
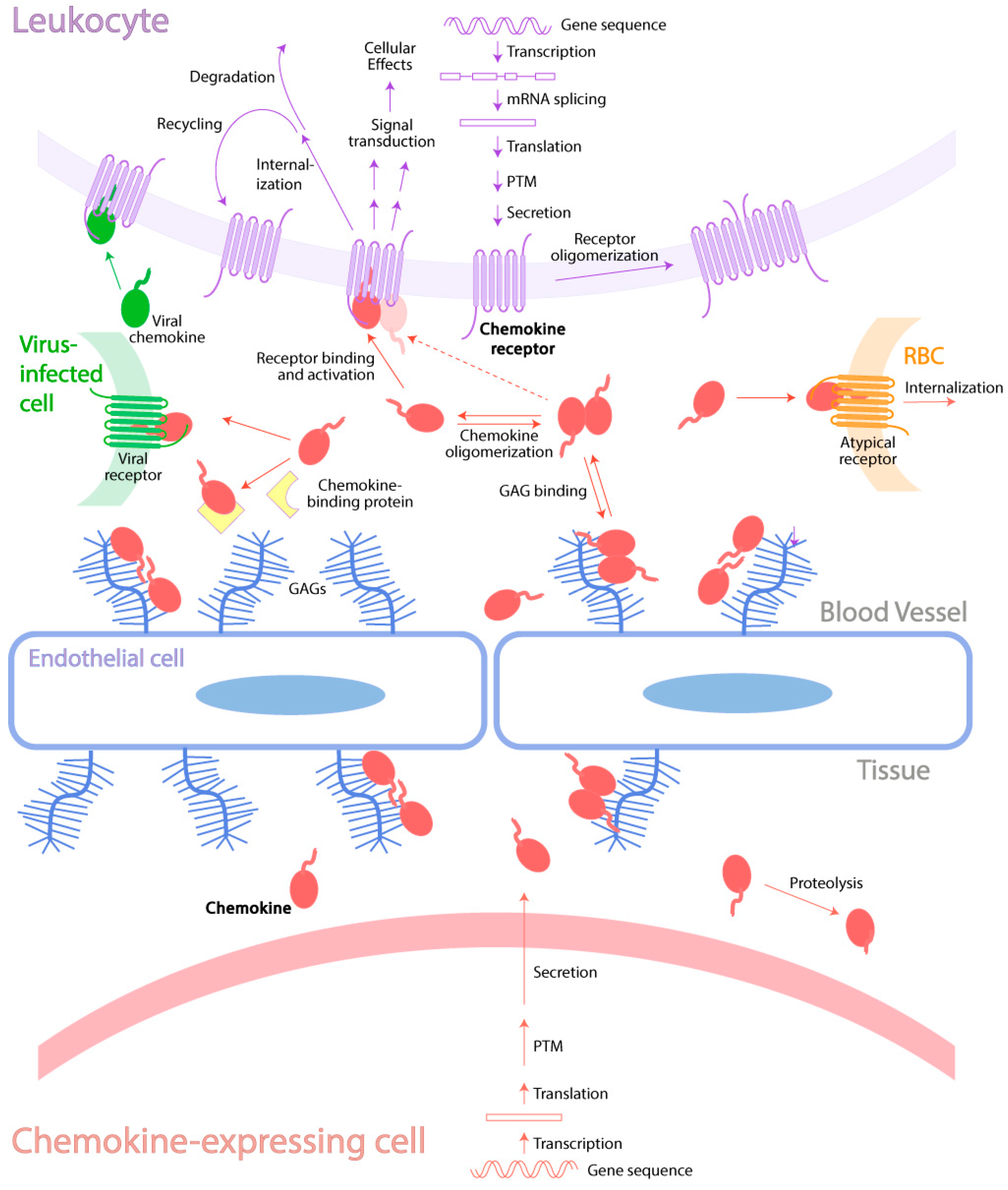
Mechanisms of Regulation of the Chemokine-Receptor Network

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The Chemokine and Chemokine Receptor Protein Families
2.1. The Chemokine Protein Family
2.2. The Chemokine Receptor Protein Family
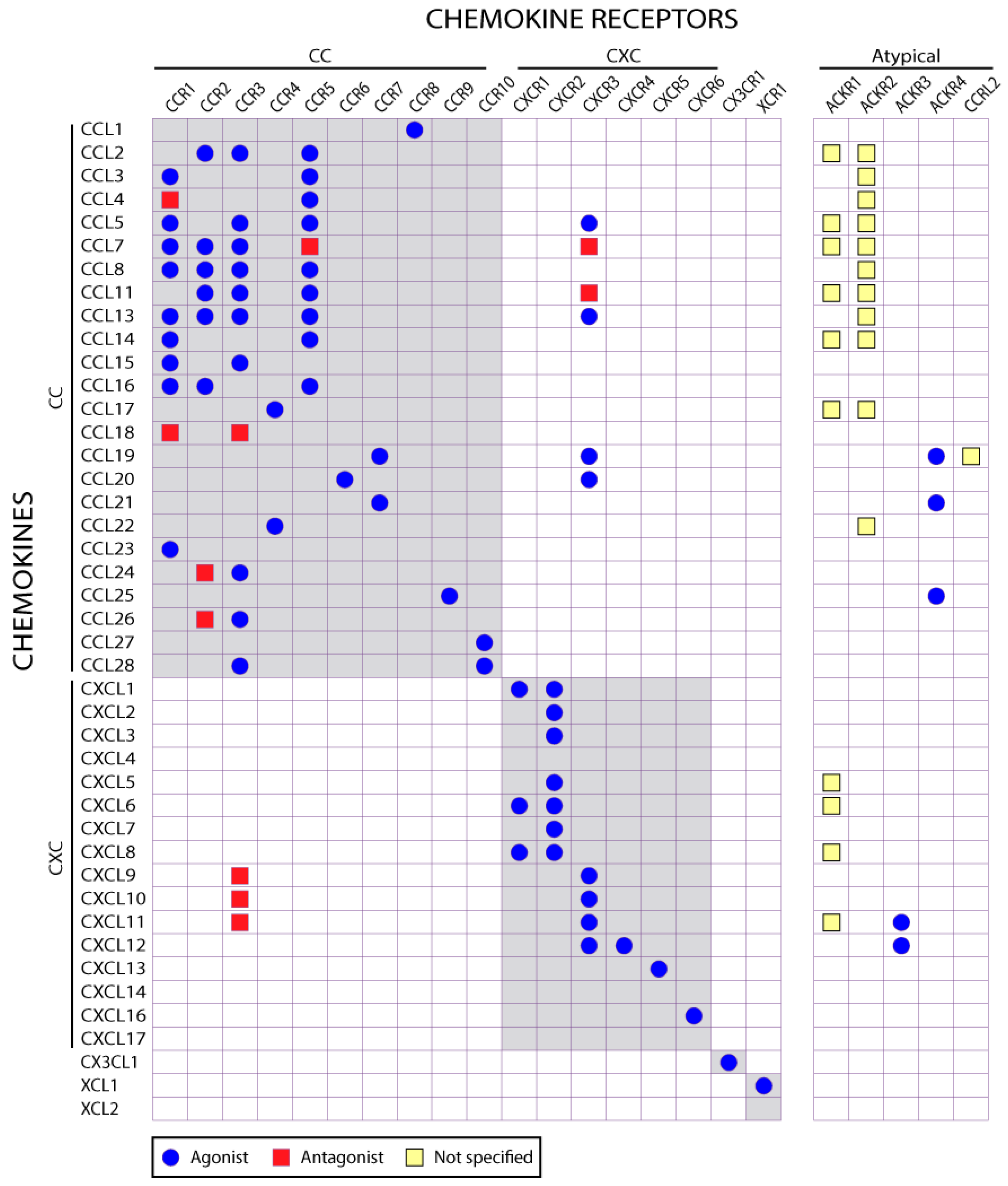
2.3. Selectivity of Chemokine-Receptor Interactions
2.4. Structural Basis of Chemokine-Receptor Recognition
3. Genetic and mRNA Splice Variants of Chemokines and Receptors
3.1. Variation Between Species
3.2. Polymorphisms in Chemokine Genes
3.3. Polymorphisms in Chemokine Receptor Genes
3.4. Chemokine Receptor Splice Variants
4. Regulation of Expression, Degradation and Localization
4.1. Expression
4.2. Internalization and Recycling or Degradation
4.3. Atypical (Decoy) Chemokine Receptors as Chemokine Scavengers
4.4. Localization by Binding to Glycosaminoglycans (GAGs)
5. Post-Translational Modifications
5.1. Proteolytic Processing of Chemokines
5.2. Other Post-Translational Modifications of Chemokines
5.3. Post-Translational Modifications of Chemokine Receptors
6. Oligomerization of Chemokines and Chemokine Receptors
6.1. Oligomerization of Chemokines
6.2. Oligomerization of Chemokine Receptors
7. Regulation of Signaling Pathways
7.1. Overview of Signaling Pathways—G Proteins and Arrestins
7.2. Regulation of Signaling in Different Cellular Contexts
7.3. Partial Agonism
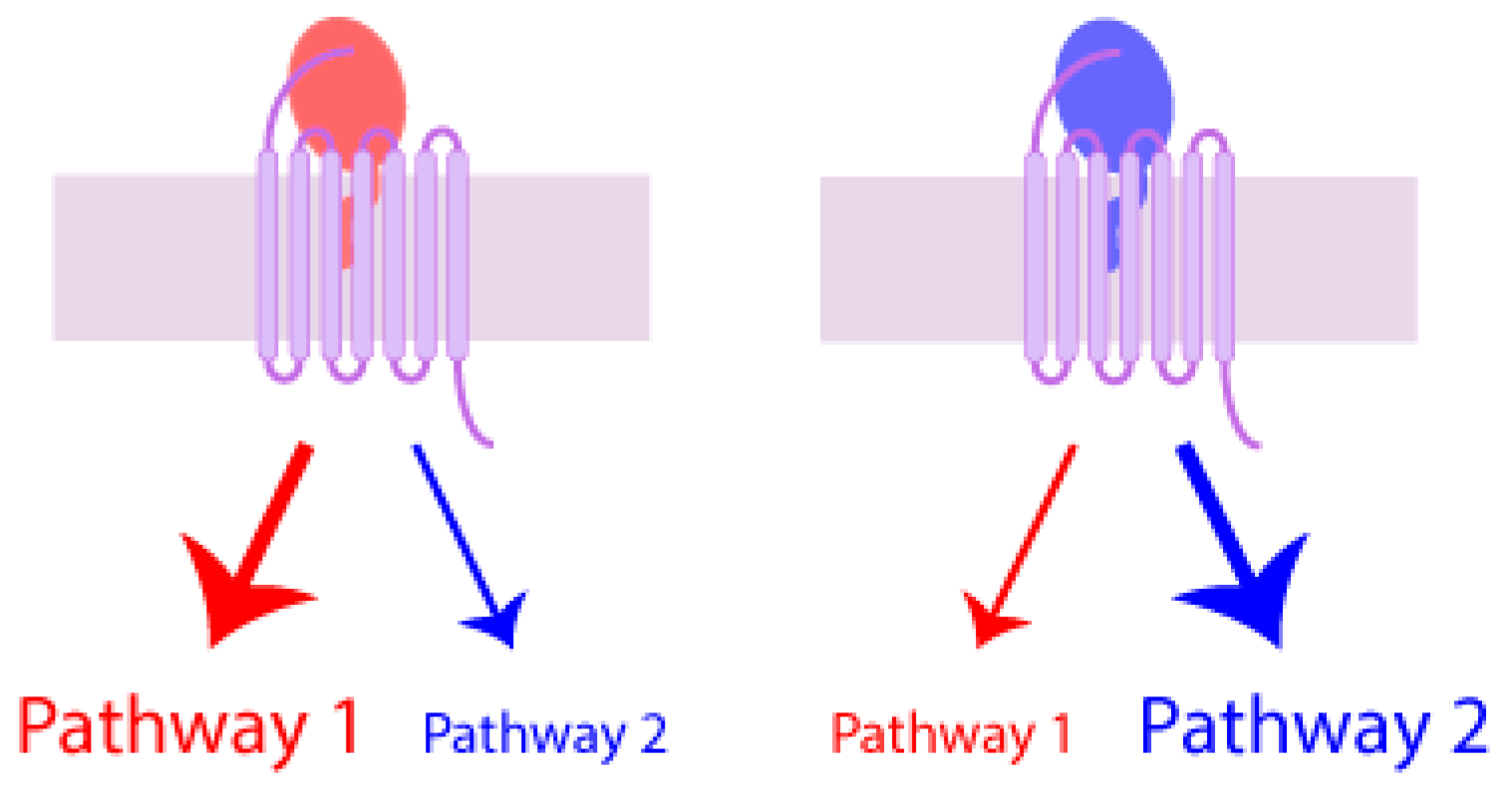
7.4. Biased Agonism
8. Natural and Pharmacological Inhibitors
8.1. Viral Chemokines and Receptors
8.2. Chemokine-Binding Proteins from Pathogens and Parasites
8.3. Pharmacological Approaches towards Inhibition of Chemokines and Receptors
9. Summary and Future Directions
Acknowledgments
Conflicts of Interest
Abbreviations
| AP2 | Adaptin 2 |
| CKBP | Chemokine-binding protein |
| DC-CK | Dendritic cell-derived CC chemokine |
| ENA | Epithelial-derived neutrophil-activating |
| GCP | Granulocyte chemotactic protein |
| GRO | Growth-regulated protein |
| IL | Interleukin |
| IP | Interferon γ-induced protein |
| IUPHAR | International Union of Basic and Clinical Pharmacology |
| I-TAC | Interferon-inducible T-cell α chemoattractant |
| MCP | Monocyte chemoattractant protein |
| MGSA | Melanoma growth stimulating activity |
| MIP | Macrophage inflammatory protein |
| PARC | Pulmonary and activation-regulated chemokine |
| PF4 | Platelet factor 4 |
| RANTES | Regulated on activation, normal T cell expressed and secreted |
| SDF | Stromal cell-derived factor |
| SECRET | Smallpox virus-encoded chemokine receptor |
| SLC | Secondary lymphoid tissue chemokine |
| SNP | Single nucleotide polymorphism |
| TARC | Thymus and activation regulated chemokine |
| TECK | Thymus-expressed chemokine |
References
- Moser, B.; Wolf, M.; Walz, A.; Loetscher, P. Chemokines: Multiple levels of leukocyte migration control. Trends Immunol. 2004, 25, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M. Chemokines in pathology and medicine. J. Intern. Med. 2001, 250, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Gerard, C.; Rollins, B.J. Chemokines and disease. Nat. Immunol. 2001, 2, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Dimberg, A. Chemokines in angiogenesis. Curr. Top. Microbiol. Immunol. 2010, 341, 59–80. [Google Scholar] [PubMed]
- Speyer, C.L.; Ward, P.A. Role of endothelial chemokines and their receptors during inflammation. J. Invest. Surg. 2011, 24, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Ben-Baruch, A. The multifaceted roles of chemokines in malignancy. Cancer Metastasis Rev. 2006, 25, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Luther, S.A.; Cyster, J.G. Chemokines as regulators of T cell differentiation. Nat. Immunol. 2001, 2, 102–107. [Google Scholar] [CrossRef]
- Choe, H.; Moore, M.J.; Owens, C.M.; Wright, P.L.; Vasilieva, N.; Li, W.; Singh, A.P.; Shakri, R.; Chitnis, C.E.; Farzan, M. Sulphated tyrosines mediate association of chemokines and Plasmodium vivax duffy binding protein with the duffy antigen/receptor for chemokines (DARC). Mol. Microbiol. 2005, 55, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, G.; Combadiere, C.; Broder, C.C.; Feng, Y.; Kennedy, P.E. CC CKR5: A RANTES, MIP-1a, MIP-1b receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 1996, 272, 1955–1958. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E. Chemokine receptors: Multifaceted therapeutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Horuk, R.; Rice, G.C.; Bennett, G.L.; Camerato, T.; Wood, W.I. Characterization of two high affinity human interleukin-8 receptors. J. Biol. Chem. 1992, 267, 16283–16287. [Google Scholar] [PubMed]
- Stone, M.J.; Mayer, K.L. Three dimensional structure of chemokines. In Chemokines in Allergic Disease; Rothenberg, M.E., Ed.; Marcel Dekker: New York, NY, USA, 2000; pp. 67–94. [Google Scholar]
- Burg, J.S.; Ingram, J.R.; Venkatakrishnan, A.J.; Jude, K.M.; Dukkipati, A.; Feinberg, E.N.; Angelini, A.; Waghray, D.; Dror, R.O.; Ploegh, H.L.; et al. Structural basis for chemokine recognition and activation of a viral G protein–coupled receptor. Science 2015, 347, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Kufareva, I.; Holden, L.G.; Wang, C.; Zheng, Y.; Zhao, C.; Fenalti, G.; Wu, H.; Han, G.W.; Cherezov, V.; et al. Crystal structure of the chemokine receptor CXCR4 in complex with a viral chemokine. Science 2015, 347, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Chien, E.Y.; Mol, C.D.; Fenalti, G.; Liu, W.; Katritch, V.; Abagyan, R.; Brooun, A.; Wells, P.; Bi, F.C.; et al. Structures of the CXCR4 chemokine GPCR with small-molecule and cyclic peptide antagonists. Science 2010, 330, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Das, B.B.; Casagrande, F.; Tian, Y.; Nothnagel, H.J.; Chu, M.; Kiefer, H.; Maier, K.; de Angelis, A.A.; Marassi, F.M.; et al. Structure of the chemokine receptor CXCR1 in phospholipid bilayers. Nature 2012, 491, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Zhu, Y.; Li, J.; Chen, Z.; Han, G.W.; Kufareva, I.; Li, T.; Ma, L.; Fenalti, G.; Zhang, W.; et al. Structure of the CCR5 chemokine receptor-HIV entry inhibitor maraviroc complex. Science 2013, 341, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Lubkowski, J.; Bujacz, G.; Boque, L.; Domaille, P.J.; Handel, T.M.; Wlodawer, A. The structure of MCP-1 in two crystal forms provides a rare example of variable quaternary interactions. Nat. Struct. Biol. 1997, 4, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Wescott, M.P.; Kufareva, I.; Paes, C.; Goodman, J.R.; Thaker, Y.; Puffer, B.A.; Berdougo, E.; Rucker, J.B.; Handel, T.M.; Doranz, B.J. Signal transmission through the CXC chemokine receptor 4 (CXCR4) transmembrane helices. Proc. Natl. Acad. Sci. USA 2016, 113, 9928–9933. [Google Scholar] [CrossRef] [PubMed]
- Skelton, N.J.; Quan, C.; Reilly, D.; Lowman, H. Structure of a CXC chemokine-receptor fragment in complex with interleukin-8. Structure 1999, 7, 157–168. [Google Scholar] [CrossRef]
- Veldkamp, C.T.; Seibert, C.; Peterson, F.C.; de la Cruz, N.B.; Haugner, J.C., 3rd; Basnet, H.; Sakmar, T.P.; Volkman, B.F. Structural basis of CXCR4 sulfotyrosine recognition by the chemokine SDF-1/CXCL12. Sci. Signal. 2008, 1, ra4. [Google Scholar] [CrossRef] [PubMed]
- Millard, C.J.; Ludeman, J.P.; Canals, M.; Bridgford, J.L.; Hinds, M.G.; Clayton, D.J.; Christopoulos, A.; Payne, R.J.; Stone, M.J. Structural basis of receptor sulfotyrosine recognition by a CC chemokine: The N-terminal region of CCR3 bound to CCL11/eotaxin-1. Structure 2014, 22, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Crump, M.P.; Gong, J.H.; Loetscher, P.; Rajarathnam, K.; Amara, A.; Arenzana-Seisdedos, F.; Virelizier, J.L.; Baggiolini, M.; Sykes, B.D.; Clark-Lewis, I. Solution structure and basis for functional activity of stromal cell-derived factor-1; dissociation of CXCR4 activation from binding and inhibition of HIV-1. EMBO J. 1997, 16, 6996–7007. [Google Scholar] [CrossRef] [PubMed]
- Kleist, A.B.; Getschman, A.E.; Ziarek, J.J.; Nevins, A.M.; Gauthier, P.A.; Chevigne, A.; Szpakowska, M.; Volkman, B.F. New paradigms in chemokine receptor signal transduction: Moving beyond the two-site model. Biochem. Pharmacol. 2016, 114, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Diversity and modularity of G protein-coupled receptor structures. Trends Pharmacol. Sci. 2012, 33, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Bajoghli, B.; Aghaallaei, N.; Hess, I.; Rode, I.; Netuschil, N.; Tay, B.H.; Venkatesh, B.; Yu, J.K.; Kaltenbach, S.L.; Holland, N.D.; et al. Evolution of genetic networks underlying the emergence of thymopoiesis in vertebrates. Cell 2009, 138, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Nomiyama, H.; Osada, N.; Yoshie, O. A family tree of vertebrate chemokine receptors for a unified nomenclature. Dev. Comp. Immunol. 2011, 35, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O.; Nomiyama, H. The chemokine and chemokine receptor superfamilies and their molecular evolution. Genome Biol. 2006, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.H.; Yang, Q.; Kathiresan, S.; Cupples, L.A.; Massaro, J.M.; Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Hirschhorn, J.N.; O’Donnell, C.J.; et al. CCL2 polymorphisms are associated with serum monocyte chemoattractant protein-1 levels and myocardial infarction in the framingham heart study. Circulation 2005, 112, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Flex, A.; Gaetani, E.; Papaleo, P.; Straface, G.; Proia, A.S.; Pecorini, G.; Tondi, P.; Pola, P.; Pola, R. Proinflammatory genetic profiles in subjects with history of ischemic stroke. Stroke 2004, 35, 2270–2275. [Google Scholar] [CrossRef] [PubMed]
- Karrer, S.; Bosserhoff, A.K.; Weiderer, P.; Distler, O.; Landthaler, M.; Szeimies, R.M.; Muller-Ladner, U.; Scholmerich, J.; Hellerbrand, C. The -2518 promotor polymorphism in the MCP-1 gene is associated with systemic sclerosis. J. Investig. Dermatol. 2005, 124, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kroner, A.; Maurer, M.; Loserth, S.; Kleinschnitz, C.; Hemmer, B.; Rosche, B.; Toyka, K.V.; Rieckmann, P. Analysis of the monocyte chemoattractant protein 1 -2518 promoter polymorphism in patients with multiple sclerosis. Tissue Antigens 2004, 64, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Escribano, M.F.; Torres, B.; Aguilar, F.; Rodriguez, R.; Garcia, A.; Valenzuela, A.; Nunez-Roldan, A. MCP-1 promoter polymorphism in Spanish patients with rheumatoid arthritis. Hum. Immunol. 2003, 64, 741–744. [Google Scholar] [CrossRef]
- Porcellini, E.; Ianni, M.; Carbone, I.; Franceschi, M.; Licastro, F. Monocyte chemoattractant protein-1 promoter polymorphism and plasma levels in Alzheimer’s disease. Immun. Ageing 2013, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.; Rovin, B.H.; Sen, L.; Cooke, G.; Dhanda, R.; Mummidi, S.; Kulkarni, H.; Bamshad, M.J.; Telles, V.; Anderson, S.A.; et al. HIV-1 infection and aids dementia are influenced by a mutant MCP-1 allele linked to increased monocyte infiltration of tissues and MCP-1 levels. Proc. Natl. Acad. Sci. USA 2002, 99, 13795–13800. [Google Scholar] [CrossRef] [PubMed]
- Hellier, S.; Frodsham, A.J.; Hennig, B.J.; Klenerman, P.; Knapp, S.; Ramaley, P.; Satsangi, J.; Wright, M.; Zhang, L.; Thomas, H.C.; et al. Association of genetic variants of the chemokine receptor CCR5 and its ligands, RANTES and MCP-2, with outcome of HCV infection. Hepatology 2003, 38, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Tsunemi, Y.; Komine, M.; Sekiya, T.; Saeki, H.; Nakamura, K.; Hirai, K.; Kakinuma, T.; Kagami, S.; Fujita, H.; Asano, N.; et al. The -431C→T polymorphism of thymus and activation-regulated chemokine increases the promoter activity but is not associated with susceptibility to atopic dermatitis in Japanese patients. Exp. Dermatol. 2004, 13, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.; Modi, W.; Smith, M.W.; Nelson, G.W.; Wu, X.; Carrington, M.; Dean, M.; Honjo, T.; Tashiro, K.; Yabe, D.; et al. Genetic restriction of AIDS pathogenesis by an SDF-1 chemokine gene variant. Alive study, hemophilia growth and development study (HGDS), multicenter AIDS cohort study (MACS), multicenter hemophilia cohort study (MHCS), San Francisco city cohort (SFCC). Science 1998, 279, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Reiche, E.M.; Bonametti, A.M.; Voltarelli, J.C.; Morimoto, H.K.; Watanabe, M.A. Genetic polymorphisms in the chemokine and chemokine receptors: Impact on clinical course and therapy of the human immunodeficiency virus type 1 infection (HIV-1). Curr. Med. Chem. 2007, 14, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Ortlepp, J.R.; Krantz, C.; Kimmel, M.; von Korff, A.; Vesper, K.; Schmitz, F.; Mevissen, V.; Janssens, U.; Franke, A.; Hanrath, P.; et al. Additive effects of the chemokine receptor 2, vitamin D receptor, interleukin-6 polymorphisms and cardiovascular risk factors on the prevalence of myocardial infarction in patients below 65 years. Int. J. Cardiol. 2005, 105, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Petrkova, J.; Cermakova, Z.; Drabek, J.; Lukl, J.; Petrek, M. CC chemokine receptor (CCR)2 polymorphism in Czech patients with myocardial infarction. Immunol. Lett. 2003, 88, 53–55. [Google Scholar] [CrossRef]
- Miyagishi, R.; Niino, M.; Fukazawa, T.; Yabe, I.; Kikuchi, S.; Tashiro, K. C-C chemokine receptor 2 gene polymorphism in Japanese patients with multiple sclerosis. J. Neuroimmunol. 2003, 145, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Goulding, C.; McManus, R.; Murphy, A.; MacDonald, G.; Barrett, S.; Crowe, J.; Hegarty, J.; McKiernan, S.; Kelleher, D. The CCR5-D32 mutation: Impact on disease outcome in individuals with hepatitis C infection from a single source. Gut 2005, 54, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Navratilova, Z. Polymorphisms in CCL2 & CCL5 chemokines/chemokine receptors genes and their association with diseases. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czechoslov. 2006, 150, 191–204. [Google Scholar]
- Smith, M.W.; Dean, M.; Carrington, M.; Winkler, C.; Huttley, G.A.; Lomb, D.A.; Goedert, J.J.; O’Brien, T.R.; Jacobson, L.P.; Kaslow, R.; et al. Contrasting genetic influence of CCR2 and CCR5 variants on HIV-1 infection and disease progression. Hemophilia growth and development study (HGDS), multicenter aids cohort study (MACS), multicenter hemophilia cohort study (MHCS), San Francisco city cohort (SFCC), alive study. Science 1997, 277, 959–965. [Google Scholar] [PubMed]
- Burton, C.T.; Gotch, F.M.; Imami, N. CCR2/64I mutation detection in a HIV-1-positive patient with slow CD4 T-cell decline and delay in disease progression. Int. J. STD AIDS 2005, 16, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Paxton, W.A.; Wolinsky, S.M.; Neumann, A.U.; Zhang, L.; He, T.; Kang, S.; Ceradini, D.; Jin, Z.; Yazdanbakhsh, K.; et al. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nat. Med. 1996, 2, 1240–1243. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, P.A.; Buckler-White, A.; Alkhatib, G.; Spalding, T.; Kubofcik, J.; Combadiere, C.; Weissman, D.; Cohen, O.; Rubbert, A.; Lam, G.; et al. Inherited resistance to HIV-1 conferred by an inactivating mutation in CC chemokine receptor 5: Studies in populations with contrasting clinical phenotypes, defined racial background, and quantified risk. Mol. Med. 1997, 3, 23–36. [Google Scholar] [PubMed]
- Suresh, P.; Wanchu, A. Chemokines and chemokine receptors in HIV infection: Role in pathogenesis and therapeutics. J. Postgrad. Med. 2006, 52, 210–217. [Google Scholar] [PubMed]
- Paxton, W.A.; Kang, S. Chemokine receptor allelic polymorphisms: Relationships to HIV resistance and disease progression. Semin. Immunol. 1998, 10, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Walton, R.T.; Rowland-Jones, S.L. HIV and chemokine binding to red blood cells—DARC matters. Cell Host Microbe 2008, 4, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Charo, I.F.; Myers, S.J.; Herman, A.; Franci, C.; Connolly, A.J.; Coughlin, S.R. Molecular cloning and functional expression of two monocyte chemoattractant protein 1 receptors reveals alternative splicing of the carboxyl-terminal tails. Proc. Natl. Acad. Sci. USA 1994, 91, 2752–2756. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, C.; Civatte, M.; Pellissier, J.F.; Figarella-Branger, D. CCR2A and CCR2B, the two isoforms of the monocyte chemoattractant protein-1 receptor are up-regulated and expressed by different cell subsets in idiopathic inflammatory myopathies. Acta Neuropathol. 2001, 102, 385–392. [Google Scholar] [PubMed]
- Berchiche, Y.A.; Sakmar, T.P. CXC chemokine receptor 3 alternative splice variants selectively activate different signaling pathways. Mol. Pharmacol. 2016, 90, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Burkhardt, A.M.; Homey, B. Homeostatic chemokine receptors and organ-specific metastasis. Nat. Rev. Immunol. 2011, 11, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.J.; Krensky, A.M. Chemokines, chemokine receptors, and allograft rejection. Immunity 2001, 14, 377–386. [Google Scholar] [CrossRef]
- Link, D.C. Neutrophil homeostasis: A new role for stromal cell-derived factor-1. Immunol. Res. 2005, 32, 169–178. [Google Scholar] [CrossRef]
- Uy, G.L.; Rettig, M.P.; Cashen, A.F. Plerixafor, a CXCR4 antagonist for the mobilization of hematopoietic stem cells. Expert. Opin. Biol. Ther. 2008, 8, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Vicari, A.P.; Figueroa, D.J.; Hedrick, J.A.; Foster, J.S.; Singh, K.P.; Menon, S.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Bacon, K.B.; et al. Teck: A novel CC chemokine specifically expressed by thymic dendritic cells and potentially involved in T cell development. Immunity 1997, 7, 291–301. [Google Scholar] [CrossRef]
- Link, A.; Vogt, T.K.; Favre, S.; Britschgi, M.R.; Acha-Orbea, H.; Hinz, B.; Cyster, J.G.; Luther, S.A. Fibroblastic reticular cells in lymph nodes regulate the homeostasis of naive T cells. Nat. Immunol. 2007, 8, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Jameson, S.C. Naive T cell homeostasis: From awareness of space to a sense of place. Nat. Rev. Immunol. 2009, 9, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A. Endocytic trafficking of chemokine receptors. Curr. Opin. Cell Biol. 2014, 27, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Okamoto, H.; Toyama, Y.; Momohara, S. Molecular aspects of rheumatoid arthritis: Chemokines in the joints of patients. FEBS J. 2008, 275, 4448–4455. [Google Scholar] [CrossRef] [PubMed]
- Niwa, Y.; Akamatsu, H.; Niwa, H.; Sumi, H.; Ozaki, Y.; Abe, A. Correlation of tissue and plasma RANTES levels with disease course in patients with breast or cervical cancer. Clin. Cancer. Res. 2001, 7, 285–289. [Google Scholar] [PubMed]
- Soria, G.; Ben-Baruch, A. The inflammatory chemokines CCL2 and CCL5 in breast cancer. Cancer Lett. 2008, 267, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Toi, M.; Saji, H.; Muta, M.; Bando, H.; Kuroi, K.; Koike, M.; Inadera, H.; Matsushima, K. Significance of macrophage chemoattractant protein-1 in macrophage recruitment, angiogenesis, and survival in human breast cancer. Clin. Cancer. Res. 2000, 6, 3282–3289. [Google Scholar] [PubMed]
- Neel, N.F.; Schutyser, E.; Sai, J.; Fan, G.H.; Richmond, A. Chemokine receptor internalization and intracellular trafficking. Cytokine Growth Factor Rev. 2005, 16, 637–658. [Google Scholar] [CrossRef] [PubMed]
- Baugher, P.J.; Richmond, A. The carboxyl-terminal PDZ ligand motif of chemokine receptor CXCR2 modulates post-endocytic sorting and cellular chemotaxis. J. Biol. Chem. 2008, 283, 30868–30878. [Google Scholar] [CrossRef] [PubMed]
- Delhaye, M.; Gravot, A.; Ayinde, D.; Niedergang, F.; Alizon, M.; Brelot, A. Identification of a post-endocytic sorting sequence in CCR5. Mol. Pharmacol. 2007, 72, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Busillo, J.M.; Benovic, J.L. Regulation of CXCR4 signaling. Biochim. Biophys. Acta 2007, 1768, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Benovic, J.L. Agonist-promoted ubiquitination of the G protein-coupled receptor CXCR4 mediates lysosomal sorting. J. Biol. Chem. 2001, 276, 45509–45512. [Google Scholar] [CrossRef] [PubMed]
- Canals, M.; Scholten, D.J.; de Munnik, S.; Han, M.K.; Smit, M.J.; Leurs, R. Ubiquitination of CXCR7 controls receptor trafficking. PLoS ONE 2012, 7, e34192. [Google Scholar] [CrossRef] [PubMed]
- Schaeuble, K.; Hauser, M.A.; Rippl, A.V.; Bruderer, R.; Otero, C.; Groettrup, M.; Legler, D.F. Ubiquitylation of the chemokine receptor CCR7 enables efficient receptor recycling and cell migration. J. Cell Sci. 2012, 125, 4463–4474. [Google Scholar] [CrossRef] [PubMed]
- Bachelerie, F.; Ben-Baruch, A.; Burkhardt, A.M.; Combadiere, C.; Farber, J.M.; Graham, G.J.; Horuk, R.; Sparre-Ulrich, A.H.; Locati, M.; Luster, A.D.; et al. International union of basic and clinical pharmacology. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol. Rev. 2014, 66, 1–79. [Google Scholar] [CrossRef] [PubMed]
- Bachelerie, F.; Graham, G.J.; Locati, M.; Mantovani, A.; Murphy, P.M.; Nibbs, R.; Rot, A.; Sozzani, S.; Thelen, M. New nomenclature for atypical chemokine receptors. Nat. Immunol. 2014, 15, 207–208. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Savino, B.; Borroni, E.M.; Mantovani, A.; Locati, M. Chemokine decoy receptors: Structure-function and biological properties. Curr. Top. Microbiol. Immunol. 2010, 341, 15–36. [Google Scholar] [PubMed]
- Massara, M.; Bonavita, O.; Mantovani, A.; Locati, M.; Bonecchi, R. Atypical chemokine receptors in cancer: Friends or foes? J. Leukoc. Biol. 2016, 99, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bonecchi, R.; Locati, M. Tuning inflammation and immunity by chemokine sequestration: Decoys and more. Nat. Rev. Immunol. 2006, 6, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Vacchini, A.; Locati, M.; Borroni, E.M. Overview and potential unifying themes of the atypical chemokine receptor family. J. Leukoc. Biol. 2016, 99, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Fra, A.M.; Locati, M.; Otero, K.; Sironi, M.; Signorelli, P.; Massardi, M.L.; Gobbi, M.; Vecchi, A.; Sozzani, S.; Mantovani, A. Cutting edge: Scavenging of inflammatory CC chemokines by the promiscuous putatively silent chemokine receptor D6. J. Immunol. 2003, 170, 2279–2282. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, T.; Cook, D.N.; Nibbs, R.J.; Rot, A.; Nixon, C.; McLean, P.; Alcami, A.; Lira, S.A.; Wiekowski, M.; Graham, G.J. The chemokine receptor D6 limits the inflammatory response in vivo. Nat. Immunol. 2005, 6, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sharp, J.S.; Handel, T.M.; Prestegard, J.H. Chemokine oligomerization in cell signaling and migration. Prog. Mol. Biol. Transl. Sci. 2013, 117, 531–578. [Google Scholar] [PubMed]
- Salanga, C.L.; Handel, T.M. Chemokine oligomerization and interactions with receptors and glycosaminoglycans: The role of structural dynamics in function. Exp. Cell Res. 2011, 317, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E.I.; Handel, T.M.; Johnson, Z.; Lau, E.K.; LiWang, P.; Clark-Lewis, I.; Borlat, F.; Wells, T.N.C.; Kosco-Vilbois, M.H. Glycosaminoglycan binding and oligomerization are essential for the in vivo activity of certain chemokines. Proc. Natl. Acad. Sci. USA 2003, 100, 1885–1890. [Google Scholar] [CrossRef] [PubMed]
- Gangavarapu, P.; Rajagopalan, L.; Kolli, D.; Guerrero-Plata, A.; Garofalo, R.P.; Rajarathnam, K. The monomer-dimer equilibrium and glycosaminoglycan interactions of chemokine CXCL8 regulate tissue-specific neutrophil recruitment. J. Leukoc. Biol. 2012, 91, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Middleton, J.; Patterson, A.M.; Gardner, L.; Schmutz, C.; Ashton, B.A. Leukocyte extravasation: Chemokine transport and presentation by the endothelium. Blood 2002, 100, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Moelants, E.A.; Mortier, A.; van Damme, J.; Proost, P. In vivo regulation of chemokine activity by post-translational modification. Immunol. Cell Biol. 2013, 91, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Mortier, A.; van Damme, J.; Proost, P. Regulation of chemokine activity by posttranslational modification. Pharmacol. Ther. 2008, 120, 197–217. [Google Scholar] [CrossRef] [PubMed]
- Starr, A.E.; Dufour, A.; Maier, J.; Overall, C.M. Biochemical analysis of matrix metalloproteinase activation of chemokines CCL15 and CCL23 and increased glycosaminoglycan binding of CCL16. J. Biol. Chem. 2012, 287, 5848–5860. [Google Scholar] [CrossRef] [PubMed]
- Mortier, A.; Berghmans, N.; Ronsse, I.; Grauwen, K.; Stegen, S.; van Damme, J.; Proost, P. Biological activity of CXCL8 forms generated by alternative cleavage of the signal peptide or by aminopeptidase-mediated truncation. PLoS ONE 2011, 6, e23913. [Google Scholar] [CrossRef] [PubMed]
- Moelants, E.A.; Loozen, G.; Mortier, A.; Martens, E.; Opdenakker, G.; Mizgalska, D.; Szmigielski, B.; Potempa, J.; van Damme, J.; Teughels, W.; et al. Citrullination and proteolytic processing of chemokines by porphyromonas gingivalis. Infect. Immun. 2014, 82, 2511–2519. [Google Scholar] [CrossRef] [PubMed]
- Detheux, M.; Standker, L.; Vakili, J.; Munch, J.; Forssmann, U.; Adermann, K.; Pohlmann, S.; Vassart, G.; Kirchhoff, F.; Parmentier, M.; et al. Natural proteolytic processing of hemofiltrate CC chemokine 1 generates a potent CC chemokine receptor (CCR)1 and CCR5 agonist with anti-HIV properties. J. Exp. Med. 2000, 192, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Forssmann, U.; Hartung, I.; Balder, R.; Fuchs, B.; Escher, S.E.; Spodsberg, N.; Dulkys, Y.; Walden, M.; Heitland, A.; Braun, A.; et al. N-nonanoyl-CC chemokine ligand 14, a potent CC chemokine ligand 14 analogue that prevents the recruitment of eosinophils in allergic airway inflammation. J. Immunol. 2004, 173, 3456–3466. [Google Scholar] [CrossRef] [PubMed]
- Savino, B.; Borroni, E.M.; Torres, N.M.; Proost, P.; Struyf, S.; Mortier, A.; Mantovani, A.; Locati, M.; Bonecchi, R. Recognition versus adaptive up-regulation and degradation of CC chemokines by the chemokine decoy receptor d6 are determined by their N-terminal sequence. J. Biol. Chem. 2009, 284, 26207–26215. [Google Scholar] [CrossRef] [PubMed]
- Eckhard, U.; Huesgen, P.F.; Schilling, O.; Bellac, C.L.; Butler, G.S.; Cox, J.H.; Dufour, A.; Goebeler, V.; Kappelhoff, R.; Keller, U.A.; et al. Active site specificity profiling of the matrix metalloproteinase family: Proteomic identification of 4300 cleavage sites by nine mmps explored with structural and synthetic peptide cleavage analyses. Matrix Biol. 2016, 49, 37–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, M.; Clark-Lewis, I.; Buri, C.; Langen, H.; Lis, M.; Mazzucchelli, L. Cathepsin D specifically cleaves the chemokines macrophage inflammatory protein-1 a, macrophage inflammatory protein-1 b, and SLC that are expressed in human breast cancer. Am. J. Pathol. 2003, 162, 1183–1190. [Google Scholar] [CrossRef]
- Jiang, Y.; Tabak, L.A.; Valente, A.J.; Graves, D.T. Initial characterization of the carbohydrate structure of MCP-1. Biochem. Biophys. Res. Commun. 1991, 178, 1400–1404. [Google Scholar] [CrossRef]
- Jiang, Y.; Valente, A.J.; Williamson, M.J.; Zhang, L.; Graves, D.T. Post-translational modification of a monocyte-specific chemoattractant synthesized by glioma, osteosarcoma, and vascular smooth muscle cells. J. Biol. Chem. 1990, 265, 18318–18321. [Google Scholar] [PubMed]
- Noso, N.; Bartels, J.; Mallet, A.I.; Mochizuki, M.; Christophers, E.; Schroder, J.M. Delayed production of biologically active O-glycosylated forms of human eotaxin by tumor-necrosis-factor-α-stimulated dermal fibroblasts. Eur. J. Biochem. 1998, 253, 114–122. [Google Scholar] [CrossRef]
- Richter, R.; Schulz-Knappe, P.; John, H.; Forssmann, W.G. Posttranslationally processed forms of the human chemokine HCC-1. Biochemistry 2000, 39, 10799–10805. [Google Scholar] [CrossRef] [PubMed]
- Kameyoshi, Y.; Dorschner, A.; Mallet, A.I.; Christophers, E.; Schroder, J.M. Cytokine RANTES released by thrombin-stimulated platelets is a potent attractant for human eosinophils. J. Exp. Med. 1992, 176, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Hermand, P.; Pincet, F.; Carvalho, S.; Ansanay, H.; Trinquet, E.; Daoudi, M.; Combadiere, C.; Deterre, P. Functional adhesiveness of the cx3cl1 chemokine requires its aggregation. Role of the transmembrane domain. J. Biol. Chem. 2008, 283, 30225–30234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.D.; Goetze, A.M.; Bass, R.B.; Flynn, G.C. N-terminal glutamate to pyroglutamate conversion in vivo for human IgG2 antibodies. J. Biol. Chem. 2011, 286, 11211–11217. [Google Scholar] [CrossRef] [PubMed]
- Uguccioni, M.; Loetscher, P.; Forssmann, U.; Dewald, B.; Li, H.D.; Lima, S.H.; Li, Y.L.; Kreider, B.; Garotta, G.; Thelen, M.; et al. Monocyte chemotactic protein 4 (MCP-4), a novel structural and functional analogue of MCP-3 and eotaxin. J. Exp. Med. 1996, 183, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, J.; Proost, P.; Lenaerts, J.P.; Opdenakker, G. Structural and functional identification of 2 human, tumor-derived monocyte chemotactic proteins (MCP-2 and MCP-3) belonging to the chemokine family. J. Exp. Med. 1992, 176, 59–65. [Google Scholar] [CrossRef]
- Yoshimura, T.; Robinson, E.A.; Appella, E.; Matsushima, K.; Showalter, S.D.; Skeel, A.; Leonard, E.J. 3 forms of monocyte-derived neutrophil chemotactic factor (MDNCF) distinguished by different lengths of the amino-terminal sequence. Mol. Immunol. 1989, 26, 87–93. [Google Scholar] [PubMed]
- Cynis, H.; Hoffmann, T.; Friedrich, D.; Kehlen, A.; Gans, K.; Kleinschmidt, M.; Rahfeld, J.U.; Wolf, R.; Wermann, M.; Stephan, A.; et al. The isoenzyme of glutaminyl cyclase is an important regulator of monocyte infiltration under inflammatory conditions. EMBO Mol. Med. 2011, 3, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.H.; Clarklewis, I. Antagonists of monocyte chemoattractant protein-1 identified by modification of functionally critical NH2-terminal residues. J. Exp. Med. 1995, 181, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Hartlage-Rubsamen, M.; Waniek, A.; Meissner, J.; Morawski, M.; Schilling, S.; Jager, C.; Kleinschmidt, M.; Cynis, H.; Kehlen, A.; Arendt, T.; et al. Isoglutaminyl cyclase contributes to CCL2-driven neuroinflammation in Alzheimer’s disease. Acta Neuropathol. 2015, 129, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Barker, C.E.; Ali, S.; O’Boyle, G.; Kirby, J.A. Transplantation and inflammation: Implications for the modification of chemokine function. Immunology 2014, 143, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Proost, P.; Loos, T.; Mortier, A.; Schutyser, E.; Gouwy, M.; Noppen, S.; Dillen, C.; Ronsse, I.; Conings, R.; Struyf, S.; et al. Citrullination of CXCL8 by peptidylarginine deiminase alters receptor usage, prevents proteolysis, and dampens tissue inflammation. J. Exp. Med. 2008, 205, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Loos, T.; Opdenakker, G.; van Damme, J.; Proost, P. Citrullination of CXCL8 increases this chemokine’s ability to mobilize neutrophils into the blood circulation. Haematologica 2009, 94, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Mortier, A.; Loos, T.; Gouwy, M.; Ronsse, I.; van Damme, J.; Proost, P. Posttranslational modification of the NH2-terminal region of CXCL5 by proteases or peptidylarginine deiminases (PAD) differently affects its biological activity. J. Biol. Chem. 2010, 285, 29750–29759. [Google Scholar] [CrossRef] [PubMed]
- Loos, T.; Mortier, A.; Gouwy, M.; Ronsse, I.; Put, W.; Lenaerts, J.P.; van Damme, J.; Proost, P. Citrullination of CXCL10 and CXCL11 by peptidylarginine deiminase: A naturally occurring posttranslational modification of chemokines and new dimension of immunoregulation. Blood 2008, 112, 2648–2656. [Google Scholar] [CrossRef] [PubMed]
- Struyf, S.; Noppen, S.; Loos, T.; Mortier, A.; Gouwy, M.; Verbeke, H.; Huskens, D.; Luangsay, S.; Parmentier, M.; Geboes, K.; et al. Citrullination of CXCL12 differentially reduces CXCR4 and CXCR7 binding with loss of inflammatory and anti-HIV-1 activity via CXCR4. J. Immunol. 2009, 182, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Molon, B.; Ugel, S.; Del Pozzo, F.; Soldani, C.; Zilio, S.; Avella, D.; de Palma, A.; Mauri, P.; Monegal, A.; Rescigno, M.; et al. Chemokine nitration prevents intratumoral infiltration of antigen-specific T cells. J. Exp. Med. 2011, 208, 1949–1962. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.; Simpson, K.L.; Grisham, M.B.; Koyama, S.; Robbins, R.A. Effects of reactive oxygen and nitrogen metabolites on MCP-1-induced monocyte chemotactic activity in vitro. Am. J. Physiol. 1999, 277, L543–L549. [Google Scholar] [PubMed]
- Sato, E.; Simpson, K.L.; Grisham, M.B.; Koyama, S.; Robbins, R.A. Effects of reactive oxygen and nitrogen metabolites on RANTES- and IL-5-induced eosinophil chemotactic activity in vitro. Am. J. Pathol. 1999, 155, 591–598. [Google Scholar] [CrossRef]
- Pi, X.; Wu, Y.; Ferguson, J.E., 3rd; Portbury, A.L.; Patterson, C. SDF-1a stimulates JNK3 activity via enos-dependent nitrosylation of MKP7 to enhance endothelial migration. Proc. Natl. Acad. Sci. USA 2009, 106, 5675–5680. [Google Scholar] [CrossRef] [PubMed]
- Yasuoka, H.; Tsujimoto, M.; Yoshidome, K.; Nakahara, M.; Kodama, R.; Sanke, T.; Nakamura, Y. Cytoplasmic CXCR4 expression in breast cancer: Induction by nitric oxide and correlation with lymph node metastasis and poor prognosis. BMC Cancer 2008, 8, 340. [Google Scholar] [CrossRef] [PubMed]
- Huttenrauch, F.; Pollok-Kopp, B.; Oppermann, M. G protein-coupled receptor kinases promote phosphorylation and β-arrestin-mediated internalization of CCR5 homo- and hetero-oligomers. J. Biol. Chem. 2005, 280, 37503–37515. [Google Scholar] [CrossRef] [PubMed]
- Barker, B.L.; Benovic, J.L. G protein-coupled receptor kinase 5 phosphorylation of HIP regulates internalization of the chemokine receptor CXCR4. Biochemistry 2011, 50, 6933–6941. [Google Scholar] [CrossRef] [PubMed]
- Raghuwanshi, S.K.; Su, Y.; Singh, V.; Haynes, K.; Richmond, A.; Richardson, R.M. The chemokine receptors CXCR1 and CXCR2 couple to distinct G protein-coupled receptor kinases to mediate and regulate leukocyte functions. J. Immunol. 2012, 189, 2824–2832. [Google Scholar] [CrossRef] [PubMed]
- Pu, Q.; Yu, C. Glycosyltransferases, glycosylation and atherosclerosis. Glycoconj. J. 2014, 31, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Grodecka, M.; Czerwinski, M.; Duk, M.; Lisowska, E.; Wasniowska, K. Analysis of recombinant Duffy protein-linked N-glycans using lectins and glycosidases. Acta Biochim. Pol. 2010, 57, 49–53. [Google Scholar] [PubMed]
- Sarmiento, J.; Kypreos, K.E.; Prado, G.N.; Suetomi, K.; Stanzel, C.; Maxwell, C.; Shumate, D.; Tandang-Silvas, M.R.; Rajarathnam, K.; Navarro, J. Adenovirus mediated expression “in vivo” of the chemokine receptor CXCR1. J. Struct. Funct. Genomics 2009, 10, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Czerwinski, M.; Kern, J.; Grodecka, M.; Paprocka, M.; Krop-Watorek, A.; Wasniowska, K. Mutational analysis of the N-glycosylation sites of Duffy antigen/receptor for chemokines. Biochem. Biophys. Res. Commun. 2007, 356, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Margulies, B.J.; Gibson, W. The chemokine receptor homologue encoded by US27 of human cytomegalovirus is heavily glycosylated and is present in infected human foreskin fibroblasts and enveloped virus particles. Virus Res. 2007, 123, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Kremer, L.; Zaballos, A.; Goya, I.; Martinez, A.C.; Marquez, G. Analysis of post-translational CCR8 modifications and their influence on receptor activity. J. Biol. Chem. 2004, 279, 14726–14733. [Google Scholar] [CrossRef]
- Blackburn, P.E.; Simpson, C.V.; Nibbs, R.J.; O’Hara, M.; Booth, R.; Poulos, J.; Isaacs, N.W.; Graham, G.J. Purification and biochemical characterization of the D6 chemokine receptor. Biochem. J. 2004, 379, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Chabot, D.J.; Chen, H.; Dimitrov, D.S.; Broder, C.C. N-linked glycosylation of CXCR4 masks coreceptor function for CCR5-dependent human immunodeficiency virus type 1 isolates. J. Virol. 2000, 74, 4404–4413. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Craig, S.; Farzan, M.; Sogah, D.; Santo, N.V.; Choe, H.; Sodroski, J. Sialylated O-glycans and sulfated tyrosines in the NH2-terminal domain of CC chemokine receptor 5 contribute to high affinity binding of chemokines. J. Exp. Med. 2001, 194, 1661–1673. [Google Scholar] [CrossRef] [PubMed]
- Preobrazhensky, A.A.; Dragan, S.; Kawano, T.; Gavrilin, M.A.; Gulina, I.V.; Chakravarty, L.; Kolattukudy, P.E. Monocyte chemotactic protein-1 receptor CCR2B is a glycoprotein that has tyrosine sulfation in a conserved extracellular N-terminal region. J. Immunol. 2000, 165, 5295–5303. [Google Scholar] [CrossRef] [PubMed]
- Hauser, M.A.; Kindinger, I.; Laufer, J.M.; Spate, A.K.; Bucher, D.; Vanes, S.L.; Krueger, W.A.; Wittmann, V.; Legler, D.F. Distinct CCR7 glycosylation pattern shapes receptor signaling and endocytosis to modulate chemotactic responses. J. Leukoc. Biol. 2016, 99, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Kiermaier, E.; Moussion, C.; Veldkamp, C.T.; Gerardy-Schahn, R.; de Vries, I.; Williams, L.G.; Chaffee, G.R.; Phillips, A.J.; Freiberger, F.; Imre, R.; et al. Polysialylation controls dendritic cell trafficking by regulating chemokine recognition. Science 2016, 351, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Ehlert, J.E.; Flad, H.D.; Brandt, E. Identification of distinct surface-expressed and intracellular CXC-chemokine receptor 2 glycoforms in neutrophils: N-glycosylation is essential for maintenance of receptor surface expression. J. Immunol. 2000, 165, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.; Latzin, P.; Hordijk, P.; Marcos, V.; Rudolph, C.; Woischnik, M.; Krauss-Etschmann, S.; Koller, B.; Reinhardt, D.; Roscher, A.A.; et al. Cleavage of CXCR1 on neutrophils disables bacterial killing in cystic fibrosis lung disease. Nat. Med. 2007, 13, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Stone, M.J.; Chuang, S.; Hou, X.; Shoham, M.; Zhu, J.Z. Tyrosine sulfation: An increasingly recognised post-translational modification of secreted proteins. New Biotechnol. 2009, 25, 299–317. [Google Scholar] [CrossRef]
- Ludeman, J.P.; Stone, M.J. The structural role of receptor tyrosine sulfation in chemokine recognition. Brit. J. Pharmacol. 2014, 171, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Farzan, M.; Mirzabekov, T.; Kolchinsky, P.; Wyatt, R.; Cayabyab, M.; Gerard, N.P.; Gerard, C.; Sodroski, J.; Choe, H. Tyrosine sulfation of the amino terminus of CCR5 facilitates HIV-1 entry. Cell 1999, 96, 667–676. [Google Scholar] [CrossRef]
- Farzan, M.; Babcock, G.J.; Vasilieva, N.; Wright, P.L.; Kiprilov, E.; Mirzabekov, T.; Choe, H. The role of post-translational modifications of the CXCR4 amino terminus in stromal-derived factor 1a association and HIV-1 entry. J. Biol. Chem. 2002, 277, 29484–29489. [Google Scholar] [CrossRef] [PubMed]
- Stone, M.J.; Payne, R.J. Homogeneous sulfopeptides and sulfoproteins: Synthetic approaches and applications to characterize the effects of tyrosine sulfation on biochemical function. Acc. Chem. Res. 2015, 48, 2251–2261. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.S.; Zhu, J.Z.; Widlanski, T.S.; Stone, M.J. Regulation of chemokine recognition by site-specific tyrosine sulfation of receptor peptides. Chem. Biol. 2009, 16, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Millard, C.J.; Ludeman, J.P.; Simpson, L.S.; Clayton, D.J.; Payne, R.J.; Widlanski, T.S.; Stone, M.J. Tyrosine sulfation influences the chemokine binding selectivity of peptides derived from chemokine receptor CCR3. Biochemistry 2011, 50, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.H.Y.; Ludeman, J.P.; Wedderburn, J.; Canals, M.; Hall, P.; Butler, S.J.; Taleski, D.; Christopoulos, A.; Hickey, M.J.; Payne, R.J.; et al. Tyrosine sulfation of chemokine receptor CCR2 enhances interactions with both monomeric and dimeric forms of the chemokine monocyte chemoattractant protein-1 (MCP-1). J. Biol. Chem. 2013, 288, 10024–10034. [Google Scholar] [CrossRef] [PubMed]
- Veldkamp, C.T.; Seibert, C.; Peterson, F.C.; Sakmar, T.P.; Volkman, B.F. Recognition of a CXCR4 sulfotyrosine by the chemokine stromal cell-derived factor-1α (SDF-1α/CXCL12). J. Mol. Biol. 2006, 359, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Huma, Z.E.; Ludeman, J.P.; Wilkinson, B.L.; Payne, R.J.; Stone, M.J. NMR characterization of cooperativity: Fast ligand binding coupled to slow protein dimerization. Chem. Sci. 2014, 5, 2783–2788. [Google Scholar] [CrossRef]
- Rajarathnam, K.; Prado, G.N.; Fernando, H.; Clark-Lewis, I.; Navarro, J. Probing receptor binding activity of interleukin-8 dimer using a disulfide trap. Biochemistry 2006, 45, 7882–7888. [Google Scholar] [CrossRef] [PubMed]
- Nasser, M.W.; Raghuwanshi, S.K.; Grant, D.J.; Jala, V.R.; Rajarathnam, K.; Richardson, R.M. Differential activation and regulation of CXCR1 and CXCR2 by CXCL8 monomer and dimer. J. Immunol. 2009, 183, 3425–3432. [Google Scholar] [CrossRef] [PubMed]
- Drury, L.J.; Ziarek, J.J.; Gravel, S.; Veldkamp, C.T.; Takekoshi, T.; Hwang, S.T.; Heveker, N.; Volkman, B.F.; Dwinell, M.B. Monomeric and dimeric CXCL12 inhibit metastasis through distinct CXCR4 interactions and signaling pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 17655–17660. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, A.; Sawant, K.V.; Sarmiento, J.; Navarro, J.; Rajarathnam, K. Chemokine CXCL1 dimer is a potent agonist for the CXCR2 receptor. J. Biol. Chem. 2013, 288, 12244–12252. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, A.; Joseph, P.R.; Rajarathnam, K. Structural basis for differential binding of the interleukin-8 monomer and dimer to the CXCR1 N-domain: Role of coupled interactions and dynamics. Biochemistry 2009, 48, 8795–8805. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.H.Y.; Canals, M.; Ludeman, J.P.; Wedderburn, J.; Boston, C.; Butler, S.J.; Carrick, A.M.; Parody, T.R.; Taleski, D.; Christopoulos, A.; et al. Design and receptor interactions of obligate dimeric mutant of chemokine monocyte chemoattractant protein-1 (MCP-1). J. Biol. Chem. 2012, 287, 14692–14702. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Shen, X.; Baggett, B.R.; Kong, X.; LiWang, P.J. The human CC chemokine MIP-1b dimer is not competent to bind to the CCR5 receptor. J. Biol. Chem. 2007, 282, 27976–27983. [Google Scholar] [CrossRef] [PubMed]
- St. Charles, R.; Walz, D.A.; Edwards, B.F. The three-dimensional structure of bovine platelet factor 4 at 3.0-a resolution. J. Biol. Chem. 1989, 264, 2092–2099. [Google Scholar]
- Skelton, N.J.; Aspiras, F.; Ogez, J.; Schall, T.J. Proton NMR assignments and solution conformation of RANTES, a chemokine of the C-C type. Biochemistry 1995, 34, 5329–5342. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Watson, C.; Sharp, J.S.; Handel, T.M.; Prestegard, J.H. Oligomeric structure of the chemokine CCL5/RANTES from NMR, MS, and SAXS data. Structure 2011, 19, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; LiWang, P.J. Characterization of the interactions of vMIP-II, and a dimeric variant of vMIP-II, with glycosaminoglycans. Biochemistry 2010, 49, 7012–7022. [Google Scholar] [CrossRef] [PubMed]
- Salanga, C.L.; O’Hayre, M.; Handel, T. Modulation of chemokine receptor activity through dimerization and crosstalk. Cell. Mol. Life Sci. 2009, 66, 1370–1386. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Sweeney, M.D.; Saad, O.M.; Crown, S.E.; Hsu, A.R.; Handel, T.M.; Leary, J.A. Chemokine-glycosaminoglycan binding: Specificity for CCR2 ligand binding to highly sulfated oligosaccharides using FTICR mass spectrometry. J. Biol. Chem. 2005, 280, 32200–32208. [Google Scholar] [CrossRef] [PubMed]
- Nesmelova, I.V.; Sham, Y.; Dudek, A.Z.; van Eijk, L.I.; Wu, G.; Slungaard, A.; Mortari, F.; Griffioen, A.W.; Mayo, K.H. Platelet factor 4 and interleukin-8 CXC chemokine heterodimer formation modulates function at the quaternary structural level. J. Biol. Chem. 2005, 280, 4948–4958. [Google Scholar] [CrossRef] [PubMed]
- Nesmelova, I.V.; Sham, Y.; Gao, J.; Mayo, K.H. CXC and CC chemokines form mixed heterodimers: Association free energies from molecular dynamics simulations and experimental correlations. J. Biol. Chem. 2008, 283, 24155–24166. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Baxter, S.A.; Dreau, D.; Nesmelova, I.V. The heterodimerization of platelet-derived chemokines. Biochim. Biophys. Acta 2013, 1834, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Paavola, C.D.; Hemmerich, S.; Grunberger, D.; Polsky, I.; Bloom, A.; Freedman, R.; Mulkins, M.; Bhakta, S.; McCarley, D.; Wiesent, L.; et al. Monomeric monocyte chemoattractant protein-1 (MCP-1) binds and activates the MCP-1 receptor CCR2B. J. Biol. Chem. 1998, 273, 33157–33165. [Google Scholar] [CrossRef] [PubMed]
- Tuinstra, R.L.; Peterson, F.C.; Kutlesa, S.; Elgin, E.S.; Kron, M.A.; Volkman, B.F. Interconversion between two unrelated protein folds in the lymphotactin native state. Proc. Natl. Acad. Sci. USA 2008, 105, 5057–5062. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.C.; Tyler, R.C.; Guzzo, C.; Tuinstra, R.L.; Peterson, F.C.; Lusso, P.; Volkman, B.F. Engineering metamorphic chemokine lymphotactin/XCL1 into the GAG-binding, HIV-inhibitory dimer conformation. ACS Chem. Biol. 2015, 10, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Stephens, B.; Handel, T.M. Chemokine receptor oligomerization and allostery. Prog. Mol. Biol. Transl. Sci. 2013, 115, 375–420. [Google Scholar] [PubMed]
- Kraemer, S.; Alampour-Rajabi, S.; El Bounkari, O.; Bernhagen, J. Hetero-oligomerization of chemokine receptors: Diversity and relevance for function. Curr. Med. Chem. 2013, 20, 2524–2536. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.M.; Holgado, B.L.; Martinez, A.C.; Rodriguez-Frade, J.M.; Mellado, M. Chemokine receptor oligomerization: A further step toward chemokine function. Immunol. Lett. 2012, 145, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.M.; Lucas, P.; Holgado, B.L.; Barroso, R.; Vega, B.; Rodriguez-Frade, J.M.; Mellado, M. Receptor oligomerization: A pivotal mechanism for regulating chemokine function. Pharmacol. Ther. 2011, 131, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Hernanz-Falcon, P.; Rodriguez-Frade, J.M.; Serrano, A.; Juan, D.; del Sol, A.; Soriano, S.F.; Roncal, F.; Gomez, L.; Valencia, A.; Martinez, A.C.; et al. Identification of amino acid residues crucial for chemokine receptor dimerization. Nat. Immunol. 2004, 5, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Percherancier, Y.; Berchiche, Y.A.; Slight, I.; Volkmer-Engert, R.; Tamamura, H.; Fujii, N.; Bouvier, M.; Heveker, N. Bioluminescence resonance energy transfer reveals ligand-induced conformational changes in CXCR4 homo- and heterodimers. J. Biol. Chem. 2005, 280, 9895–9903. [Google Scholar] [CrossRef] [PubMed]
- El-Asmar, L.; Springael, J.Y.; Ballet, S.; Andrieu, E.U.; Vassart, G.; Parmentier, M. Evidence for negative binding cooperativity within CCR5-CCR2B heterodimers. Mol. Pharmacol. 2005, 67, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Rodriguez-Frade, J.M.; Vila-Coro, A.J.; Fernandez, S.; Martin de Ana, A.; Jones, D.R.; Toran, J.L.; Martinez, A.C. Chemokine receptor homo- or heterodimerization activates distinct signaling pathways. EMBO J. 2001, 20, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Chakera, A.; Seeber, R.M.; John, A.E.; Eidne, K.A.; Greaves, D.R. The duffy antigen/receptor for chemokines exists in an oligomeric form in living cells and functionally antagonizes CCR5 signaling through hetero-oligomerization. Mol. Pharmacol. 2008, 73, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.; See, H.B.; Seeber, R.M.; Armstrong, S.P.; White, C.W.; Ventura, S.; Ayoub, M.A.; Pfleger, K.D. Identification and profiling of novel α1a-adrenoceptor-CXC chemokine receptor 2 heteromer. J. Biol. Chem. 2012, 287, 12952–12965. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Chuang, L.F.; Yau, P.; Doi, R.H.; Chuang, R.Y. Interactions of opioid and chemokine receptors: Oligomerization of m, k, and d with CCR5 on immune cells. Exp. Cell Res. 2002, 280, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Pello, O.M.; Martinez-Munoz, L.; Parrillas, V.; Serrano, A.; Rodriguez-Frade, J.M.; Toro, M.J.; Lucas, P.; Monterrubio, M.; Martinez, A.C.; Mellado, M. Ligand stabilization of CXCR4/d-opioid receptor heterodimers reveals a mechanism for immune response regulation. Eur. J. Immunol. 2008, 38, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Parenty, G.; Appelbe, S.; Milligan, G. CXCR2 chemokine receptor antagonism enhances DOP opioid receptor function via allosteric regulation of the CXCR2-DOP receptor heterodimer. Biochem. J. 2008, 412, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, J.; Bot, G.; Szabo, I.; Rogers, T.J.; Liu-Chen, L.Y. Heterodimerization and cross-desensitization between the m-opioid receptor and the chemokine CCR5 receptor. Eur. J. Pharmacol. 2004, 483, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J. Historical review: A brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol. Sci. 2004, 25, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Hamm, H.E. The many faces of G protein signaling. J. Biol. Chem. 1998, 273, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, D.E.; Kurachi, Y.; Galper, J.; Neer, E.J.; Clapham, D.E. The b g subunits of GTP-binding proteins activate the muscarinic K+ channel in heart. Nature 1987, 325, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Violin, J.D.; Lefkowitz, R.J. b-arrestin-biased ligands at seven-transmembrane receptors. Trends Pharmacol. Sci. 2007, 28, 416–422. [Google Scholar] [CrossRef] [PubMed]
- DeWire, S.M.; Ahn, S.; Lefkowitz, R.J.; Shenoy, S.K. b-arrestins and cell signaling. Annu. Rev. Physiol. 2007, 69, 483–510. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J.; Shenoy, S.K. Transduction of receptor signals by b-arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.K.; Drake, M.T.; Nelson, C.D.; Houtz, D.A.; Xiao, K.; Madabushi, S.; Reiter, E.; Premont, R.T.; Lichtarge, O.; Lefkowitz, R.J. b-arrestin-dependent, G protein-independent ERK1/2 activation by the b2 adrenergic receptor. J. Biol. Chem. 2006, 281, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chu, J.; Qiu, Y.; Loh, H.H.; Law, P.Y. Agonist-selective signaling is determined by the receptor location within the membrane domains. Proc. Natl. Acad. Sci. USA 2008, 105, 9421–9426. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.; Fisher, R.A. Introduction: G protein-coupled receptors and RGS proteins. Prog. Mol. Biol. Transl. Sci. 2015, 133, 1–11. [Google Scholar] [PubMed]
- Berman, D.M.; Wilkie, T.M.; Gilman, A.G. GAIP and RGS4 are GTPase-activating proteins for the Gi subfamily of G protein a subunits. Cell 1996, 86, 445–452. [Google Scholar] [CrossRef]
- Ross, E.M.; Wilkie, T.M. GTPase-activating proteins for heterotrimeric G proteins: Regulators of G protein signaling (RGS) and RGS-like proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef] [PubMed]
- Chini, B.; Parenti, M. G-protein coupled receptors in lipid rafts and caveolae: How, when and why do they go there? J. Mol. Endocrinol. 2004, 32, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Escriba, P.V.; Wedegaertner, P.B.; Goni, F.M.; Vogler, O. Lipid-protein interactions in GPCR-associated signaling. Biochim. Biophys. Acta 2007, 1768, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, P.; Bardi, G.; Clark-Lewis, I.; Baggiolini, M.; Uguccioni, M. Eotaxin is a natural antagonist for CCR2 and an agonist for CCR5. Blood 2001, 97, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, R.; Sabroe, I.; LaRosa, G.; Williams, T.J.; Pease, J.E. The CC chemokine eotaxin (CCL11) is a partial agonist of CC chemokine receptor 2B. J. Biol. Chem. 2001, 276, 42957–42964. [Google Scholar] [CrossRef] [PubMed]
- Parody, T.R.; Stone, M.J. High level expression, activation, and antagonism of CC chemokine receptors CCR2 and CCR3 in Chinese hamster ovary cells. Cytokine 2004, 27, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Prazeres, D.M.; Martins, S.A. G protein-coupled receptors: An overview of signaling mechanisms and screening assays. Methods Mol. Biol. 2015, 1272, 3–19. [Google Scholar] [PubMed]
- Berchiche, Y.A.; Gravel, S.; Pelletier, M.E.; St-Onge, G.; Heveker, N. Different effects of the different natural CC chemokine receptor 2B ligands on b-arrestin recruitment, Gai signaling, and receptor internalization. Mol. Pharmacol. 2011, 79, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.D.; Clarke, W.P.; von Zastrow, M.; Nichols, D.E.; Kobilka, B.; Weinstein, H.; Javitch, J.A.; Roth, B.L.; Christopoulos, A.; Sexton, P.M.; et al. Functional selectivity and classical concepts of quantitative pharmacology. J Pharmacol Exp Ther 2007, 320, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shonberg, J.; Lopez, L.; Scammells, P.J.; Christopoulos, A.; Capuano, B.; Lane, J.R. Biased agonism at G protein-coupled receptors: The promise and the challenges—A medicinal chemistry perspective. Med. Res. Rev. 2014, 34, 1286–1330. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Rajagopal, K.; Lefkowitz, R.J. Teaching old receptors new tricks: Biasing seven-transmembrane receptors. Nat. Rev. Drug Discov. 2010, 9, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Kenakin, T.; Watson, C.; Muniz-Medina, V.; Christopoulos, A.; Novick, S. A simple method for quantifying functional selectivity and agonist bias. ACS Chem. Neurosci. 2012, 3, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Ahn, S.; Rominger, D.H.; Gowen-MacDonald, W.; Lam, C.M.; Dewire, S.M.; Violin, J.D.; Lefkowitz, R.J. Quantifying ligand bias at seven-transmembrane receptors. Mol. Pharmacol. 2011, 80, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Kenakin, T.; Christopoulos, A. Measurements of ligand bias and functional affinity. Nat. Rev. Drug Discov. 2013, 12, 483. [Google Scholar] [CrossRef] [PubMed]
- Kohout, T.A.; Nicholas, S.L.; Perry, S.J.; Reinhart, G.; Junger, S.; Struthers, R.S. Differential desensitization, receptor phosphorylation, b-arrestin recruitment, and ERK1/2 activation by the two endogenous ligands for the CC chemokine receptor 7. J. Biol. Chem. 2004, 279, 23214–23222. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Bassoni, D.L.; Campbell, J.J.; Gerard, N.P.; Gerard, C.; Wehrman, T.S. Biased agonism as a mechanism for differential signaling by chemokine receptors. J. Biol. Chem. 2013, 288, 35039–35048. [Google Scholar] [CrossRef] [PubMed]
- Corbisier, J.; Gales, C.; Huszagh, A.; Parmentier, M.; Springael, J.Y. Biased signaling at chemokine receptors. J. Biol. Chem. 2015, 290, 9542–9554. [Google Scholar] [CrossRef] [PubMed]
- Montaner, S.; Kufareva, I.; Abagyan, R.; Gutkind, S.J. Molecular mechanisms deployed by virally encoded G protein–coupled receptors in human diseases. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 331–354. [Google Scholar] [CrossRef] [PubMed]
- Dagna, L.; Lusso, P. Virus-encoded chemokines, chemokine receptors and chemokine-binding proteins: New paradigms for future therapy. Futur. Virol. 2007, 2, 353–368. [Google Scholar] [CrossRef]
- Vischer, H.F.; Siderius, M.; Leurs, R.; Smit, M.J. Herpesvirus-encoded GPCRs: Neglected players in inflammatory and proliferative diseases? Nat. Rev. Drug Discov. 2014, 13, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Sozzani, S.; Luini, W.; Bianchi, G.; Allavena, P.; Wells, T.N.C.; Napolitano, M.; Bernardini, G.; Vecchi, A.; D’Ambrosio, D.; Mazzeo, D.; et al. The viral chemokine macrophage inflammatory protein-II is a selective Th2 chemoattractant. Blood 1998, 92, 4036–4039. [Google Scholar] [PubMed]
- Gershengorn, M.C.; Geras-Raaka, E.; Varma, A.; Clark-Lewis, I. Chemokines activate Kaposi’s sarcoma-associated herpesvirus G protein-coupled receptor in mammalian cells in culture. J. Clin. Investig. 1998, 102, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Kledal, T.N.; Rosenkilde, M.M.; Coulin, F.; Simmons, G.; Johnsen, A.H.; Alouani, S.; Power, C.A.; Luttichau, H.R.; Gerstoft, J.; Clapham, P.R.; et al. A broad-spectrum chemokine antagonist encoded by Kaposi's sarcoma-associated herpesvirus. Science 1997, 277, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Nijmeijer, S.; Leurs, R.; Smit, M.J.; Vischer, H.F. The Epstein-Barr virus-encoded G protein-coupled receptor BILF1 hetero-oligomerizes with human CXCR4, scavenges Gai proteins, and constitutively impairs CXCR4 functioning. J. Biol. Chem. 2010, 285, 29632–29641. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Fallon, R.E.; Mangan, N.E.; Walsh, C.M.; Saraiva, M.; Sayers, J.R.; McKenzie, A.N.J.; Alcami, A.; Fallon, P.G. Schistosoma mansoni secretes a chemokine binding protein with anti-inflammatory activity. J. Exp. Med. 2005, 202, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Deruaz, M.; Frauenschuh, A.; Alessandri, A.L.; Dias, J.M.; Coelho, F.M.; Russo, R.C.; Ferreira, B.R.; Graham, G.J.; Shaw, J.P.; Wells, T.N.; et al. Ticks produce highly selective chemokine binding proteins with anti-inflammatory activity. J. Exp. Med. 2008, 205, 2019–2031. [Google Scholar] [CrossRef] [PubMed]
- Frauenschuh, A.; Power, C.A.; Deruaz, M.; Ferreira, B.R.; Silva, J.S.; Teixeira, M.M.; Dias, J.M.; Martin, T.; Wells, T.N.; Proudfoot, A.E. Molecular cloning and characterization of a highly selective chemokine-binding protein from the tick Rhipicephalus sanguineus. J. Biol. Chem. 2007, 282, 27250–27258. [Google Scholar] [CrossRef] [PubMed]
- Seet, B.T.; McFadden, G. Viral chemokine-binding proteins. J. Leukoc. Biol. 2002, 72, 24–34. [Google Scholar] [PubMed]
- Heidarieh, H.; Hernaez, B.; Alcami, A. Immune modulation by virus-encoded secreted chemokine binding proteins. Virus Res. 2015, 209, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E.; Bonvin, P.; Power, C.A. Targeting chemokines: Pathogens can, why can’t we? Cytokine 2015, 74, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.M.; Dairaghi, D.J.; Deitz, M.; Tsang, M.; Schall, T.J. Comprehensive mapping of poxvirus vCCI chemokine-binding protein: Expanded range of ligand interactions and unusual dissociation kinetics. J. Biol. Chem. 2002, 277, 2785–2789. [Google Scholar] [CrossRef] [PubMed]
- Carfí, A.; Smith, C.A.; Smolak, P.J.; McGrew, J.; Wiley, D.C. Structure of a soluble secreted chemokine inhibitor vCCI (p35) from cowpox virus. Proc. Natl. Acad. Sci. USA 1999, 96, 12379–12383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; DeRider, M.; McCornack, M.A.; Jao, S.; Isern, N.; Ness, T.; Moyer, R.; LiWang, P.J. Solution structure of the complex between poxvirus-encoded CC chemokine inhibitor vCCI and human MIP-1b. Proc. Natl. Acad. Sci. USA 2006, 103, 13985–13990. [Google Scholar] [CrossRef] [PubMed]
- Bahar, M.W.; Kenyon, J.C.; Putz, M.M.; Abrescia, N.G.; Pease, J.E.; Wise, E.L.; Stuart, D.I.; Smith, G.L.; Grimes, J.M. Structure and function of A41, a vaccinia virus chemokine binding protein. PLoS Path. 2008, 4, e5. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Lu, Q.; Wei, H.; Wang, D.; Chen, D.; He, G.; Huang, L.; Wang, H.; Wang, X. Structural basis of chemokine sequestration by CrmD, a poxvirus-encoded tumor necrosis factor receptor. PLoS Path. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.M.; Nelson, C.A.; van Berkel, V.; Lau, E.K.; Studts, J.M.; Brett, T.J.; Speck, S.H.; Handel, T.M.; Virgin, H.W.; Fremont, D.H. Structural basis of chemokine sequestration by a herpesvirus decoy receptor. Cell 2002, 111, 343–356. [Google Scholar] [CrossRef]
- Wang, D.; Bresnahan, W.; Shenk, T. Human cytomegalovirus encodes a highly specific RANTES decoy receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 16642–16647. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.M.; Losberger, C.; Deruaz, M.; Power, C.A.; Proudfoot, A.E.; Shaw, J.P. Structural basis of chemokine sequestration by a tick chemokine binding protein: The crystal structure of the complex between evasin-1 and CCL3. PLoS ONE 2009, 4, e8514. [Google Scholar] [CrossRef] [PubMed]
- Pease, J.; Horuk, R. Chemokine receptor antagonists. J. Med. Chem. 2012, 55, 9363–9392. [Google Scholar] [CrossRef] [PubMed]
- Scholten, D.J.; Canals, M.; Maussang, D.; Roumen, L.; Smit, M.J.; Wijtmans, M.; de Graaf, C.; Vischer, H.F.; Leurs, R. Pharmacological modulation of chemokine receptor function. Br. J. Pharmacol. 2012, 165, 1617–1643. [Google Scholar] [CrossRef] [PubMed]
- Dorr, P.; Westby, M.; Dobbs, S.; Griffin, P.; Irvine, B.; Macartney, M.; Mori, J.; Rickett, G.; Smith-Burchnell, C.; Napier, C.; et al. Maraviroc (UK-427,857), a potent, orally bioavailable, and selective small-molecule inhibitor of chemokine receptor CCR5 with broad-spectrum anti-human immunodeficiency virus type 1 activity. Antimicrob. Agents Chemother. 2005, 49, 4721–4732. [Google Scholar] [CrossRef] [PubMed]
- Saita, Y.; Kodama, E.; Orita, M.; Kondo, M.; Miyazaki, T.; Sudo, K.; Kajiwara, K.; Matsuoka, M.; Shimizu, Y. Structural basis for the interaction of CCR5 with a small molecule, functionally selective CCR5 agonist. J. Immunol. 2006, 177, 3116–3122. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.W.; Liu, Y.; Getschman, A.E.; Peterson, F.C.; Ziarek, J.J.; Li, R.; Volkman, B.F.; Chen, Y. Structural analysis of a novel small molecule ligand bound to the CXCL12 chemokine. J. Med. Chem. 2014, 57, 9693–9699. [Google Scholar] [CrossRef] [PubMed]
- Mao-Yu, Z.; Jin-Jian, L.; Wang, L.; Zi-Chao, G.; Hu, H.; Carolina Oi Lam, U.; Yi-Tao, W. Development of monoclonal antibodies in China: Overview and prospects. BioMed Res. Int. 2015, 2015, 168935. [Google Scholar] [CrossRef]
- Klarenbeek, A.; Maussang, D.; Blanchetot, C.; Saunders, M.; van der Woning, S.; Smit, M.; de Haard, H.; Hofman, E. Targeting chemokines and chemokine receptors with antibodies. Drug Discov. Today Technol. 2012, 9, e237–e244. [Google Scholar] [CrossRef] [PubMed]
- Duvic, M.; Evans, M.; Wang, C. Mogamulizumab for the treatment of cutaneous T-cell lymphoma: Recent advances and clinical potential. Therap. Adv. Hematol. 2016, 7, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.; Dolezal, O.; Cao, B.; Nilsson, S.K.; See, H.B.; Pfleger, K.D.G.; Roche, M.; Gorry, P.R.; Pow, A.; Viduka, K.; et al. I-bodies, human single domain antibodies that antagonize chemokine receptor CXCR4. J. Biol. Chem. 2016, 291, 12641–12657. [Google Scholar] [CrossRef] [PubMed]
- Oberthür, D.; Achenbach, J.; Gabdulkhakov, A.; Buchner, K.; Maasch, C.; Falke, S.; Rehders, D.; Klussmann, S.; Betzel, C. Crystal structure of a mirror-image L-RNA aptamer (Spiegelmer) in complex with the natural L-protein target CCL2. Nat. Commun. 2015, 6, 6923. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stone, M.J.; Hayward, J.A.; Huang, C.; E. Huma, Z.; Sanchez, J. Mechanisms of Regulation of the Chemokine-Receptor Network. Int. J. Mol. Sci. 2017, 18, 342. https://doi.org/10.3390/ijms18020342
Stone MJ, Hayward JA, Huang C, E. Huma Z, Sanchez J. Mechanisms of Regulation of the Chemokine-Receptor Network. International Journal of Molecular Sciences. 2017; 18(2):342. https://doi.org/10.3390/ijms18020342
Chicago/Turabian StyleStone, Martin J., Jenni A. Hayward, Cheng Huang, Zil E. Huma, and Julie Sanchez. 2017. "Mechanisms of Regulation of the Chemokine-Receptor Network" International Journal of Molecular Sciences 18, no. 2: 342. https://doi.org/10.3390/ijms18020342
APA StyleStone, M. J., Hayward, J. A., Huang, C., E. Huma, Z., & Sanchez, J. (2017). Mechanisms of Regulation of the Chemokine-Receptor Network. International Journal of Molecular Sciences, 18(2), 342. https://doi.org/10.3390/ijms18020342