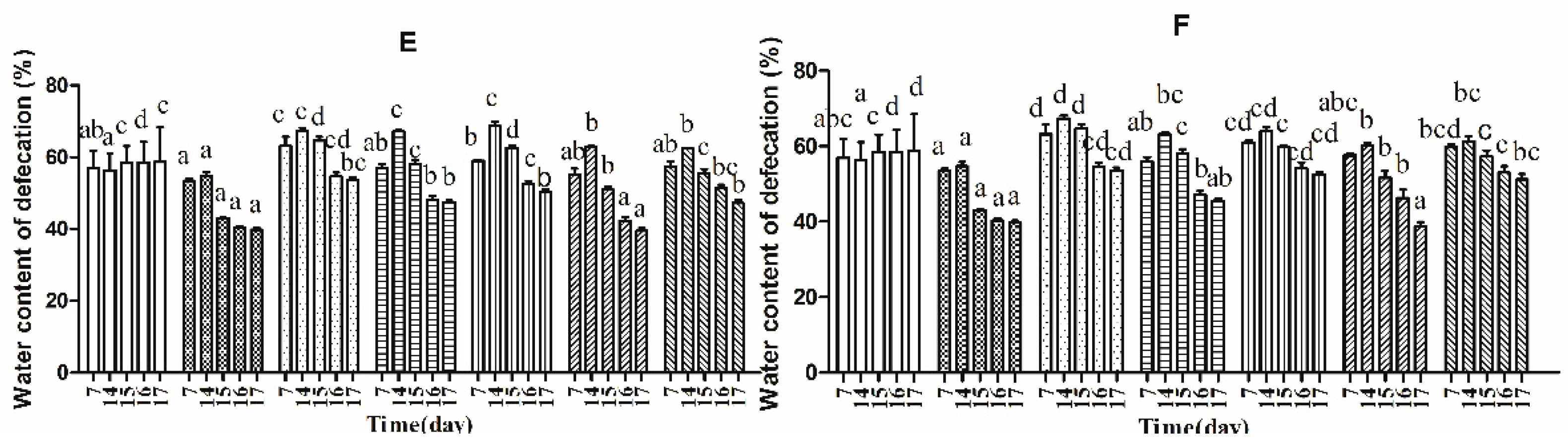
2.4. Effects of B. adolescentis on Defecation Status of Mice
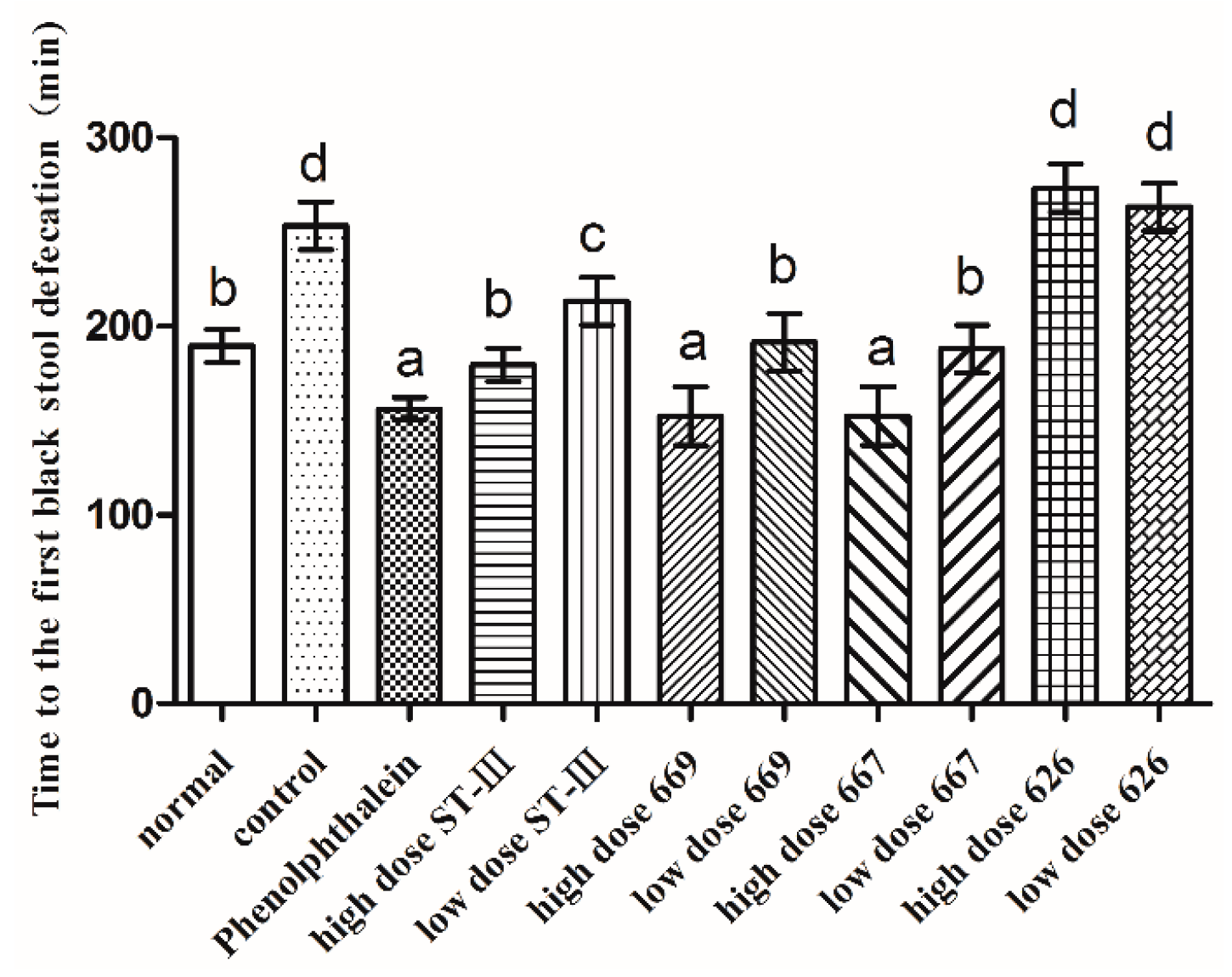
2.5. Time to the First Black Stool Defecation
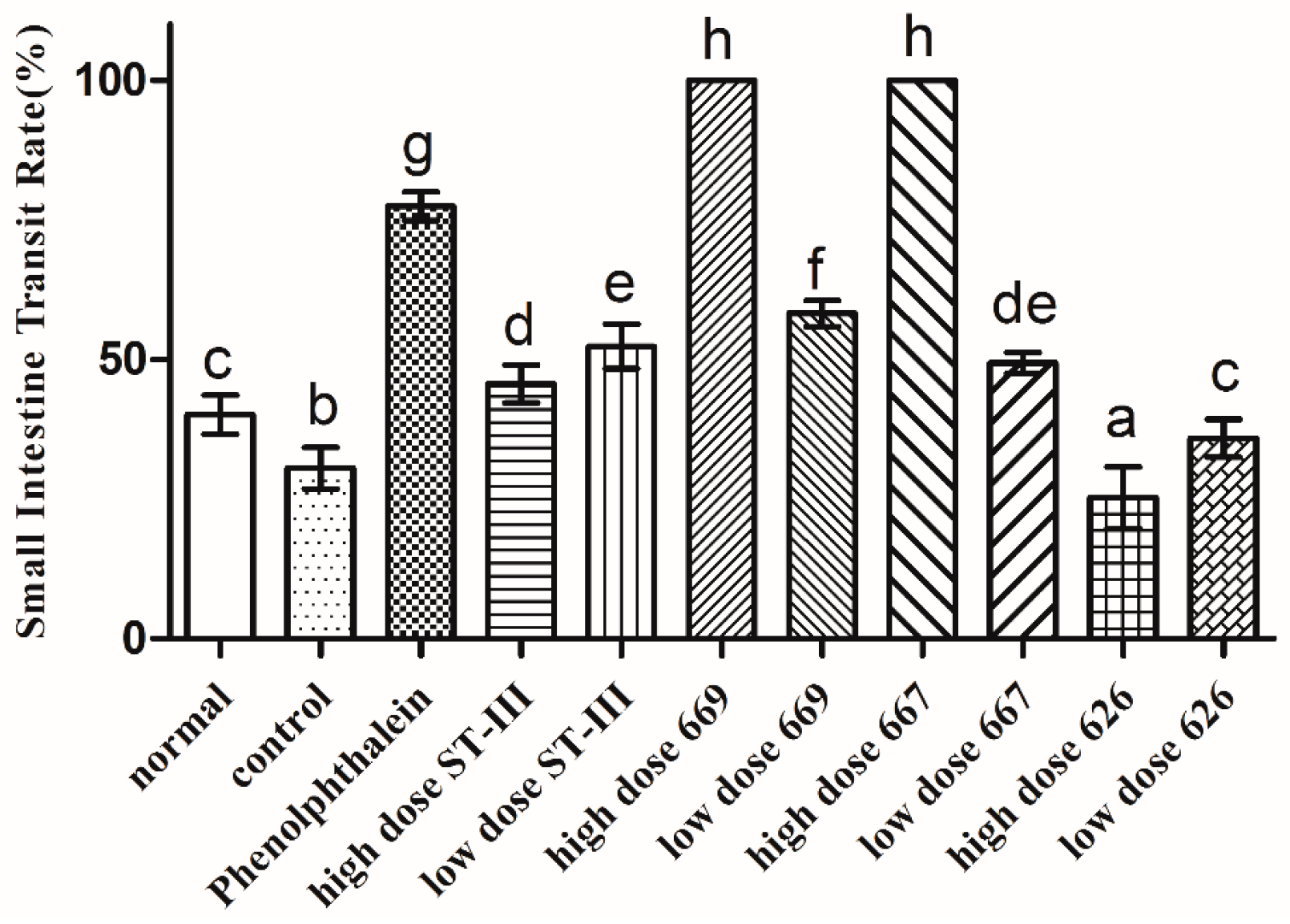
2.6. Gastrointestinal (GI) Transit Rate
2.7. Short-Chain Fatty Acids (SCFAs) in Feces
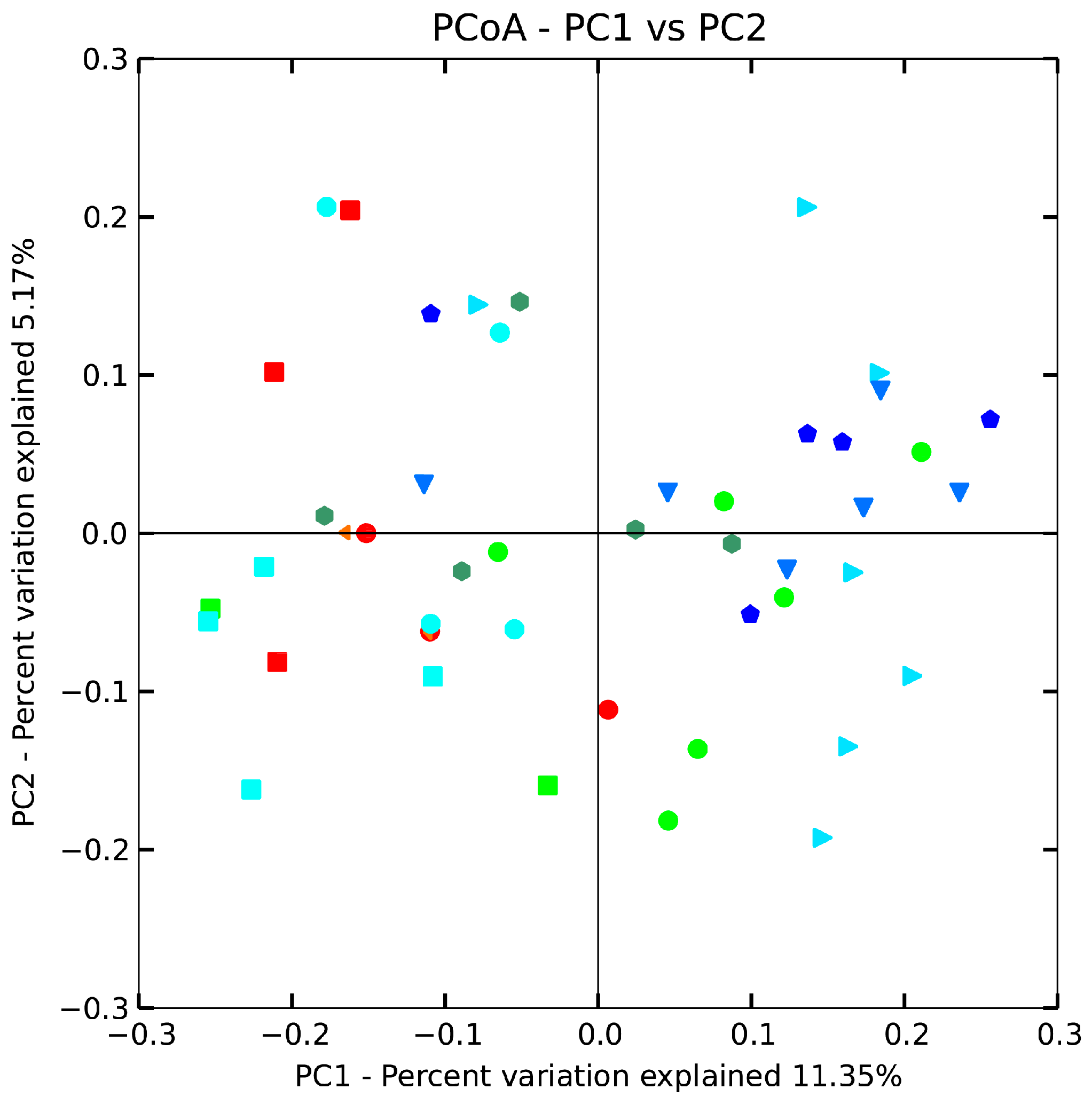
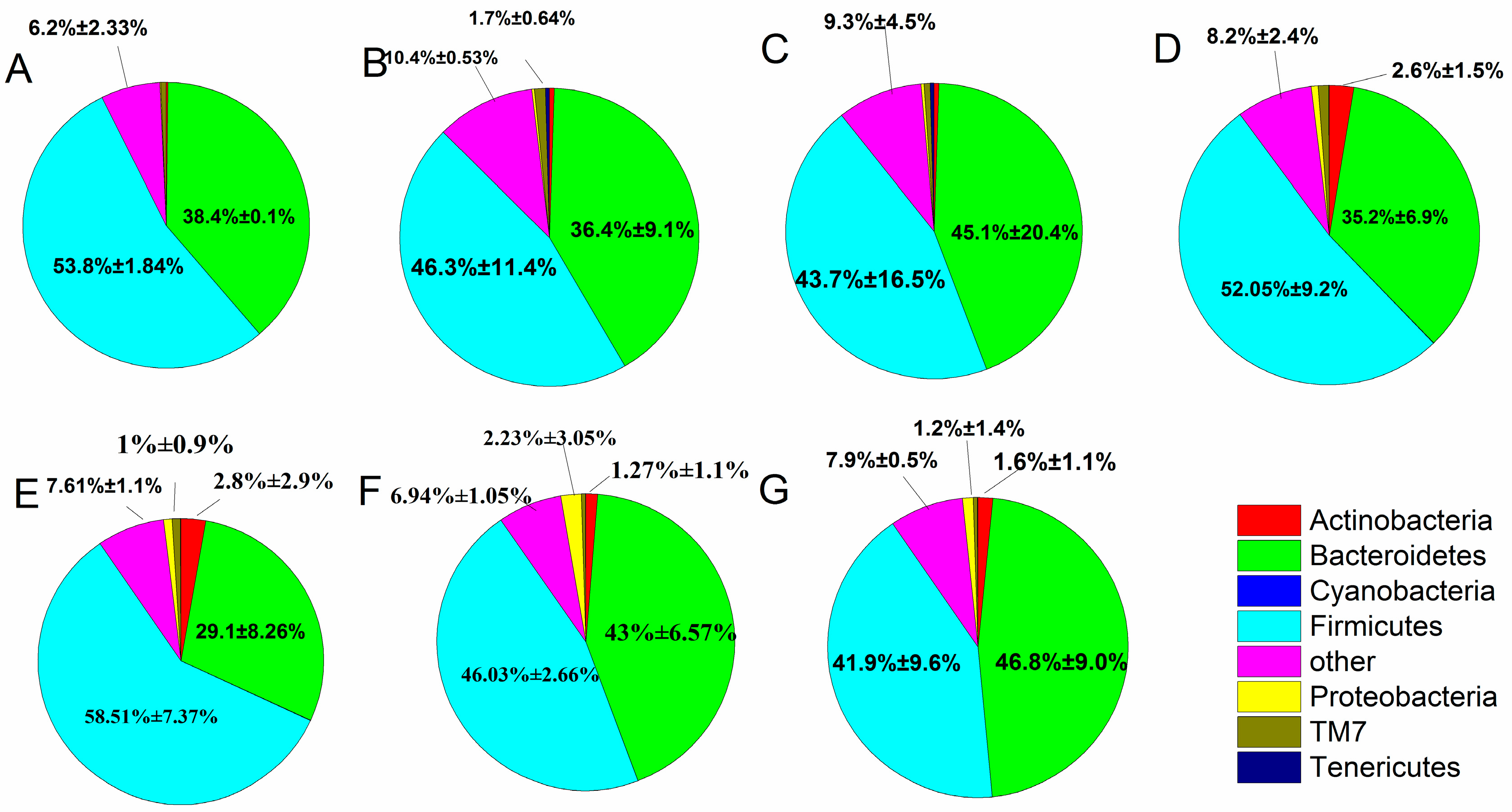
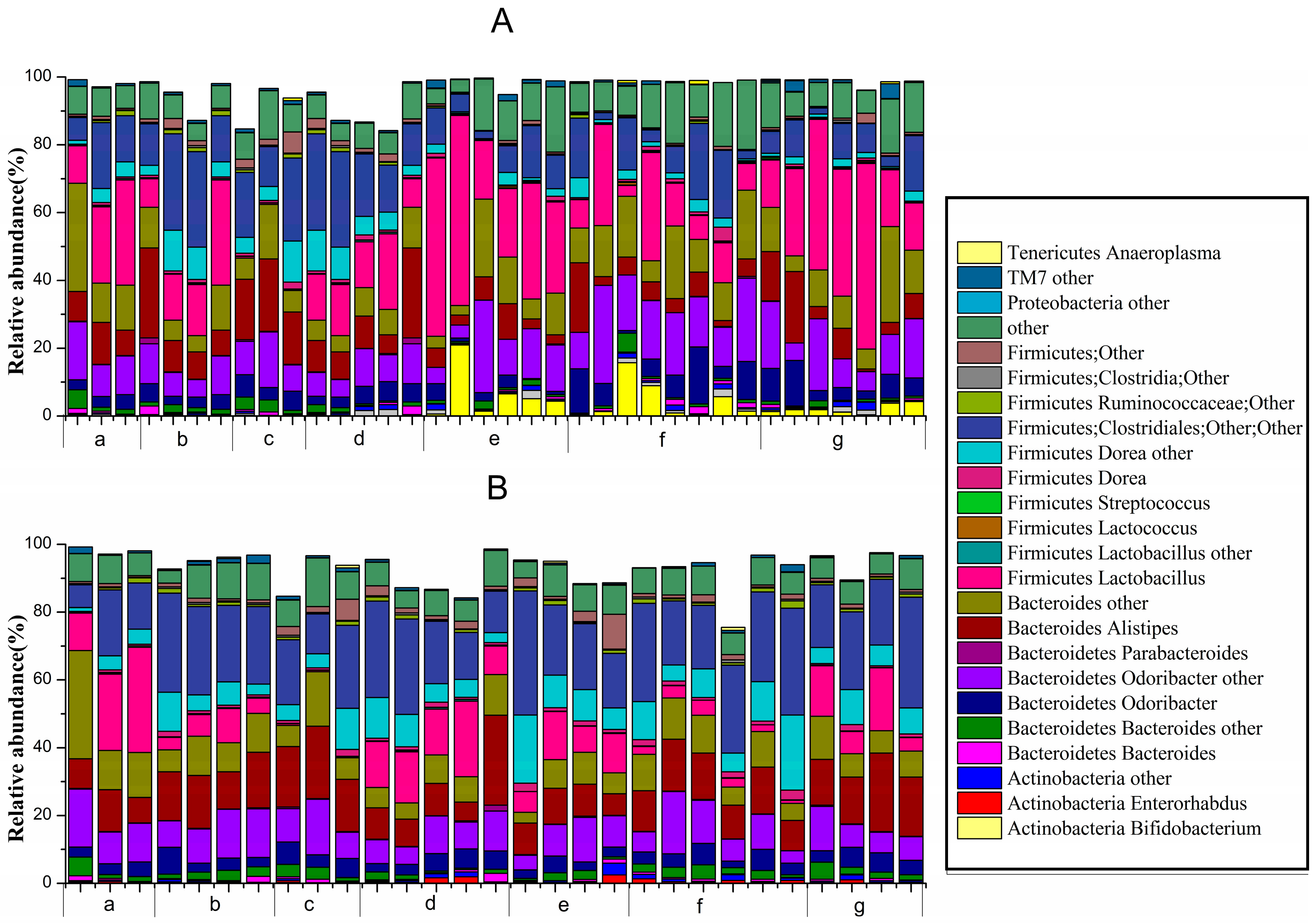
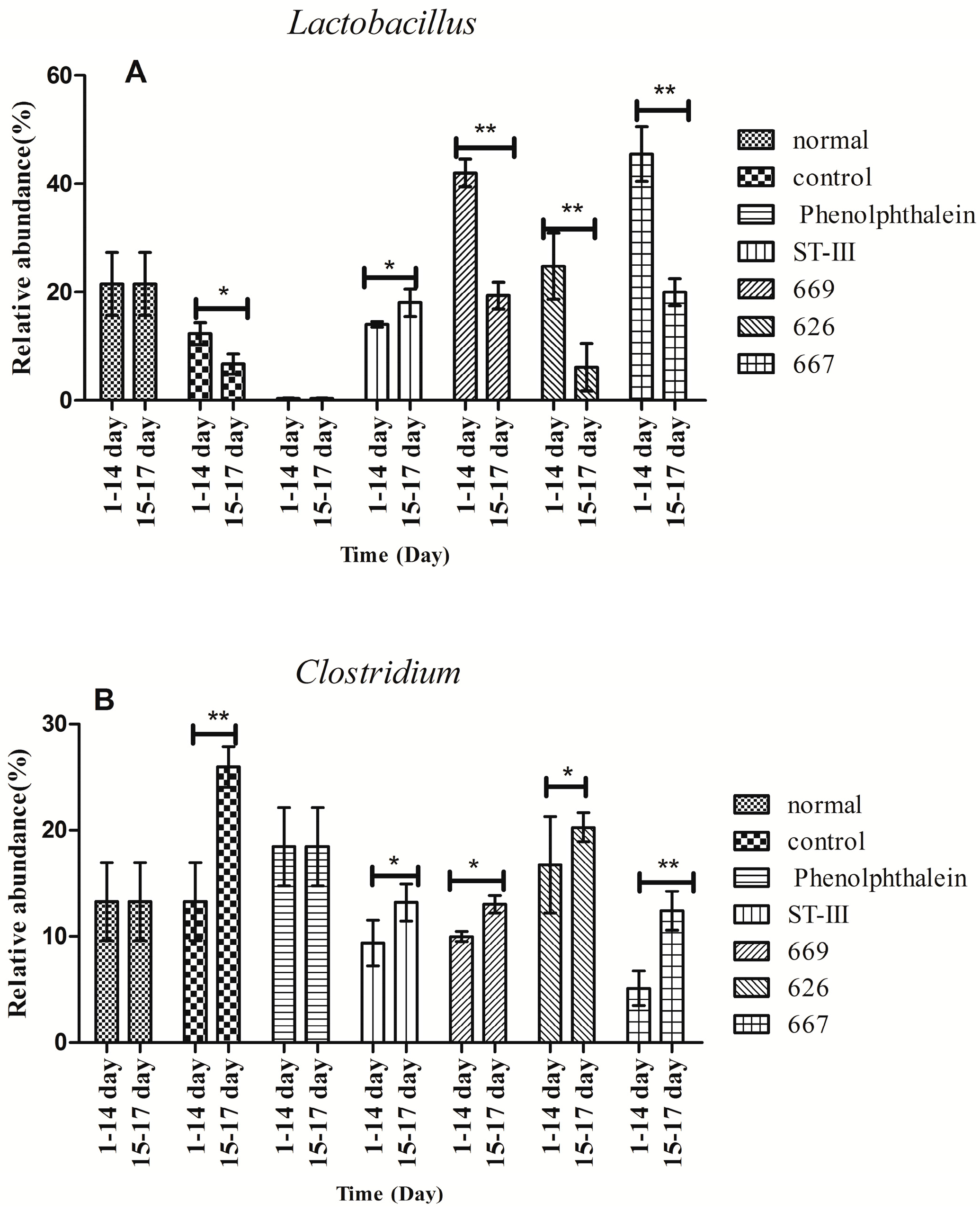
2.8. Effect of B. adolescentis on Composition of Fecal Microbiota
2.9. Characterization of Serum Parameters of Experimental Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Bacterial Strain and Culture Conditions
4.3. Growth Curve of B. adolescentis
4.4. Tolerance Capacity of B. adolescentis to Simulated Gastric and Small Intestine Juices
4.5. Determination of Adhesion Properties of Bifidobacterium spp. to HT-29 Cells In Vitro
4.6. Animal Experiments
4.6.1. Animals
4.6.2. Induction of Constipation and Experimental Design
4.6.3. Measurement of Defecation Status of Mice
4.6.4. Determination of the Time to the First Black Stool Defecation
4.6.5. Determination of GI Transit
4.6.6. Determination of SCFAs in Feces
4.6.7. MiSeq Genome Sequencing Analysis of Community Structures
4.6.8. Determination of MTL, Gas, ET, SS, SP and VIP Levels in Serum
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stern, T.; Davis, A.M. Evaluation and treatment of constipation. JAMA Clin. Guidel. Synop. 2016, 315, 192–193. [Google Scholar]
- McQuaid, K.R.; Katzung, B.G.; Masters, S.B. Drugs Used in the Treatment of Gastrointestinal Diseases. In Basic & Clinical Pharmacology, 12th ed.; Trevor, A.J., Ed.; McGraw-Hill: New York, NY, USA, 2012. [Google Scholar]
- Xu, J.; Zhou, X.; Chen, C.; Deng, Q.; Huang, Q.; Yang, J.; Yang, N.; Huang, F. Laxative effects of partially defatted flaxseed meal on normal and experimental constipated mice. BMC Complement. 2012, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; di Baise, J.K.; Ilhan, Z.E.; Crowell, M.D.; Rideout, J.R.; Caporaso, J.G.; Rittmann, B.E.; Krajmalnik-Brown, R. Gut microbial and short-chain fatty acid profiles in adults with chronic constipation before and after treatment with lubiprostone. Anaerobe 2015, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, G.; Cinquetti, M.; Luciano, A.; Benini, A.; Muner, A.; Bertazzoni Minelli, E. The intestinal ecosystem in chronic functional constipation. Acta Paediatr. 1998, 87, 836–841. [Google Scholar] [CrossRef] [PubMed]
- De Vrese, M.; Marteau, P.R. Probiotics and Prebiotics: Effects on Diarrhea. J. Nutr. 2007, 137, 803S–811S. [Google Scholar] [PubMed]
- Gopal, P.K.; Prasad, J.; Smart, J.; Gill, H.S. In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int. J. Food Microbiol. 2001, 67, 207–216. [Google Scholar] [CrossRef]
- Quigley, E.M. The enteric microbiota in the pathogenesis and management of constipation. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Maurice, C.F.; Haiser, H.J.; Turnbaugh, P.J. Xenobiotics shape the physiology and gene expression of the active human gut microbiome. Cell 2013, 152, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, G.; Leggio, F.; Vacante, M.; Motta, M.; Giordano, M.; Biondi, A.; Basile, F.; Mastrojeni, S.; Mistretta, A.; Malaguarnera, M.; et al. Probiotics in the gastrointestinal diseases of the elderly. J. Nutr. Health Aging 2012, 16, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Arioli, S.; Wang, A.; Villa, C.R.; Jahani, R.; Song, Y.S.; Mora, D.; Guglielmetti, S.; Comelli, E.M. Impact of Bifidobacterium bifidum MIMBb75 on mouse intestinal microorganisms. FEMS Microbiol. Ecol. 2013, 85, 369–375. [Google Scholar] [CrossRef] [PubMed]
- O’mahony, L.; Mccarthy, J.; Kelly, P.; Hurley, G.; Luo, F.; Chen, K.; O’sullivan, G.C.; Kiely, B.; Collins, J.K.; Shanahan, F.; et al. Lactobacillus and bifidobacterium in irritable bowel syndrome: Symptom responses and relationship to cytokine profiles. Gastroenterology 2005, 38, 1–8. [Google Scholar] [CrossRef]
- Kassinen, A.; Krogius-Kurikka, L.; Mäkivuokko, H.; Rinttilä, T.; Paulin, L.; Corander, J.; Malinen, E.; Apajalahti, J.; Palva, A. The Fecal Microbiota of Irritable Bowel Syndrome Patients Differs Significantly From That of Healthy Subjects. Gastroenterology 2007, 131, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M. Probiotics in the Management of Functional Bowel Disorders Promise Fulfilled? Gastroenterol. Clin. N. Am. 2012, 41, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Urita, Y.; Goto, M.; Watanabe, T.; Matsuzaki, M.; Gomi, A.; Kano, M.; Miyazaki, K.; Kaneko, H. Continuous consumption of fermented milk containing Bifidobacterium bifidum YIT 10347 improves gastrointestinal and psychological symptoms in patients with functional gastrointestinal disorders. Biosci. Microbiota Food Health 2015, 34, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ojetti, V.; Ianiro, G.; Tortora, A.; D’Angelo, G.; di Rienzo, T.A.; Bibbo, S.; Migneco, A.; Gasbarrini, A. The Effect of Lactobacillus reuteri Supplementation in Adults with Chronic Functional Constipation: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Gastrointest. Liver Dis. 2014, 23, 387–391. [Google Scholar]
- Kim, S.; Choi, S.C.; Park, K.S.; Park, M.I.; Shin, J.E.; Lee, T.H.; Jung, K.W.; Koo, H.S.; Myung, S.; Constipation Research Group of Korean Society of Neurogastroenterology and Motili. Change of Fecal Flora and Effectiveness of the Short-term VSL#3 Probiotic Treatment in Patients with Functional Constipation. J. Neurogastroenterol. Motil. 2015, 21, 111–120. [Google Scholar] [PubMed]
- Waitzberg, D.L.; Quilici, F.A.; Michzputen, S.; Passos, M.D.C.F. The effect of probiotic fermented milk that includes Bifidobacterium lactis CNCM I-2494 on the reduction of gastrointestinal discomfort and symptoms in adults: A narrative review. Nutr. Hosp. 2015, 32, 501–509. [Google Scholar] [PubMed]
- Riezzo, G.; Orlando, A.; D’Attoma, B.; Guerra, V.; Valerio, F.; Lavermicocca, P.; de Candia, S.; Russo, F. Randomised clinical trial: Efficacy of Lactobacillus paracasei-enriched artichokes in the treatment of patients with functional constipation—A double-blind, controlled, crossover study. Aliment. Pharmacol. Ther. 2012, 35, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Mazlyn, M.M.; Nagarajah, L.H.L.; Fatimah, A.; Norimah, A.K.; Goh, K.L. Effects of a probiotic fermented milk on functional constipation: A randomized, double-blind, placebo-controlled study. J. Gastroenterol. Hepatol. 2013, 28, 1584. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.S.; Wagner, K. Influence of dietary phytochemicals and microbiota on colon cancer risk. J. Agric. Food Chem. 2012, 60, 6728–6735. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, X.E. Efficacy of Bifidobacterium tetragenous Viable Bacteria Tablets for Cancer Patients with Functional Constipation. Asian Pac. J. Cancer Prev. 2014, 15, 10241–10244. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Cuillerier, E.; Meance, S.; Gerhardt, M.F.; Myara, A.; Bouvier, M.; Bouley, C.; Tondu, F.; Bommelaer, G.; Grimaud, J.C. Bifidobacterium animalis strain DN-173 010 shortens the colonic transit time in healthy women: A double-blind, randomized, controlled study. Aliment. Pharmacol. Ther. 2002, 16, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, T.; Takahashi, R.; Kimura, M.; Fukuda, Y. Study of the mechanisms of a Japanese traditional fermented medicine in the improvement of constipation. J. Gastroenterol. Hepatol. 2015, 30, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Christodoulides, S.; Fragkos, K.C.; Scott, S.M.; Whelan, K. The effect of probiotics on functional constipation in adults: A systematic review and meta-analysis of randomized. Am. J. Clin. Nutr. 2014, 100, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.J.; Cummings, J.H.; Macfarlane, G.T. Inter-species differences in maximum specific growth rates and cell yields of Bifidobacteria cultured on oligosaccharides and other simple carbohydrate sources. J. Appl. Microbiol. 1998, 85, 381–386. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Pemberton, J.H.; Locke, G.R., III. American Gastroenterological Association technical review on constipation. Gastroenterology 2013, 144, 218–238. [Google Scholar] [CrossRef] [PubMed]
- Aichbichler, B.W.; Wenzl, H.H.; Santa Ana, C.A.; Porter, J.L.; Schiller, L.R.; Fordtran, J.S. A comparison of stool characteristics from normal and constipated people. Dig. Dis. Sci. 1998, 43, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Sohji, Y.; Kawashima, K.; Shimizu, M. Pharmacological studies of loperamide, an anti-diarrheal agent II. Effects on peristalsis of the small intestine and colon in guinea pigs. Folia Pharmacol. Jpn. 1978, 74, 155–163. [Google Scholar] [CrossRef]
- Salminen, S.; Bouley, C.; Boutron-Ruault, M.C.; Cummings, J.H.; Franck, A.; Gibson, G.R.; Isolauri, E.; Moreau, M.C.; Roberfroid, M.; Rowland, I. Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 1998, 80, S147–S171. [Google Scholar] [CrossRef] [PubMed]
- Van der Werf, M.J.; Venema, K. Bifidobacteria: Genetic modification and the study of their role in the colon. J. Agric. Food Chem. 2001, 49, 378–383. [Google Scholar] [CrossRef] [PubMed]
- De Milliano, I.; Tabbers, M.M.; van der Post, J.A.; Benninga, M.A. Is a multispecies probiotic mixture effective in constipation during pregnancy? ‘A pilot study’. Nutr. J. 2012, 11, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Prasad, J.; Gill, H.; Stevenson, L.; Gopal, P. Impact of consumption of different levels of Bifidobacterium lactis HN019 on the intestinal microflora of elderly human subjects. J. Nutr. Health Aging 2007, 11, 26–31. [Google Scholar] [PubMed]
- Waller, P.A.; Gopal, P.K.; Leyer, G.J.; Ouwehand, A.C.; Reifer, C.; Stewart, M.E.; Miller, L.E. Dose-response effect of Bifidobacterium lactis HN019 on whole gut transit time and functional gastrointestinal symptoms in adults. Scand. J. Gastroenterol. 2011, 46, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Favretto, D.C.; Pontin, B.; Moreira, T.R. Effect of the consumption of a cheese enriched with probiotic organisms (Bifidobacterium lactis bi-07) in improving symptoms of constipation. Arq. Gastroenterol. 2013, 50, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Vaishnavi, C. Clinical spectrum & pathogenesis of Clostridium difficile associated diseases. Indian J. Med. Res. 2010, 131, 487–499. [Google Scholar] [PubMed]
- Kleessen, B.; Sykura, B.; Zunft, H.-J.; Blaut, M. Effects of inulin and lactose on fecal microflora, microbial activity, and bowel habit in elderly constipated persons. Am. J. Clin. Nutr. 1997, 65, 1397–1402. [Google Scholar] [PubMed]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Khalif, I.L.; Quigley, E.M.M.; Konovitch, E.A.; Maximova, I.D. Alterations in the colonic flora and intestinal permeability and evidence of immune activation in chronic constipation. Dig. Liver Dis. 2005, 37, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Ritzhaupt, A.; Wood, I.S.; Ellis, A.; Hosie, K.B.; Shirazi-Beechey, S.P. Identification and characterization of a monocarboxylate transporter (MCT1) in pig and human colon: Its potential to transport l-lactate as well as butyrate. J. Physiol. 1998, 513, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Short chain fatty acids and intestinal microflora. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Satapati, S.; Kucejova, B.; Duarte, J.A.; Fletcher, J.A.; Reynolds, L.; Sunny, N.E.; He, T.; Nair, L.A.; Livingston, K.; Fu, X.; et al. Mitochondrial metabolism mediates oxidative stress and inflammation in fatty liver. J. Clin. Investig. 2015, 125, 4447–4462. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.C.; Kien, C.L.; Bouthillier, L.; Levy, E. Short-chain fatty acids: Ready for prime time? Nutr. Clin. Pract. 2006, 21, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C.; Dapoigny, M.; Scott, K.P.; Crouzet, L.; Del’homme, C.; Marquet, P.; Martin, J.C.; Pickering, G.; Ardid, D.; Eschalier, A.; et al. Functional dysbiosis within the gut microbiota of patients with constipated-irritable bowel syndrome. Aliment. Pharmacol. Ther. 2012, 35, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a three-stage compound continuous culture system for investigating the effect of retention time on the ecology and metabolism of bacteria in the human colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Mariadason, J.M.; Corner, G.A.; Augenlicht, L.H. Genetic reprogramming of colonic cell maturation induced by short chain fatty acids: Comparison with trichostatin A, sulindac, and curcumin and implications for chemoprevention of colon cancer. Cancer Res. 2000, 60, 4561–4572. [Google Scholar] [PubMed]
- Menzel, T.; Schauber, J.; Kreth, F.; Kudlich, T.; Melcher, R.; Gostner, A.; Scheppach, W.; Lührs, H. Butyrate and aspirin in combination have an enhanced effect on proliferation and apoptosis in human colorectal cancer cells. Eur. J. Cancer Prev. 2002, 11, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S.; Rocken, C.; Mawrin, C.; Weise, W.; Hollt, V.; Schulz, S. Immunocyto chemical identification of VPAC1, VPAC2, and PAC1 receptors in normal and neoplastic human tissues with subtype-specific antibodies. Clin. Cancer Res. 2004, 10, 8235–8242. [Google Scholar] [CrossRef] [PubMed]
- Peracchi, M.; Basilisco, G.; Tagliabue, R.; Terrani, C.; Locati, A.; Bianchi, P.A.; Velio, P. Postprandial gut peptide plasma levels in women with idiopathic slow transit constipation. Scand. J. Gastroenterol. 1999, 34, 25–28. [Google Scholar]
- Stern, H.P.; Stroh, S.E.; Fiedorek, S.C.; Kelleher, K.; Mellon, M.W.; Pope, S.K.; Rayford, P.L. Increased plasma levels of pancreatic polypeptide and decreased plasms levels of motilin in encopretic children. Rayford PL Pediatr. 1995, 96, 111–117. [Google Scholar]
- EL-Salhy, M.; Norrgard, O. Colonic neuroendocrine peptide levels in patients with chronic idiopathic slow transit constipation. Ups. J. Med. Sci. 1998, 103, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Colucci, W.S. Myocardial Endothelin Does It Play a Role in Myocardial Failure? Circulation 1996, 93, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, Q.; Huang, Y.; Ni, L.; Liu, J.; Jiang, J.; Li, N. Function and clinical implications of short-chain fatty acids in patients with mixed refractory constipation. Colorectal Dis. 2016, 18, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Fasth, S. Neuropeptides in idiopathic chronic constipation (slow transit constipation). Neurogastroenterol. Motil. 1995, 9, 143–150. [Google Scholar]
- Beerens, H. An elective and selective insulation medium for Bifidobacterium spp. Lett. Appl. Microbiol. 1990, l11, 155–157. [Google Scholar] [CrossRef]
- Bibiloni, R.; Perez, P.F.; de Antoni, G.L. An enzymatic–colourimetric assay for the quantification of Bifidobacterium. J. Food Prot. 2000, 63, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Coconnier, M.; Klaenhammer, T.R.; Kerneis, S.; Bernet, M.; Servin, A.L. Protein-Mediated Adhesion of Lactobacillus acidophilus BG2FO4 on Human Enterocyte and Mucus-Secreting Cell Lines in Culture. Appl. Environ. Microbiol. 1992, 58, 2034–2039. [Google Scholar] [PubMed]
- Lin, Z.; Wu, J.; Xiao, Z. The curative effect of aloe on constipation and its primary mechanism. Guangdong Med. J. 2005, 26, 1336–1338. [Google Scholar]
- Battish, R.; Cao, G.Y.; Lynn, R.B.; Chakder, S.; Rattan, S. Heme oxygenase-2 distribution in anorectum: Colocalization with neuronal nitric oxide synthase. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, 148–155. [Google Scholar]
- Qian, Y.; Zhao, X.; Kan, J. Preventive effect of resistant starch on activated carbon-induced constipation in mice. Exp. Ther. Med. 2013, 6, 228–232. [Google Scholar] [PubMed]
- Lee, H.Y.; Kim, J.H.; Jeung, H.W.; Lee, C.U.; Kim, D.S.; Li, B.; Lee, G.H.; Sung, M.S.; Ha, K.C.; Back, H.I.; et al. Effects of Ficuscarica paste on loperamide-induced constipation in rats. Food Chem. Toxicol. 2012, 50, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, Y.; Naitoh, Y.; Kamato, T.; Amano, M.; Miyata, K. Compounds possessing 5-HT3 receptor antagonistic activity inhibit intestinal propulsion in mice. Eur. J. Pharmacol. 1996, 311, 67–72. [Google Scholar] [CrossRef]
- Mao, B.; Li, D.; Ai, C.; Zhao, J.; Zhang, H.; Chen, W. Lactulose Differently Modulates the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice. J. Agric. Food Chem. 2016, 64, 6240–6247. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative genetic background of the host influences gut microbiomes in chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]





 , control;
, control;  , normal;
, normal;  , phenolphthalein;
, phenolphthalein;  , ST-III;
, ST-III;  , high-dose 667;
, high-dose 667;  , low-dose 667;
, low-dose 667;  , high-dose 669;
, high-dose 669;  , low-dose 669;
, low-dose 669;  , high-dose 626;
, high-dose 626;  , low-dose 626.
, control; , normal; , phenolphthalein; , ST-III; , high-dose 667; , low-dose 667; , high-dose 669; , low-dose 669; , high-dose 626; , low-dose 626.
, low-dose 626.
, control; , normal; , phenolphthalein; , ST-III; , high-dose 667; , low-dose 667; , high-dose 669; , low-dose 669; , high-dose 626; , low-dose 626.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Time (h) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 2 | 4 | 8 | |
| Survival Rate (%) | ||||||
| Simulated Gastric Juice | Simulated Intestinal Juice | |||||
| B. adolescentis CCFM 626 | 99.30 ± 0.2 | 98.36 ± 0.19 | 85.50 ± 0.20 b | 81.57 ± 0.25 | 79.11 ± 0.41 | 71.51 ± 0.23 a |
| B. adolescentis CCFM 669 | 98.16 ± 0.27 | 96.26 ± 0.31 | 81.43 ± 0.27 a | 78.59 ± 0.33 | 74.55 ± 0.26 | 74.55 ± 0.22 b |
| B. adolescentis CCFM 667 | 99.91 ± 0.07 | 99.34 ± 0.24 | 94.93 ± 0.42 d | 90.17 ± 0.21 | 81.90 ± 0.19 | 79.50 ± 0.30 c |
| L. plantarum ST-III | 92.86 ± 0.28 | 88.28 ± 0.21 | 86.11 ± 0.31 c | 85.64 ± 0.18 | 81.90 ± 0.28 | 80.07 ± 0.28 d |
| Strain | Adhesion |
|---|---|
| B. adolescentis CCFM 667 | – |
| B. adolescentis CCFM 626 | – |
| B. adolescentis CCFM 669 | ++ |
| Lactobacillus plantarum ST-III | +++ |
| Treatment | Time (Day) | Acetic Acid (µmol/g) | Propanoic Acid (µmol/g) | Butyric Acid (µmol/g) | Total Acid (µmol/g) |
|---|---|---|---|---|---|
| Normal | 7 | 13.71 ± 1.33 b | 3.25 ± 0.14 d | 5.00 ± 0.11 e | 21.96 ± 1.57 c |
| 14 | 13.84 ± 1.15 a | 3.38 ± 0.16 d | 4.69 ± 0.66 e | 21.91 ± 1.71 b | |
| 17 | 13.59 ± 0.69 b | 3.07 ± 0.47 c,d | 5.57 ± 0.47 f | 22.23 ± 1.09 b | |
| Control | 7 | 13.32 ± 2.19 b | 2.98 ± 0.3 d | 4.42 ± 0.36 d | 20.72 ± 2.11 b |
| 14 | 13.57 ± 0.43 a | 3.28 ± 0.3 d | 4.92 ± 0.23 f | 21.77 ± 0.80 b | |
| 17 | 8.90 ± 0.51 a | 1.45 ± 0.27 a | 1.67 ± 0.32 a | 12.02 ± 0.94 a | |
| Phenolphthalein | 7 | 7.62 ± 1.45 a | 1.49 ± 0.26 a | 2.44 ± 0.27 a | 11.55 ± 1.52 a |
| 14 | 13.87 ± 1.96 a | 1.48 ± 0.26 a | 2.44 ± 0.27 a | 17.80 ± 1.89 a | |
| 17 | 12.66 ± 0.87 b | 1.53 ± 0.29 a | 2.98 ± 0.05 b | 17.17 ± 1.12 a | |
| ST-III | 7 | 19.63 ± 1.35 d,e | 3.79 ± 0.29 e | 4.90 ± 0.06 e | 28.32 ± 1.22 f |
| 14 | 21.60 ± 0.62 b | 2.70 ± 0.24 c | 2.94 ± 0.34 b | 27.24 ± 0.65 c | |
| 17 | 25.28 ± 2.70 c | 2.56 ± 0.78 b,c | 3.34 ± 1.15 c | 31.19 ± 4.33 c | |
| High dose of 669 | 7 | 24.62 ± 0.67 f | 2.38 ± 0.25 c | 3.62 ± 0.16 c | 30.62 ± 0.99 g |
| 14 | 35.90 ± 2.29 d | 3.37 ± 0.28 d | 4.46 ± 0.36 e | 43.73 ± 2.76 f | |
| 17 | 44.19 ± 3.37 e,f | 3.46 ± 0.12 d | 5.40 ± 0.52 e | 53.05 ± 3.96 e | |
| Low dose of 669 | 7 | 20.13 ± 0.72 e | 1.76 ± 0.11 a,b | 2.46 ± 0.07 a | 24.35 ± 0.75 d |
| 14 | 27.06 ± 1.12 c | 2.52 ± 0.12 b,c | 3.40 ± 0.05 c | 32.98 ± 1.07 e | |
| 17 | 47.29 ± 0.8 f | 3.59 ± 0.59 d | 3.57 ± 0.53 c,d | 54.45 ± 1.91 e | |
| High dose of 626 | 7 | 16.30 ± 0.32 c | 2.42 ± 0.23 c | 4.76 ± 0.24 e | 23.48 ± 0.28 d |
| 14 | 22.97 ± 1.03 b | 2.24 ± 0.26 b | 4.24 ± 0.35 e | 29.45 ± 0.69 d | |
| 17 | 37.64 ± 1.34 d | 2.02 ± 0.63 a,b | 7.66 ± 2.05 g | 47.32 ± 3.94 d | |
| Low dose of 626 | 7 | 18.54 ± 0.58 d | 3.28 ± 0.32 d | 4.31 ± 0.45 d | 26.13 ± 1.03 e |
| 14 | 20.49 ± 0.24 b | 2.20 ± 0.19 b | 3.89 ± 0.46 d | 26.58 ± 0.55 c | |
| 17 | 28.33 ± 0.59 c | 2.08 ± 0.01 a,b | 3.46 ± 0.17 c | 33.87 ± 0.75 c | |
| High dose of 667 | 7 | 16.15 ± 0.35 c | 1.88 ± 0.07 b | 3.21 ± 0.22 b | 21.24 ± 0.36 c |
| 14 | 21.71 ± 6.51 b | 2.48 ± 0.56 b,c | 4.23 ± 1.23 e | 28.42 ± 7.42 d | |
| 17 | 41.29 ± 0.91 e | 2.37 ± 0.16 b,c | 5.17 ± 0.03 e | 48.83 ± 0.83 d | |
| Low dose of 667 | 7 | 20.45 ± 0.12 e | 1.88 ± 0.11 b | 3.63 ± 0.25 c | 25.96 ± 0.38 e |
| 14 | 28.59 ± 0.30 c | 2.24 ± 0.19 b | 4.51 ± 0.40 e | 35.34 ± 0.86 e | |
| 17 | 43.54 ± 3.35 e | 3.19 ± 0.56 c,d | 5.25 ± 0.98 e | 51.97 ± 1.90 e |
| Groups | p < 0.05 | |
|---|---|---|
| Normal | Control | |
| Normal | - | 0.8176 |
| Control | 0.8176 | - |
| Phenolphthalein | 0.3694 | 0.2912 |
| ST-III | 0.4562 | 0.2821 |
| High 669 | 0.0174 | 0.0074 |
| Low 669 | 0.0711 | 0.017 |
| High 667 | 0.0069 | 0.0032 |
| Low 667 | 0.0398 | 0.0037 |
| High 626 | 0.1436 | 0.0687 |
| Low 626 | 0.8034 | 0.4104 |
| Groups | Test Index (ng/L) | |||||
|---|---|---|---|---|---|---|
| MTL | Gas | SP | SS | ET-1 | VIP | |
| Normal | 176.3 ± 7.25 c | 206.44 ± 0.18 d | 58.22 ± 3.4 c | 61.41 ± 1.5 a,b,c | 58.48 ± 5.99 b | 42.02 ± 7.14 a |
| Control | 124.15 ± 16.6 a | 114.37 ± 2.39 b | 49.2 ± 7.44 a,b | 96.16 ± 5.68 d | 105.95 ± 1.95 d | 67.73 ± 5.37 d |
| Phenolphthalein | 144.76 ± 4.04 b | 137.35 ± 12 b,c | 49.9 ± 5.89 a,b | 53.98 ± 1.08 a | 60.47 ± 11.69 b | 50.3 ± 4.36 c |
| High dose of ST-III | 174.44 ± 7.89 c | 126.8 ± 1.5 b | 54.78 ± 4.16 c | 64.16 ± 2.41 b,c | 75.94 ± 3.48 c | 49.05 ± 3.36 b,c |
| Low dose of ST-III | 155.35 ± 7.89 c | 144.76 ± 3.79 c | 70.04 ± 4.16 d | 64.16 ± 2.41 b,c | 75.94 ± 3.48 c | 70.10 ± 3.63 e |
| High dose of 669 | 174.06 ± 21.68 c | 144.01 ± 1.3 c | 64.6 ± 1.51 c,d | 58.96 ± 3.49 a,b,c | 60.87 ± 3.27 b | 45.85 ± 5.53 a,b |
| Low dose of 669 | 137.11 ± 3.09 a,b | 121.41 ± 8.3 b | 64.6 ± 1.51 c,d | 65.99 ± 4.38 c | 63.45 ± 4.88 b | 66.78 ± 4.34 d,e |
| High dose of 626 | 143.40 ± 5.19 b | 132.21 ± 3.54 b,c | 50.86 ± 3.48 a,b | 97.23 ± 3.49 d | 91.66 ± 6.48 d | 61.92 ± 3.13 d |
| Low dose of 626 | 138.72 ± 3.02 a,b | 65.66 ± 6.98 a | 46.99 ± 3.48 a | 97.33 ± 4.38 d | 93.68 ± 5.59 d | 61.92 ± 3.13 d |
| High dose of 667 | 199.83 ± 10.59 d | 255.77 ± 27.5 e | 70.04 ± 1.08 d | 56.29 ± 0.54 a,b | 53.36 ± 2.13 a | 44.93 ± 3.57 a,b,c |
| Low dose of 667 | 140.20 ± 1.93 b | 132.21 ± 3.54 b,c | 50.86 ± 1.08 a,b | 65.98 ± 0.54 c | 63.45 ± 4.88 b | 44.93 ± 3.57 a,b,c |
| Group | Treatment | |
|---|---|---|
| 1–14 Days | 15–17 Days | |
| Normal | Normal saline solution | Normal saline solution |
| Control | Normal saline solution | loperamide |
| Phenolphthalein control | phenolphthalein | loperamide |
| High dose of ST-III | ST-III | loperamide |
| Low dose of ST-III | ST-III | loperamide |
| High dose of 626 | 626 | loperamide |
| Low dose of 626 | 626 | loperamide |
| High dose of 667 | 667 | loperamide |
| Low dose of 667 | 667 | loperamide |
| High dose of 669 | 669 | loperamide |
| Low dose of 669 | 669 | loperamide |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Hu, L.; Xu, Q.; Yin, B.; Fang, D.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice. Int. J. Mol. Sci. 2017, 18, 318. https://doi.org/10.3390/ijms18020318
Wang L, Hu L, Xu Q, Yin B, Fang D, Wang G, Zhao J, Zhang H, Chen W. Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice. International Journal of Molecular Sciences. 2017; 18(2):318. https://doi.org/10.3390/ijms18020318
Chicago/Turabian StyleWang, Linlin, Lujun Hu, Qi Xu, Boxing Yin, Dongsheng Fang, Gang Wang, Jianxin Zhao, Hao Zhang, and Wei Chen. 2017. "Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice" International Journal of Molecular Sciences 18, no. 2: 318. https://doi.org/10.3390/ijms18020318
APA StyleWang, L., Hu, L., Xu, Q., Yin, B., Fang, D., Wang, G., Zhao, J., Zhang, H., & Chen, W. (2017). Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice. International Journal of Molecular Sciences, 18(2), 318. https://doi.org/10.3390/ijms18020318