
Developmental Programming of Adult Disease: Reprogramming by Melatonin?




Abstract
:1. Introduction
2. Mechanisms of Developmental Programming
2.1. Insight Provided by Human Studies
2.2. Insight Provided by Animal Models
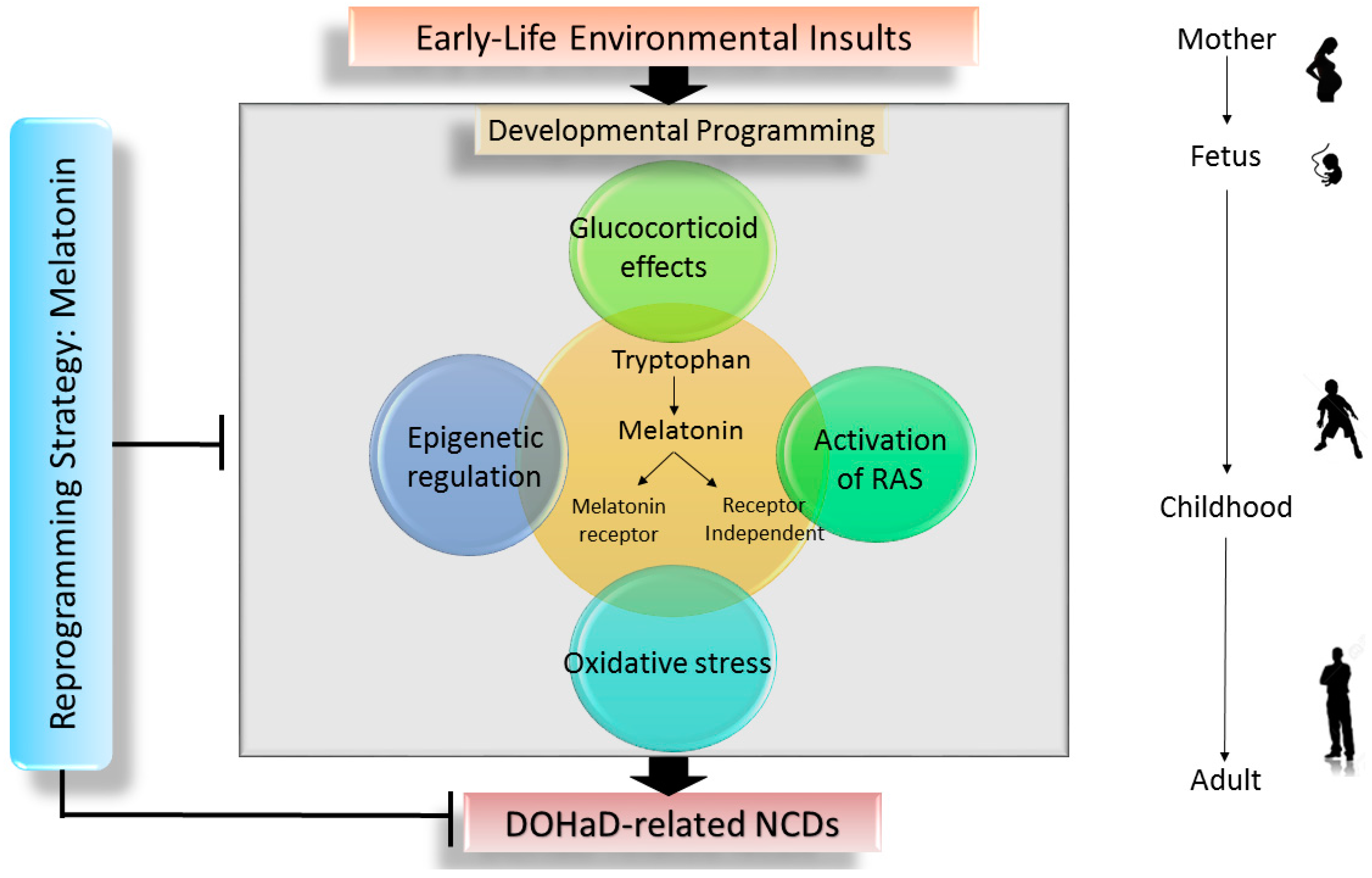
2.3 The Interrelationship between Melatonin and Underlying Mechanisms of Developmental Programming
3. The Impact of Melatonin on Pregnancy and Fetal Development
3.1. Synthesis, Metabolism, and Signaling Pathway of Melatonin
3.2. Melatonin in Pregnancy and Fetus
4. Melatonin as a Reprogramming Therapy in Animal Models of Developmental Programming
5. Long-Term Effects of Melatonin in Normal Offspring
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CVD | Cardiovascular disease |
| DNMT | DNA methyltransferases |
| DOHaD | Developmental origins of health and disease |
| HDAC | Histone deacetylase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| l-NAME | NG-nitro-l-arginine-methyester |
| MT | Melatonin receptor |
| NCD | Non-communicable disease |
| NGS | Next generation RNA sequencing |
| NOS | Nitric oxide synthase |
| RAS | Renin-angiotensin system |
| ROS | Reactive oxygen species |
References
- Zarocostas, J. Need to increase focus on non-communicable diseases in global health, says WHO. BMJ 2010, 341, c7065. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.; Gluckman, P. Developmental origins of noncommunicable disease: Population and public health implications. Am. J. Clin. Nutr. 2011, 94, 1754S–1758S. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A. Programming by early nutrition: An experimental approach. J. Nutr. 1998, 128, 401S–406S. [Google Scholar] [PubMed]
- Tain, Y.L.; Joles, J.A. Reprogramming: A preventive strategy in hypertension focusing on the kidney. Int. J. Mol. Sci. 2015, 17, E23. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Reiter, R.J. Epigenetic regulation: A new research area for melatonin? J. Pineal Res. 2008, 44, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Sheen, J.M.; Tiao, M.M.; Tain, Y.L.; Huang, L.T. Roles of melatonin in fetal programming in compromised pregnancies. Int. J. Mol. Sci. 2013, 14, 5380–5401. [Google Scholar] [CrossRef] [PubMed]
- Voiculescu, S.E.; Zygouropoulos, N.; Zahiu, C.D.; Zagrean, A.M. Role of melatonin in embryo fetal development. J. Med. Life 2014, 7, 488–492. [Google Scholar] [PubMed]
- Tain, Y.L.; Huang, L.T.; Chan, J.Y. Transcriptional regulation of programmed hypertension by melatonin: An epigenetic perspective. Int. J. Mol. Sci. 2014, 15, 18484–18495. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Roseboom, T.; de Rooij, S.; Painter, R. The Dutch famine and its long-term consequences for adult health. Early Hum. Dev. 2006, 82, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Burgner, D.P.; Ponsonby, A.L.; Saffery, R.; Huang, R.C.; Vuillermin, P.J.; Cheung, M.; Craig, J.M. Effects of early-life environment and epigenetics on cardiovascular disease risk in children: Highlighting the role of twin studies. Pediatr. Res. 2013, 73, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Vågerö, D.; Leon, D. Ischaemic heart disease and low birth weight: A test of the fetal-origins hypothesis from the Swedish Twin Registry. Lancet 1994, 343, 260–263. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736. [Google Scholar] [CrossRef] [PubMed]
- Cianfarani, S.; Germani, D.; Branca, F. Low birthweight and adult insulin resistance: The “catch-up growth” hypothesis. Arch. Dis. Child. Fetal Neonatal. 1999, 81, F71–F73. [Google Scholar] [CrossRef]
- Chavatte-Palmer, P.; Tarrade, A.; Rousseau-Ralliard, D. Diet before and during Pregnancy and Offspring Health: The Importance of Animal Models and What Can Be Learned from Them. Int. J. Environ. Res. Public Health 2016, 13, E586. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Williams, L.; Vuguin, P.M.; Charron, M.J. Minireview: Epigenetic programming of diabetes and obesity: Animal models. Endocrinology 2012, 153, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Rinaudo, P.; Wang, E. Fetal programming and metabolic syndrome. Annu. Rev. Physiol. 2012, 74, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Rabadán-Diehl, C.; Nathanielsz, P. From Mice to Men: Research models of developmental programming. J. Dev. Orig. Health Dis. 2013, 4, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.T.; Dasinger, J.H.; Intapad, S. Fetal programming and cardiovascular pathology. Compr. Physiol. 2015, 5, 997–1025. [Google Scholar] [PubMed]
- Walker, C.L.; Ho, S.M. Developmental reprogramming of cancer susceptibility. Nat. Rev. Cancer 2012, 12, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Krauss-Etschmann, S.; Meyer, K.F.; Dehmel, S.; Hylkema, M.N. Inter- and transgenerational epigenetic inheritance: Evidence in asthma and COPD? Clin. Epigenet. 2015, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Bogdarina, I.; Welham, S.; King, P.J.; Burns, S.P.; Clark, A.J. Epigenetic modification of the renin-angiotensin system in the fetal programming of hypertension. Circ. Res. 2007, 100, 520–526. [Google Scholar] [CrossRef] [PubMed]
- McMullen, S.; Langley-Evans, S.C.; Gambling, L.; Lang, C.; Swali, A.; McArdle, H.J. A common cause for a common phenotype: The gatekeeper hypothesis in fetal programming. Med. Hypotheses 2012, 78, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.P.; Al-Hasan, Y. Impact of oxidative stress in fetal programming. J. Pregnancy 2012, 2012, 582748. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, E.C.; Seckl, J.R. Prenatal stress, glucocorticoids and the programming of adult disease. Front. Behav. Neurosci. 2009, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Forrestel, A.C.; Miedlich, S.U.; Yurcheshen, M.; Wittlin, S.D.; Sellix, M.T. Chronomedicine and type 2 diabetes: Shining some light on melatonin. Diabetologia 2016. [Google Scholar] [CrossRef] [PubMed]
- Haus, E.L.; Smolensky, M.H. Shift work and cancer risk: Potential mechanistic roles of circadian disruption, light at night, and sleep deprivation. Sleep Med. Rev. 2013, 17, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol. 2007, 54, 1–9. [Google Scholar] [PubMed]
- Pozo, D.; Reiter, R.J.; Calvo, J.R.; Guerrero, J.M. Physiological concentrations of melatonin inhibit nitric oxide synthase in rat cerebellum. Life Sci. 1994, 55, 455–460. [Google Scholar] [CrossRef]
- Escames, G.; López, L.C.; Tapias, V.; Utrilla, P.; Reiter, R.J.; Hitos, A.B.; León, J.; Rodríguez, M.I.; Acuña-Castroviejo, D. Melatonin counteracts inducible mitochondrial nitric oxide synthase-dependent mitochondrial dysfunction in skeletal muscle of septic mice. J. Pineal Res. 2006, 40, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.H.; Kuo, H.C.; Lin, I.C.; Chien, S.J.; Huang, L.T.; Tain, Y.L. Melatonin prevents neonatal dexamethasone induced programmed hypertension: Histone deacetylase inhibition. J. Steroid Biochem. Mol. Biol. 2014, 144, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Haldar, C. Physiological crosstalk between melatonin and glucocorticoid receptor modulates T-cell mediated immune responses in a wild tropical rodent, Funambulus pennanti. J. Steroid Biochem. Mol. Biol. 2013, 134, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Tain, Y.L. Postnatal dexamethasone-induced programmed hypertension is related to the regulation of melatonin and its receptors. Steroids 2016, 108, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tiao, M.M.; Huang, L.T.; Chen, C.J.; Sheen, J.M.; Tain, Y.L.; Chen, C.C.; Kuo, H.C.; Huang, Y.H.; Tang, K.S.; Chu, E.W.; Yu, H.R. Melatonin in the regulation of liver steatosis following prenatal glucocorticoid exposure. Biomed. Res. Int. 2014, 2014, 942172. [Google Scholar] [CrossRef] [PubMed]
- Lui, C.C.; Hsu, M.H.; Kuo, H.C.; Chen, C.C.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Tain, Y.L.; Chang, K.A.; Huang, L.T. Effects of melatonin on prenatal dexamethasone-induced epigenetic alterations in hippocampal morphology and reelin and glutamic acid decarboxylase 67 levels. Dev. Neurosci. 2015, 37, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Chen, C.C.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Melatonin attenuates prenatal dexamethasone-induced blood pressure increase in a rat model. J. Am. Soc. Hypertens. 2014, 8, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Sheen, J.M.; Yu, H.R.; Chen, C.C.; Tiao, M.M.; Hsu, C.N.; Lin, Y.J.; Kuo, K.C.; Huang, L.T. Maternal Melatonin Therapy Rescues Prenatal Dexamethasone and Postnatal High-Fat Diet Induced Programmed Hypertension in Male Rat Offspring. Front. Physiol. 2015, 6, 377. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Reiter, R.J.; Pechanova, O.; Paulis, L. Experimental models of melatonin-deficient hypertension. Front. Biosci. 2013, 18, 616–625. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.T. Melatonin therapy prevents programmed hypertension and nitric oxide deficiency in offspring exposed to maternal caloric restriction. Oxid. Med. Cell Longev. 2014, 2014, 283180. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal N(G)-Nitro-l-arginine-methyl ester (l-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obs. Gynecol. 2016, 215, 636. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Nakamura, Y.; Terron, M.P.; Flores, L.J.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and pregnancy in the human. Reprod. Toxicol. 2008, 25, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and reproduction revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Waddell, B.J.; Wharfe, M.D.; Crew, R.C.; Mark, P.J. A rhythmic placenta? Circadian variation, clock genes and placental function. Placenta 2012, 33, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Lanoix, D.; Guérin, P.; Vaillancourt, C. Placental melatonin production and melatonin receptor expression are altered in preeclampsia: New insights into the role of this hormone in pregnancy. J. Pineal Res. 2012, 53, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Serón-Ferré, M.; Mendez, N.; Abarzua-Catalan, L.; Vilches, N.; Valenzuela, F.J.; Reynolds, H.E.; Llanos, A.J.; Rojas, A.; Valenzuela, G.J.; Torres-Farfan, C. Circadian rhythms in the fetus. Mol. Cell Endocrinol. 2012, 349, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Simonneaux, V. Naughty melatonin: How mothers tick off their fetus. Endocrinology 2011, 152, 1734–1738. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, D.; Achten, C.; Dallmann, F.; Oster, H. Embryonic development and maternal regulation of murine circadian clock function. Chronobiol. Int. 2015, 32, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Matsuzuka, T.; Sakamoto, N.; Ozawa, M.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Alleviation of maternal hyperthermia-induced early embryonic death by administration of melatonin to mice. J. Pineal Res. 2005, 39, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Domínguez Rubio, A.P.; Sordelli, M.S.; Salazar, A.I.; Aisemberg, J.; Bariani, M.V.; Cella, M.; Rosenstein, R.E.; Franchi, A.M. Melatonin prevents experimental preterm labor and increases offspring survival. J. Pineal Res. 2014, 56, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Mendez, N.; Abarzua-Catalan, L.; Vilches, N.; Galdames, H.A.; Spichiger, C.; Richter, H.G.; Valenzuela, G.J.; Seron-Ferre, M.; Torres-Farfan, C. Timed maternal melatonin treatment reverses circadian disruption of the fetal adrenal clock imposed by exposure to constant light. PLoS ONE 2012, 7, e42713. [Google Scholar] [CrossRef] [PubMed]
- Torres-Farfan, C.; Valenzuela, F.J.; Mondaca, M.; Valenzuela, G.J.; Krause, B.; Herrera, E.A.; Riquelme, R.; Llanos, A.J.; Seron-Ferre, M. Evidence of a role for melatonin in fetal sheep physiology: Direct actions of melatonin on fetal cerebral artery, brown adipose tissue and adrenal gland. J. Physiol. 2008, 586, 4017–4027. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Aversa, S.; Reiter, R.J.; Barberi, I.; Pellegrino, S. Update on the use of melatonin in pediatrics. J. Pineal Res. 2011, 50, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Tain, Y.L.; Sheen, J.M.; Huang, L.T. Melatonin utility in neonates and children. J. Formos. Med. Assoc. 2012, 111, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Carpentieri, A.; Díaz de Barboza, G.; Areco, V.; Peralta López, M.; Tolosa de Talamoni, N. New perspectives in melatonin uses. Pharmacol. Res. 2012, 65, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Paulis, L. Melatonin as a potential antihypertensive treatment. J. Pineal Res. 2007, 42, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Braam, W.; Smits, M.G.; Didden, R.; Korzilius, H.; Van Geijlswijk, I.M.; Curfs, L.M. Exogenous melatonin for sleep problems in individuals with intellectual disability: A meta-analysis. Dev. Med. Child Neurol. 2009, 51, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; Dhandapany, P.S.; Brown, G.M.; Cardinali, D.P. Melatonin and human cardiovascular disease. J. Cardiovasc. Pharmacol. Ther. 2017, 22, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Opie, L.H.; Lecour, S. Melatonin has multiorgan effects. Eur. Heart J. Cardiovasc. Pharmacother. 2016, 2, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Leu, S.; Wu, K.L.; Lee, W.C.; Chan, J.Y. Melatonin prevents maternal fructose intake-induced programmed hypertension in the offspring: Roles of nitric oxide and arachidonic acid metabolites. J. Pineal Res. 2014, 57, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Dubovický, M.; Ujházy, E.; Kovacovský, P.; Navarová, J.; Juráni, M.; Soltés, L. Effect of melatonin on neurobehavioral dysfunctions induced by intrauterine hypoxia in rats. Cent. Eur. J. Public Health 2004, 12, S23–S25. [Google Scholar] [PubMed]
- Cisternas, C.D.; Compagnucci, M.V.; Conti, N.R.; Ponce, R.H.; Vermouth, N.T. Protective effect of maternal prenatal melatonin administration on rat pups born to mothers submitted to constant light during gestation. Braz. J. Med. Biol. Res. 2010, 43, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Baxi, D.B.; Singh, P.K.; Vachhrajani, K.D.; Ramachandran, A.V. Neonatal corticosterone programs for thrifty phenotype adult diabetic manifestations and oxidative stress: Countering effect of melatonin as a deprogrammer. J. Matern. Fetal Neonatal Med. 2012, 25, 1574–1585. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.P.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2016, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.J.; Keah, L.S.; Kumar, A.; Sirajudeen, K.N. Adverse effects of melatonin on rat pups of Wistar-Kyoto dams receiving melatonin supplementation during pregnancy. Exp. Toxicol. Pathol. 2012, 64, 751–752. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, S.A.; Akhondzadeh, S.; Mohammadi, M.R.; Keshtkar, A.A.; Hosseini, S.; Eshraghian, M.R.; Ahmadi, F.; Alipour, R.; Keshavarz, S.A. Role of melatonin in body weight: A systematic review and meta-analysis. Curr. Pharm. Des. 2016. [Google Scholar] [CrossRef]
- Chan, W.Y.; Ng, T.B. Development of pre-implantation mouse embryos under the influence of pineal indoles. J. Neural Transm. Gen. Sect. 1994, 96, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, G.; Marr, M.; Myers, C.; Wilson, R.; Travlos, G.; Price, C. Maternal and developmental toxicity evaluation of melatonin administered orally to pregnant Sprague-Dawley rats. Toxicol. Sci. 1999, 50, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Kleber, A.; Ruf, C.G.; Wolf, A.; Fink, T.; Glas, M.; Wolf, B.; Volk, T.; Abend, M.; Mathes, A.M. Melatonin or ramelteon therapy differentially affects hepatic gene expression profiles after haemorrhagic shock in rat--A microarray analysis. Exp. Mol. Pathol. 2015, 99, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Singhal, N.K.; Chauhan, A.K.; Jain, S.K.; Shanker, R.; Singh, C.; Singh, M.P. Silymarin- and melatonin-mediated changes in the expression of selected genes in pesticides-induced Parkinsonism. Mol. Cell. Biochem. 2013, 384, 47–58. [Google Scholar] [CrossRef] [PubMed]
- NIH DAVID Bioinformatics Resources 6.8. Available online: https://david.ncifcrf.gov/ (access on 9 January 2017).


{kind=link}
| Programming Models | Melatonin Treatment | Reprogramming Effects | Age at Evaluation | Ref. |
|---|---|---|---|---|
| 50% caloric restriction during pregnancy and lactation | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Increased renal NO | 3 mo | [42] |
| l-NAME 60 mg/kg/day subcutaneously during pregnancy | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Increased renal NO | 3 mo | [43] |
| 60% high fructose intake during pregnancy and lactation | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Increased renal NO | 3 mo | [63] |
| Phenytoin 50 mg/kg orally from gestational day 7 to 18 | Melatonin (40 μg/mL) in drinking water from gestational day 0 to 19 | Protected neurobehavioral dysfunctions | 3 mo | [64] |
| Neonatal dexamethasone exposure | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Preserved histone deacetylase gene expression | 4 mo | [33] |
| Neonatal dexamethasone exposure | 0.01% melatonin in drinking water during lactation | Prevented hypertension, Preserved renal melatonin receptor-2 protein, Increased renal melatonin level | 4 mo | [36] |
| Prenatal dexamethasone exposure | 0.01% melatonin in drinking water during pregnancy and lactation | Reversed methylation of leptin, Decreased liver steatosis | 4 mo | [37] |
| Prenatal dexamethasone exposure | 0.01% melatonin in drinking water during pregnancy and lactation | Reversed hippocampal morphology and reelin level | 4 mo | [38] |
| Prenatal dexamethasone exposure | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Increased nephron number | 4 mo | [39] |
| Prenatal dexamethasone exposure plus post-weaning high-fat diet | 0.01% melatonin in drinking water during pregnancy and lactation | Prevented hypertension, Upregulated Agtr1b and Mas1 expression | 4 mo | [40] |
| Corticosterone 1 µg/day in the morning from postnatal day 2 to 14 | Melatonin 40 μg/day at night from postnatal day 2 to 14 | Protected diabetic manifestations and oxidative stress | 4 mo | [65] |
| Constant light exposure from gestational day 10 to 21 | Melatonin 1 mg/kg at circadian time 12, from day 17 to 21 of pregnancy | Protected anxiety-like and sexual behaviors | 4 mo | [66] |
| Term | Count | % | p-Value | Benjamini |
|---|---|---|---|---|
| Focal adhesion | 50 | 2.3 | 4.4 × 10−8 | 1.2 × 10−5 |
| Regulation of actin cytoskeleton | 49 | 2.3 | 3.9 × 10−7 | 5.6 × 10−5 |
| Pathways in cancer | 75 | 3.5 | 4.8 × 10−7 | 4.5 × 10−5 |
| Axon guidance | 32 | 1.5 | 6.0 × 10−6 | 4.3 × 10−4 |
| ErbB signaling pathway | 25 | 1.2 | 1.5 × 10−5 | 8.3 × 10−4 |
| AMPK signaling pathway | 30 | 1.4 | 3.9 × 10−5 | 1.9 × 10−3 |
| Metabolic pathways | 177 | 8.2 | 6.7 × 10−5 | 2.7 × 10−3 |
| Chemokine signaling pathway | 37 | 1.7 | 6.8 × 10−5 | 2.4 × 10−3 |
| Insulin signaling pathway | 31 | 1.4 | 1.0 × 10−4 | 3.2 × 10−3 |
| Proteoglycans in cancer | 40 | 1.8 | 1.0 × 10−4 | 2.9 × 10−3 |
| PI3K-Akt signaling pathway | 59 | 2.7 | 1.1 × 10−4 | 2.7 × 10−3 |
| Ras signaling pathway | 44 | 2.0 | 1.2 × 10−4 | 2.9 × 10−3 |
| Adherens junction | 20 | 0.9 | 1.4 × 10−4 | 3.0 × 10−3 |
| PPAR signaling pathway | 20 | 0.9 | 2.9 × 10−4 | 5.9 × 10−3 |
| Butanoate metabolism | 11 | 0.5 | 3.5 × 10−4 | 6.6 × 10−3 |
| Renal cell carcinoma | 18 | 0.8 | 4.2 × 10−4 | 7.4 × 10−3 |
| MAPK signaling pathway | 46 | 2.1 | 4.4 × 10−4 | 7.3 × 10−3 |
| mTOR signaling pathway | 17 | 0.8 | 5.0 × 10−4 | 7.9 × 10−3 |
| Prostate cancer | 21 | 1.0 | 6.4 × 10−4 | 9.6 × 10−3 |
| Tryptophan metabolism | 14 | 0.6 | 6.5 × 10−4 | 9.3 × 10−3 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tain, Y.-L.; Huang, L.-T.; Hsu, C.-N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. https://doi.org/10.3390/ijms18020426
Tain Y-L, Huang L-T, Hsu C-N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? International Journal of Molecular Sciences. 2017; 18(2):426. https://doi.org/10.3390/ijms18020426
Chicago/Turabian StyleTain, You-Lin, Li-Tung Huang, and Chien-Ning Hsu. 2017. "Developmental Programming of Adult Disease: Reprogramming by Melatonin?" International Journal of Molecular Sciences 18, no. 2: 426. https://doi.org/10.3390/ijms18020426
APA StyleTain, Y. -L., Huang, L. -T., & Hsu, C. -N. (2017). Developmental Programming of Adult Disease: Reprogramming by Melatonin? International Journal of Molecular Sciences, 18(2), 426. https://doi.org/10.3390/ijms18020426