
Overexpression of Transforming Acidic Coiled Coil‑Containing Protein 3 Reflects Malignant Characteristics and Poor Prognosis of Glioma

Abstract
:1. Introduction
2. Results
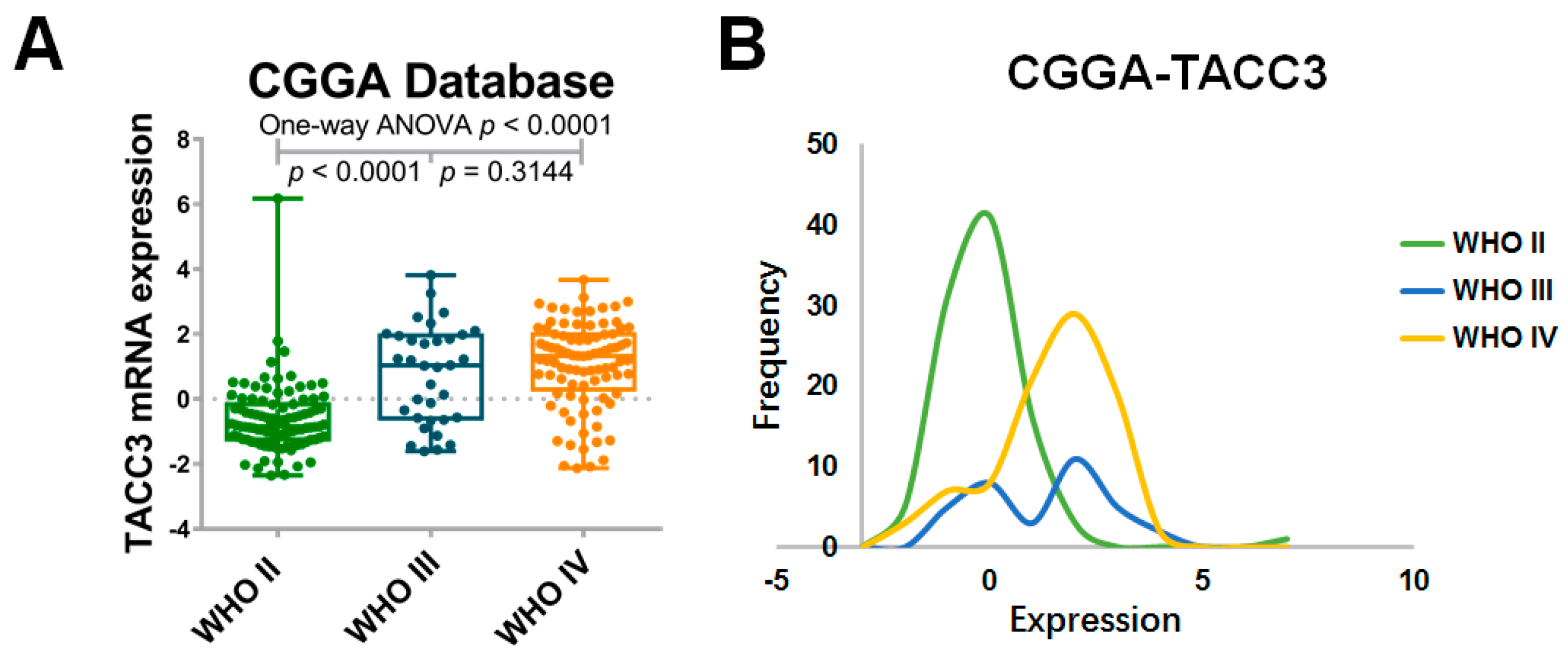
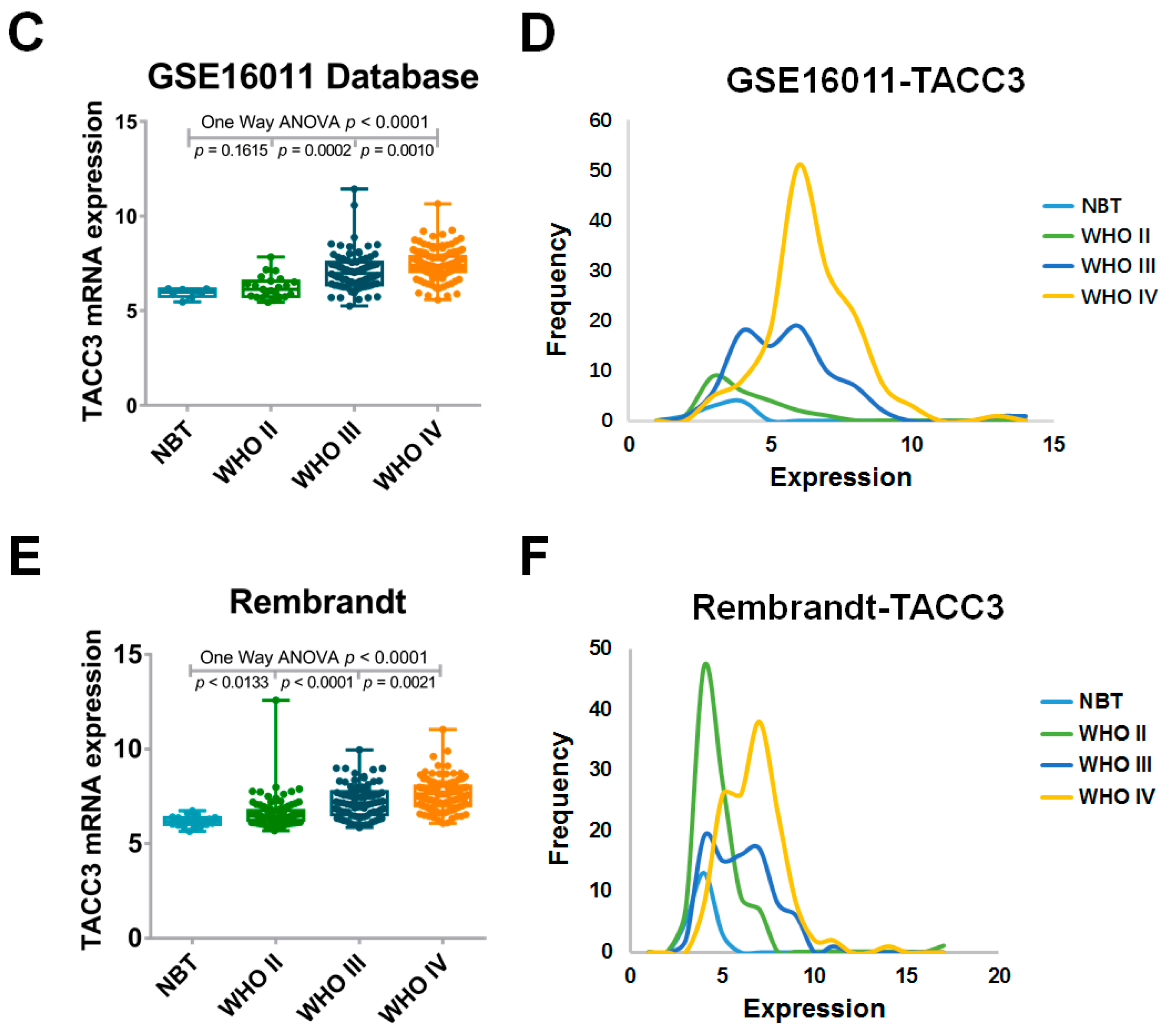
2.1. High Expression of Transforming Acidic Coiled-Coil-Containing Protein 3 (TACC3) Strongly Correlates with Glioma Grade
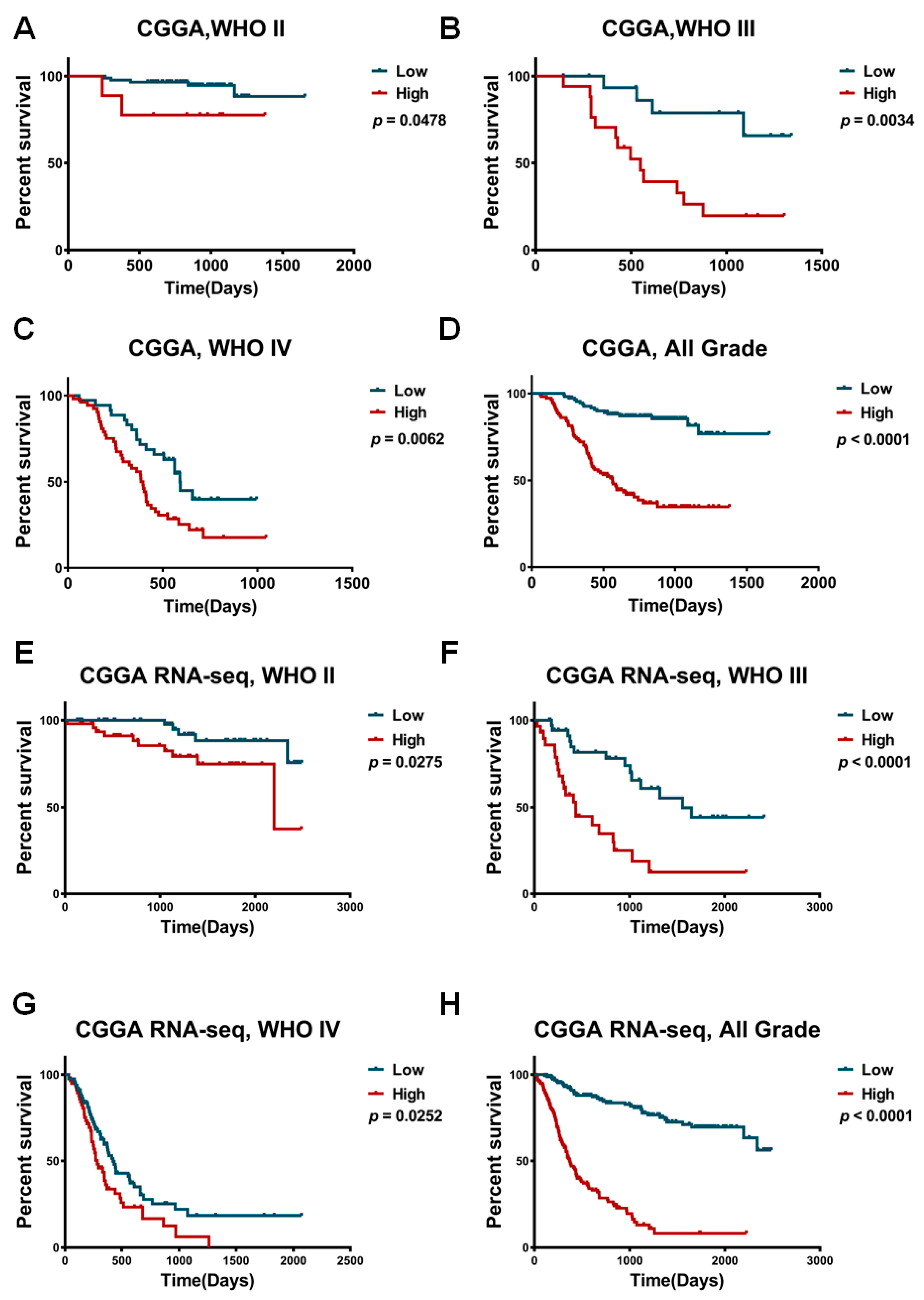
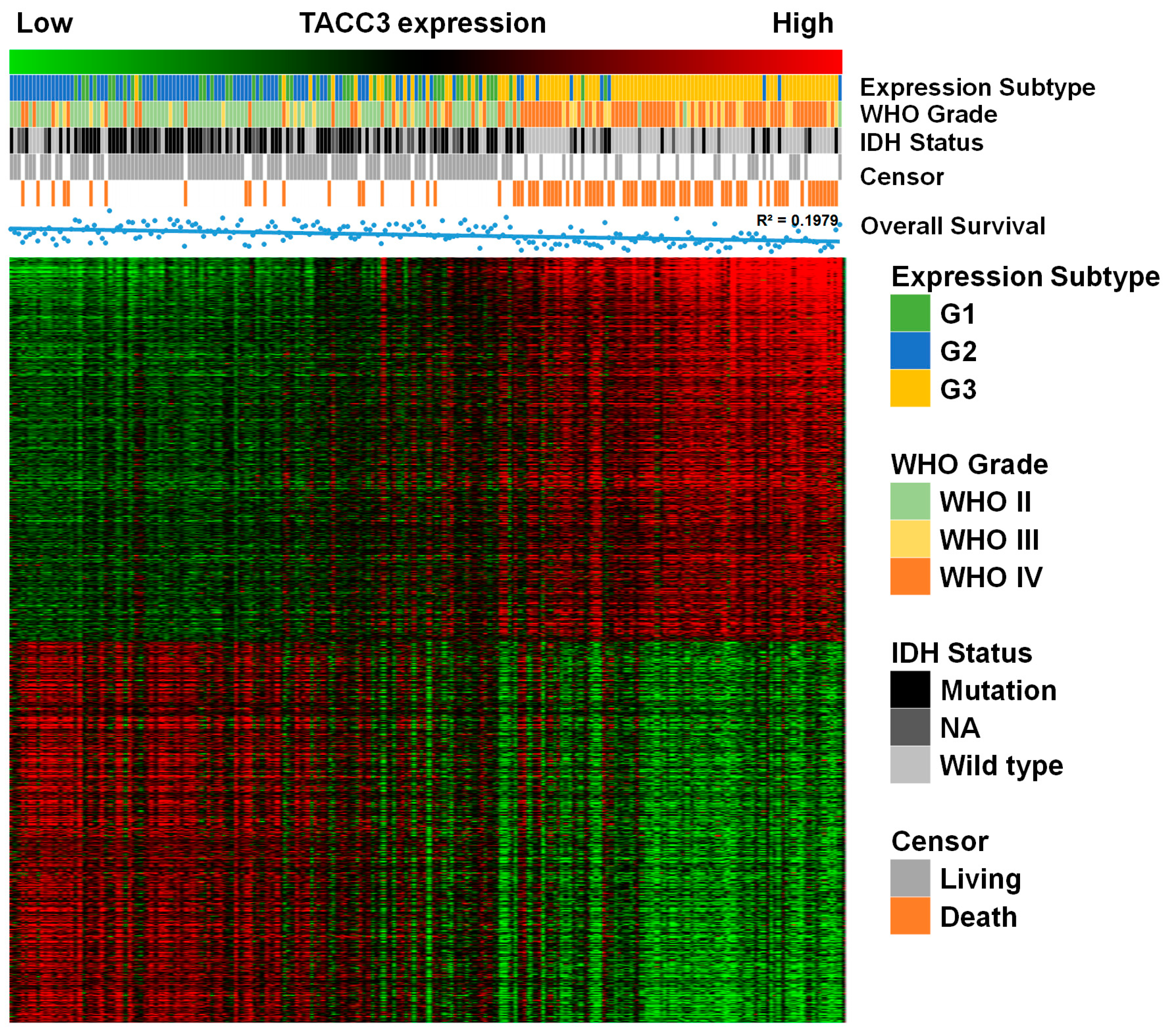
2.2. TACC3 as an Independent Prognostic Marker in Gliomas
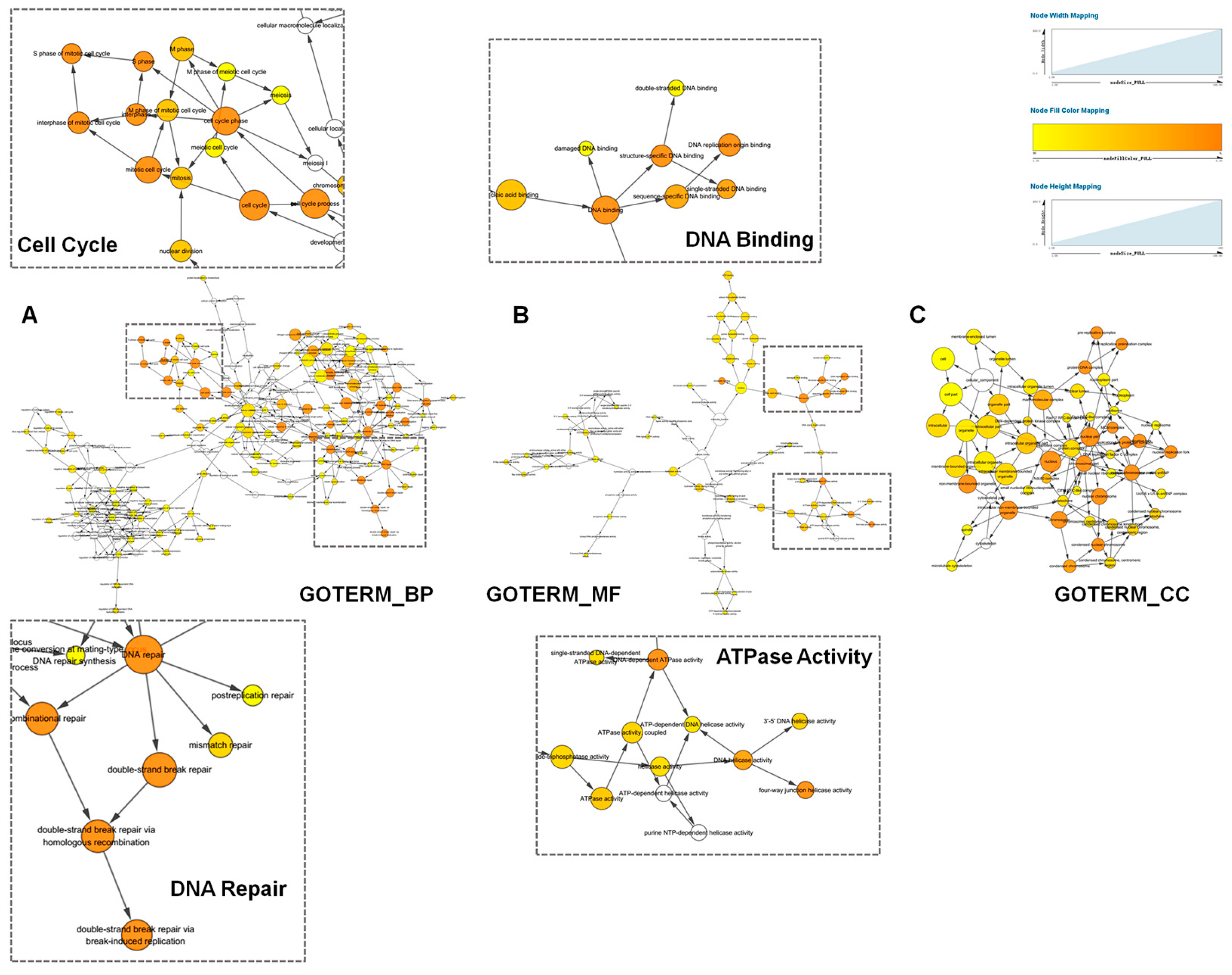
2.3. Functional Annotation of TACC3 Positive Correlated Genes
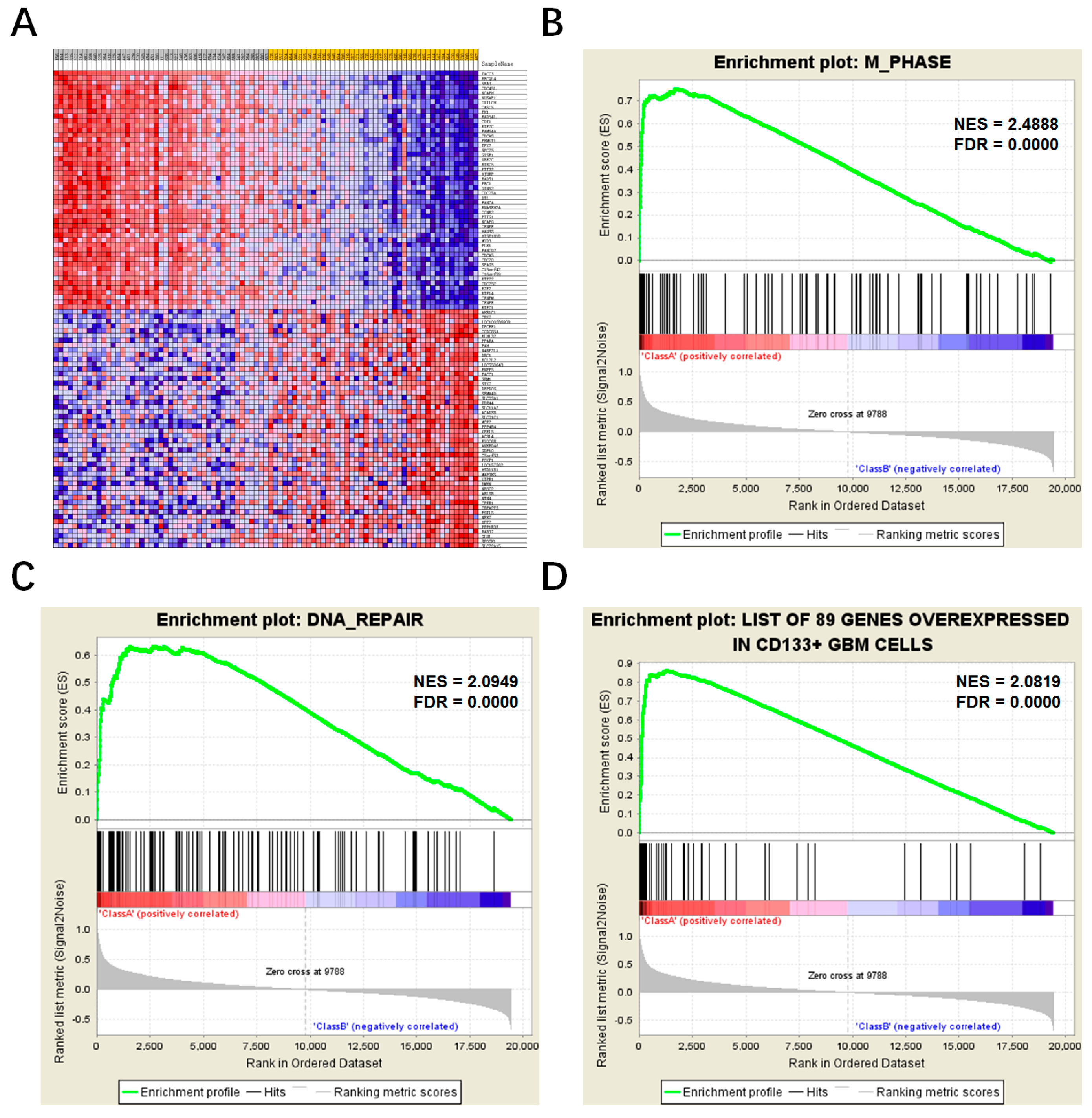
2.4. Comparison of Different TACC3 Expression Groups by Gene Set Enrichment Analysis (GSEA)
2.5. TACC3 Could Serve as an Indicator for Therapy Efficiency
2.6. TACC3-miRNA-mRNA Network
3. Discussion
4. Materials and Methods
4.1. Microarray and RNA-Sequencing Data
4.2. Functional Annotation and Gene Set Enrichment Analysis
4.3. Integrative Analysis of TACC3-miRNA-mRNA Interactions
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CGGA | Chinese Glioma Genome Atlas |
| TCGA | The Cancer Genome Atlas |
| GBM | Glioblastoma |
| OS | Overall survival |
| TACC3 | Transforming acidic coiled coil‑containing protein 3 |
| IDH | Isocitrate dehydrogenase |
| EMT | Epithelium-mesenchymal transition |
| HR | Hazards ratio |
| 95% CI | 95% confidence interval |
| KPS | Karnofsky performance status |
| NES | Normalized enrichment score |
| GSCs | Glioma stem cells |
| GSEA | Gene set enrichment analysis |
| FDR | False discovery rate |
References
- Shi, L.; Cheng, Z.; Zhang, J.; Li, R.; Zhao, P.; Fu, Z.; You, Y. hsa-mir-181a and hsa-mir-181b function as tumor suppressors in human glioma cells. Brain Res. 2008, 1236, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhu, W.; Shi, D.; Lv, L.; Zhang, C.; Liu, P.; Hu, W. MicroRNA-181a sensitizes human malignant glioma U87MG cells to radiation by targeting Bcl-2. Oncol. Rep. 2010, 23, 997–1003. [Google Scholar] [PubMed]
- Wang, H.; Tao, T.; Yan, W.; Feng, Y.; Wang, Y.; Cai, J.; You, Y.; Jiang, T.; Jiang, C. Upregulation of miR-181s reverses mesenchymal transition by targeting KPNA4 in glioblastoma. Sci. Rep. 2015, 5, 13072. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Mao, Y.; Ma, W.; Mao, Q.; You, Y.; Yang, X.; Jiang, C.; Kang, C.; Li, X.; Chen, L.; et al. CGCG clinical practice guidelines for the management of adult diffuse gliomas. Cancer Lett. 2016, 375, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Gergely, F.; Karlsson, C.; Still, I.; Cowell, J.; Kilmartin, J.; Raff, J.W. The TACC domain identifies a family of centrosomal proteins that can interact with microtubules. Proc. Natl. Acad. Sci. USA 2000, 97, 14352–14357. [Google Scholar] [CrossRef] [PubMed]
- Ha, G.H.; Kim, J.L.; Breuer, E.K. Transforming acidic coiled-coil proteins (TACCs) in human cancer. Cancer Lett. 2013, 336, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Ha, G.H.; Kim, J.L.; Breuer, E.K. TACC3 is essential for EGF-mediated EMT in cervical cancer. PLoS ONE 2013, 8, e70353. [Google Scholar] [CrossRef]
- Yun, M.; Rong, J.; Lin, Z.R.; He, Y.L.; Zhang, J.X.; Peng, Z.W.; Tang, L.Q.; Zeng, M.S.; Zhong, Q.; Ye, S. High expression of transforming acidic coiled coil-containing protein 3 strongly correlates with aggressive characteristics and poor prognosis of gastric cancer. Oncol. Rep. 2015, 34, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Ha, G.H.; Park, J.S.; Breuer, E.K. TACC3 promotes epithelial-mesenchymal transition (EMT) through the activation of PI3K/Akt and ERK signaling pathways. Cancer Lett. 2013, 332, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.G.; Kudla, D.M.; Deloia, J.A.; Chu, T.J.; Fairfull, L.; Edwards, R.P.; Ferrell, R.E. Comparative gene expression analysis of ovarian carcinoma and normal ovarian epithelium by serial analysis of gene expression. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1717–1723. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Liu, C.; Shen, N.; Yi, P.; Dong, F.; Li, X.; Zhang, N.; Huang, T. Overexpression of TACC3 in breast cancer associates with poor prognosis. Appl. Immunohistochem. Mol. Morphol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.L.; Lin, Z.R.; Xiao, Y.R.; Cao, X.; Zhu, L.C.; Zeng, M.S.; Zhong, Q.; Wen, Z.S. High expression of TACC3 in esophageal squamous cell carcinoma correlates with poor prognosis. Oncotarget 2015, 6, 6850–6861. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Chan, J.M.; Zoppoli, P.; Niola, F.; Sullivan, R.; Castano, A.; Liu, E.M.; Reichel, J.; Porrati, P.; Pellegatta, S.; et al. Transforming fusions of FGFR and TACC genes in human glioblastoma. Science 2012, 337, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature 2014, 507, 315–322. [Google Scholar]
- Capelletti, M.; Dodge, M.E.; Ercan, D.; Hammerman, P.S.; Park, S.I.; Kim, J.; Sasaki, H.; Jablons, D.M.; Lipson, D.; Young, L.; et al. Identification of recurrent FGFR3-TACC3 fusion oncogenes from lung adenocarcinoma. Clin. Cancer Res. 2014, 20, 6551–6558. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bao, S.; Wu, Q.; Wang, H.; Eyler, C.; Sathornsumetee, S.; Shi, Q.; Cao, Y.; Lathia, J.; McLendon, R.E.; et al. Hypoxia-inducible factors regulate tumorigenic capacity of glioma stem cells. Cancer Cell 2009, 15, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Ma, L.; Yi, D.; Yoon, J.G.; Diercks, A.; Foltz, G.; Price, N.D.; Hood, L.E.; Tian, Q. A CD133-related gene expression signature identifies an aggressive glioblastoma subtype with excessive mutations. Proc. Natl. Acad. Sci. USA 2011, 108, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Ying, H.; Yan, H.; Kimmelman, A.C.; Hiller, D.J.; Chen, A.J.; Perry, S.R.; Tonon, G.; Chu, G.C.; Ding, Z.; et al. p53 and Pten control neural and glioma stem/progenitor cell renewal and differentiation. Nature 2008, 455, 1129–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrió, D.; Rodriguez-Pinilla, S.M.; Hardisson, D.; Cano, A.; Moreno-Bueno, G.; Palacios, J. Epithelial-mesenchymal transition in breast cancer relates to the basal-like phenotype. Cancer Res. 2008, 68, 989–997. [Google Scholar]
- Ray, D.; Terao, Y.; Fuhrken, P.G.; Ma, Z.Q.; DeMayo, F.J.; Christov, K.; Heerema, N.A.; Franks, R.; Tsai, S.Y.; Papoutsakis, E.T.; et al. Deregulated CDC25A expression promotes mammary tumorigenesis with genomic instability. Cancer Res. 2007, 67, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kessler, P.; Williams, B.R. Transcript profiling of Wilms tumors reveals connections to kidney morphogenesis and expression patterns associated with anaplasia. Oncogene 2005, 24, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Johansson, F.K.; Göransson, H.; Westermark, B. Expression analysis of genes involved in brain tumor progression driven by retroviral insertional mutagenesis in mice. Oncogene 2005, 24, 3896–3905. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. Large-scale meta-analysis of cancer microarray data identifies common transcriptional profiles of neoplastic transformation and progression. Proc. Natl. Acad. Sci. USA 2004, 101, 9309–9314. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Goh, C.H.; Ooi, L.L.; Hui, K.M. Identification of unique and common low abundance tumour-specific transcripts by suppression subtractive hybridization and oligonucleotide probe array analysis. Oncogene 2008, 27, 4128–4136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, L.; Rodriguez-Aguayo, C.; Yuan, Y.; Debeb, B.G.; Chen, D.; Sun, Y.; You, M.J.; Liu, Y.; Dean, D.C.; et al. miR-205 acts as a tumour radiosensitizer by targeting ZEB1 and Ubc13. Nat. Commun. 2014, 5, 5671. [Google Scholar] [CrossRef] [PubMed]
- Sadik, H.; Korangath, P.; Nguyen, N.K.; Győrffy, B.; Kumar, R.; Hedayati, M.; Teo, W.W.; Park, S.; Panday, H.; Gonzalez, M.T.; et al. HOXC10 expression supports the development of chemotherapy resistance by fine tuning DNA repair in breast cancer cells. Cancer Res. 2016, 76, 4443–4456. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Li, R.; Wang, Y.Y.; Shi, Y.; Luan, W.K.; Tao, T.; Zhang, J.X.; Xu, Y.C.; You, Y.P. miR-1224–5p acts as a tumor suppressor by targeting CREB1 in malignant gliomas. Mol. Cell. Biochem. 2015, 403, 33–41. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, F.; Callari, M.; Dugo, M.; Merlino, G.; Musella, V.; Miodini, P.; Paolini, B.; Cappelletti, V.; Daidone, M.G. miR-30e* is an independent subtype-specific prognostic marker in breast cancer. Br. J. Cancer 2015, 113, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lu, G.; Ke, X.; Lu, X.; Wang, X.; Li, H.; Ren, M.; He, S. miR-488 acts as a tumor suppressor gene in gastric cancer. Tumour Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Denoyelle, C.; Lambert, B.; Meryet-Figuière, M.; Vigneron, N.; Brotin, E.; Lecerf, C.; Abeilard, E.; Giffard, F.; Louis, M.H.; Gauduchon, P.; et al. miR-491–5p-induced apoptosis in ovarian carcinoma depends on the direct inhibition of both BCL-XL and EGFR leading to BIM activation. Cell Death Dis. 2014, 5, e1445. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Liang, Y.; Wang, D.; Luo, F. MiR-940 inhibits hepatocellular carcinoma growth and correlates with prognosis of hepatocellular carcinoma patients. Cancer Sci. 2015, 106, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Qiu, H.; Cui, H.; Ni, X.; Li, J.; Liao, W.; Lu, L.; Ding, K. MicroRNA-885–3p inhibits the growth of HT-29 colon cancer cell xenografts by disrupting angiogenesis via targeting BMPR1A and blocking BMP/Smad/Id1 signaling. Oncogene 2015, 34, 1968–1978. [Google Scholar] [CrossRef] [PubMed]
- Yim, E.K.; Tong, S.Y.; Ho, E.M.; Bae, J.H.; Um, S.J.; Park, J.S. Anticancer effects on TACC3 by treatment of paclitaxel in HPV-18 positive cervical carcinoma cells. Oncol. Rep. 2009, 21, 549–557. [Google Scholar] [PubMed]
- Zhao, Q.; Kho, A.; Kenney, A.M.; Yuk, D.D.I.; Kohane, I.; Rowitch, D.H. Identification of genes expressed with temporal-spatial restriction to developing cerebellar neuron precursors by a functional genomic approach. Proc. Natl. Acad. Sci. USA 2002, 99, 5704–5709. [Google Scholar] [CrossRef] [PubMed]
- Cordenonsi, M.; Zanconato, F.; Azzolin, L.; Forcato, M.; Rosato, A.; Frasson, C.; Inui, M.; Montagner, M.; Parenti, A.R.; Poletti, A.; et al. The Hippo transducer TAZ confers cancer stem cell-related traits on breast cancer cells. Cell 2011, 147, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Basu-Roy, U.; Bayin, N.S.; Rattanakorn, K.; Han, E.; Placantonakis, D.G.; Mansukhani, A.; Basilico, C. Sox2 antagonizes the Hippo pathway to maintain stemness in cancer cells. Nat Commun. 2015. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Liu, X.; Chen, L.; Dou, Z.; Lei, X.; Chang, L.; Cai, J.; Cui, Y.; Yang, D.; Sun, Y.; et al. Targeting the SMO oncogene by miR-326 inhibits glioma biological behaviors and stemness. Neuro Oncol. 2015, 17, 243–253. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Subramanian, R.; Bangs, F.; Omelchenko, T.; Liem, K.F., Jr.; Kapoor, T.M.; Anderson, K.V. The kinesin-4 protein Kif7 regulates mammalian Hedgehog signalling by organizing the cilium tip compartment. Nat. Cell Biol. 2014, 16, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, M.; Lee, M.S.; Kim, C.H.; Lim, D.S. The MST1/2-SAV1 complex of the Hippo pathway promotes ciliogenesis. Nat. Commun. 2014, 5, 5370. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, J.; Qian, X.; Han, L.; Zhang, K.; Chen, L.; Liu, J.; Ren, Y.; Yang, M.; Zhang, A.; et al. AC1MMYR2, an inhibitor of dicer-mediated biogenesis of Oncomir miR-21, reverses epithelial-mesenchymal transition and suppresses tumor growth and progression. Cancer Res. 2013, 73, 5519–5531. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhang, W.; You, G.; Zhang, J.; Han, L.; Bao, Z.; Wang, Y.; Liu, Y.; Jiang, C.; Kang, C.; et al. Molecular classification of gliomas based on whole genome gene expression: A systematic report of 225 samples from the Chinese Glioma Cooperative Group. Neuro Oncol. 2012, 14, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhang, W.; Yang, P.; Wang, Y.; Li, M.; Zhang, C.; Wang, Z.; Hu, H.; Liu, Y.; Li, Q.; et al. Identification of a 6-cytokine prognostic signature in patients with primary glioblastoma harboring M2 microglia/macrophage phenotype relevance. PLoS ONE 2015, 10, e0126022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.B.; Zhu, P.; Yang, P.; Cai, J.Q.; Wang, Z.L.; Li, Q.B.; Bao, Z.S.; Zhang, W.; Jiang, T. Identification of high risk anaplastic gliomas by a diagnostic and prognostic signature derived from mRNA expression profiling. Oncotarget 2015, 6, 36643–36651. [Google Scholar] [PubMed]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed]
- Dweep, H.; Gretz, N. miRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods 2015, 12, 697. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Univariate | Multivariate | ||
|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | |
| Gender | ||||
| Male | 1.059 (0.683–1.642) | 0.799 | ||
| Female | ||||
| Age (years) | ||||
| <45 | 0.526 (0.339–0.814) | 0.004 | 0.795 (0.499–1.266) | 0.333 |
| ≥45 | ||||
| WHO Grade | ||||
| LGG | 0.068 (0.031–0.149) | <0.001 | 0.170 (0.069–0.419) | <0.001 |
| HGG | ||||
| KPS | ||||
| ≥80 | 0.194 (0.125–0.304) | <0.001 | 0.279 (0.171–0.457) | <0.001 |
| <80 | ||||
| IDH1 status | ||||
| Mutation | 0.244 (0.143–0.416) | <0.001 | 0.799 (0.437–1.460) | 0.465 |
| Wild-type | ||||
| TACC3 expression | ||||
| Low | 0.169 (0.099–0.290) | <0.001 | 0.402 (0.212–0.763) | 0.005 |
| High | ||||
| Variable | TACC3-Low (n = 110) | TACC3-High (n = 110) | p-Value | |
|---|---|---|---|---|
| Median age | 38 | 45 | ||
| Age | ≥45 | 36 | 56 | <0.01 |
| <45 | 74 | 54 | ||
| Gender | Male | 56 | 68 | >0.05 |
| Female | 54 | 42 | ||
| Preoperative KPS score | ≥80 | 95 | 79 | <0.01 |
| <80 | 15 | 31 | ||
| Grade | WHO II | 79 | 18 | <0.01 |
| WHO III | 13 | 21 | ||
| WHO IV | 18 | 71 | ||
| IDH1 status | Mutation | 60 | 26 | <0.01 |
| Wild type | 29 | 72 | ||
| NA | 21 | 12 | ||
| CGGA subtype | G1 | 29 | 13 | <0.01 |
| G2 | 70 | 12 | ||
| G3 | 11 | 95 | ||
| TCGA subtype | Classical | 12 | 26 | <0.01 |
| Mesenchymal | 14 | 62 | ||
| Neural | 50 | 5 | ||
| Proneural | 34 | 17 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Tian, Y.; Wang, G.-Z.; Zhao, S.-H.; Han, B.; Li, Y.-L.; Jiang, C.-L. Overexpression of Transforming Acidic Coiled Coil‑Containing Protein 3 Reflects Malignant Characteristics and Poor Prognosis of Glioma. Int. J. Mol. Sci. 2017, 18, 235. https://doi.org/10.3390/ijms18030235
Sun Y, Tian Y, Wang G-Z, Zhao S-H, Han B, Li Y-L, Jiang C-L. Overexpression of Transforming Acidic Coiled Coil‑Containing Protein 3 Reflects Malignant Characteristics and Poor Prognosis of Glioma. International Journal of Molecular Sciences. 2017; 18(3):235. https://doi.org/10.3390/ijms18030235
Chicago/Turabian StyleSun, Ying, Yu Tian, Guang-Zhi Wang, Shi-Hong Zhao, Bo Han, Yong-Li Li, and Chuan-Lu Jiang. 2017. "Overexpression of Transforming Acidic Coiled Coil‑Containing Protein 3 Reflects Malignant Characteristics and Poor Prognosis of Glioma" International Journal of Molecular Sciences 18, no. 3: 235. https://doi.org/10.3390/ijms18030235
APA StyleSun, Y., Tian, Y., Wang, G. -Z., Zhao, S. -H., Han, B., Li, Y. -L., & Jiang, C. -L. (2017). Overexpression of Transforming Acidic Coiled Coil‑Containing Protein 3 Reflects Malignant Characteristics and Poor Prognosis of Glioma. International Journal of Molecular Sciences, 18(3), 235. https://doi.org/10.3390/ijms18030235