
Cellular Reprogramming Using Protein and Cell-Penetrating Peptides


Abstract
:1. Introduction
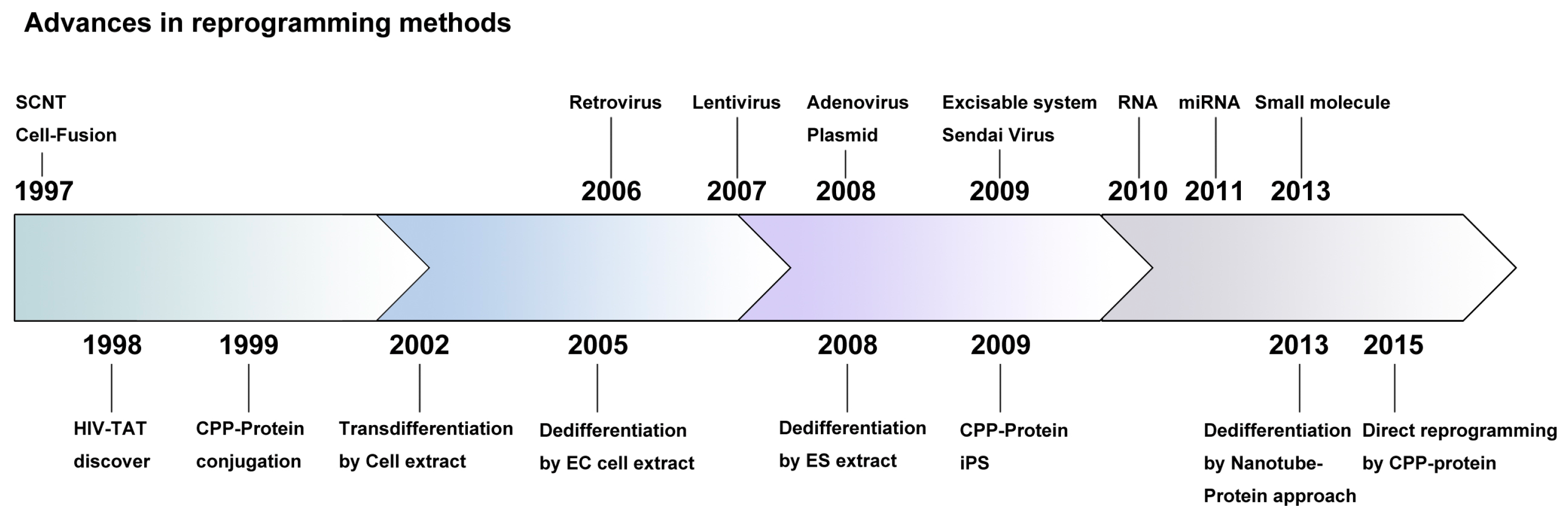
2. History of Exogene-Free Induced Pluripotent Stem Cells
3. Protein-Based Reprogramming Approaches
4. Development of Protein Transduction Technology
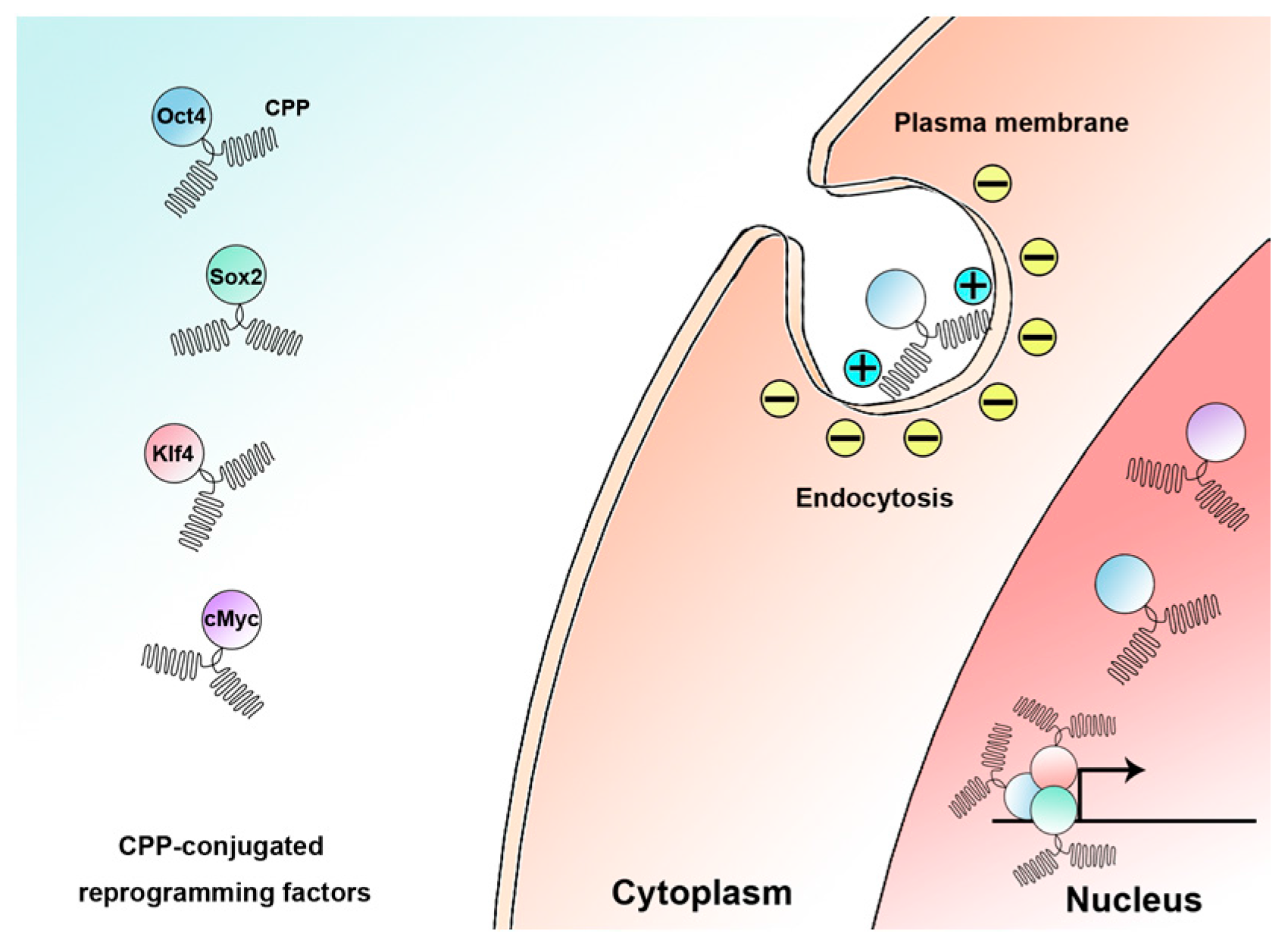
5. Reprogramming via Cell-Penetrating Peptide-Mediated Protein Transduction
5.1. Groundbreaking Success in Reprogramming by Using Cell-Penetrating Peptides
5.2. Efforts to Increase Cell-Penetrating Efficiency Using the Other Peptides
5.3. Cell-Penetrating Peptide-Based Direct Lineage Conversion
5.4. Various Approaches for Protein Transduction in Reprogramming
6. Limitations of Cell-Penetrating Peptide-Mediated Reprogramming
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CPP | Cell-penetrating peptide |
| PTD | Protein transduction domain |
| SCNT | Somatic cell nuclear transfer |
| MII | Metaphase II |
| iPSCs | Induced pluripotent stem cells |
| ESCs | Embryonic stem cells |
| SeV | Sendai virus |
| EC | Embryonic carcinoma |
| HIV-TAT | Human immunodeficiency virus trans-activator of transcription |
| FITC | Fluorescein isothiocyanate |
| HSCs | Hematopoietic stem cells |
| NLS | Nuclear localization signal |
| VPA | Valproic acid |
| TAT | Transactivator of transcription |
| HDFs | Human dermal fibroblasts |
| CE | Corneal endothelia |
| CendR | C-end rule |
| ADSCs | Adipose-derived stem cells |
| RPE | Reprogramming of pigmented epithelial |
| CPCs | Cardiac progenitor cells |
| MITT | Macromolecule intracellular transduction technology |
| MTDs | Macromolecule transduction domains |
References
- Yamanaka, S.; Blau, H.M. Nuclear reprogramming to a pluripotent state by three approaches. Nature 2010, 465, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.A.; Atienza, J.; Melton, D.A.; Eggan, K. Nuclear reprogramming of somatic cells after fusion with human embryonic stem cells. Science 2005, 309, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Bru, T.; Clarke, C.; McGrew, M.J.; Sang, H.M.; Wilmut, I.; Blow, J.J. Rapid induction of pluripotency genes after exposure of human somatic cells to mouse ES cell extracts. Exp. Cell Res. 2008, 314, 2634–2642. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakayama, T.; Shinkai, Y.; Tamashiro, K.L.; Niida, H.; Blanchard, D.C.; Blanchard, R.J.; Ogura, A.; Tanemura, K.; Tachibana, M.; Perry, A.C.; et al. Cloning of mice to six generations. Nature 2000, 407, 318–319. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.C.; Wakayama, T. Untimely ends and new beginnings in mouse cloning. Nat. Genet. 2002, 30, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Do, J.T.; Scholer, H.R. Nuclei of embryonic stem cells reprogram somatic cells. Stem Cells 2004, 22, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Kim, J.S.; Choi, S.; Hong, Y.J.; Kim, M.J.; Seo, H.G.; Do, J.T. Neural stem cells differentiated from iPS cells spontaneously regain pluripotency. Stem Cells 2014, 32, 2596–2604. [Google Scholar] [CrossRef] [PubMed]
- Stadtfeld, M.; Nagaya, M.; Utikal, J.; Weir, G.; Hochedlinger, K. Induced pluripotent stem cells generated without viral integration. Science 2008, 322, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Soldner, F.; Hockemeyer, D.; Beard, C.; Gao, Q.; Bell, G.W.; Cook, E.G.; Hargus, G.; Blak, A.; Cooper, O.; Mitalipova, M.; et al. Parkinson’s disease patient-derived induced pluripotent stem cells free of viral reprogramming factors. Cell 2009, 136, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Norrby, K.; Paca, A.; Mileikovsky, M.; Mohseni, P.; Woltjen, K. Virus-free induction of pluripotency and subsequent excision of reprogramming factors. Nature 2009, 458, 771–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woltjen, K.; Michael, I.P.; Mohseni, P.; Desai, R.; Mileikovsky, M.; Hamalainen, R.; Cowling, R.; Wang, W.; Liu, P.; Gertsenstein, M.; et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. Nature 2009, 458, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Harui, A.; Suzuki, S.; Kochanek, S.; Mitani, K. Frequency and stability of chromosomal integration of adenovirus vectors. J. Virol. 1999, 73, 6141–6146. [Google Scholar] [PubMed]
- Li, H.O.; Zhu, Y.F.; Asakawa, M.; Kuma, H.; Hirata, T.; Ueda, Y.; Lee, Y.S.; Fukumura, M.; Iida, A.; Kato, A.; et al. A cytoplasmic RNA vector derived from nontransmissible Sendai virus with efficient gene transfer and expression. J. Virol. 2000, 74, 6564–6569. [Google Scholar] [CrossRef] [PubMed]
- Fusaki, N.; Ban, H.; Nishiyama, A.; Saeki, K.; Hasegawa, M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Yuasa, S.; Oda, M.; Egashira, T.; Yae, K.; Kusumoto, D.; Nakata, H.; Tohyama, S.; Hashimoto, H.; Kodaira, M.; et al. Generation of induced pluripotent stem cells from human terminally differentiated circulating T cells. Cell Stem Cell 2010, 7, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, K.; Smuga-Otto, K.; Tian, S.; Stewart, R.; Slukvin, I.I.; Thomson, J.A. Human induced pluripotent stem cells free of vector and transgene sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Matsumura, Y.; Sato, Y.; Okada, A.; Morizane, A.; Okamoto, S.; Hong, H.; Nakagawa, M.; Tanabe, K.; Tezuka, K.; et al. A more efficient method to generate integration-free human iPS cells. Nat. Methods 2011, 8, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, L.; Manos, P.D.; Ahfeldt, T.; Loh, Y.H.; Li, H.; Lau, F.; Ebina, W.; Mandal, P.K.; Smith, Z.D.; Meissner, A.; et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 2010, 7, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.K.; Rossi, D.J. Reprogramming human fibroblasts to pluripotency using modified mRNA. Nat. Protoc. 2013, 8, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, N.; Gros, E.; Li, H.R.; Kumar, S.; Deacon, D.C.; Maron, C.; Muotri, A.R.; Chi, N.C.; Fu, X.D.; Yu, B.D.; et al. Efficient generation of human iPSCs by a synthetic self-replicative RNA. Cell Stem Cell 2013, 13, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Anokye-Danso, F.; Trivedi, C.M.; Juhr, D.; Gupta, M.; Cui, Z.; Tian, Y.; Zhang, Y.; Yang, W.; Gruber, P.J.; Epstein, J.A.; et al. Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency. Cell Stem Cell 2011, 8, 376–388. [Google Scholar] [CrossRef]
- Hakelien, A.M.; Landsverk, H.B.; Robl, J.M.; Skalhegg, B.S.; Collas, P. Reprogramming fibroblasts to express T-cell functions using cell extracts. Nat. Biotechnol. 2002, 20, 460–466. [Google Scholar] [CrossRef]
- Landsverk, H.B.; Hakelien, A.M.; Kuntziger, T.; Robl, J.M.; Skalhegg, B.S.; Collas, P. Reprogrammed gene expression in a somatic cell-free extract. EMBO Rep. 2002, 3, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Collas, P.; Hakelien, A.M. Teaching cells new tricks. Trends Biotechnol. 2003, 21, 354–361. [Google Scholar] [CrossRef]
- Taranger, C.K.; Noer, A.; Sorensen, A.L.; Hakelien, A.M.; Boquest, A.C.; Collas, P. Induction of dedifferentiation, genomewide transcriptional programming, and epigenetic reprogramming by extracts of carcinoma and embryonic stem cells. Mol. Biol. Cell 2005, 16, 5719–5735. [Google Scholar] [CrossRef] [PubMed]
- Freberg, C.T.; Dahl, J.A.; Timoskainen, S.; Collas, P. Epigenetic reprogramming of OCT4 and NANOG regulatory regions by embryonal carcinoma cell extract. Mol. Biol. Cell 2007, 18, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Lee, C.S.; Kwon, Y.W.; Paek, J.S.; Lee, S.H.; Hur, J.; Lee, E.J.; Roh, T.Y.; Chu, I.S.; Leem, S.H.; et al. Induction of pluripotent stem cells from adult somatic cells by protein-based reprogramming without genetic manipulation. Blood 2010, 116, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.D.; Pabo, C.O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 1988, 55, 1189–1193. [Google Scholar] [CrossRef]
- Elliott, G.; O’Hare, P. Intercellular trafficking and protein delivery by a herpesvirus structural protein. Cell 1997, 88, 223–233. [Google Scholar] [CrossRef]
- Joliot, A.; Pernelle, C.; Deagostini-Bazin, H.; Prochiantz, A. Antennapedia homeobox peptide regulates neural morphogenesis. Proc. Natl. Acad. Sci. USA 1991, 88, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, S.R.; Ho, A.; Vocero-Akbani, A.; Dowdy, S.F. In vivo protein transduction: Delivery of a biologically active protein into the mouse. Science 1999, 285, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Krosl, J.; Austin, P.; Beslu, N.; Kroon, E.; Humphries, R.K.; Sauvageau, G. In vitro expansion of hematopoietic stem cells by recombinant TAT-HOXB4 protein. Nat. Med. 2003, 9, 1428–1432. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.D.; Oh, S.K.; Kim, H.S.; Ku, S.Y.; Kim, S.H.; Choi, Y.M.; Moon, S.Y. Cellular manipulation of human embryonic stem cells by TAT-PDX1 protein transduction. Mol. Ther. J. Am. Soc. Gene Ther. 2005, 12, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Stock, K.; Nolden, L.; Edenhofer, F.; Quandel, T.; Brustle, O. Transcription factor-based modulation of neural stem cell differentiation using direct protein transduction. Cell. Mol. Life Sci. 2010, 67, 2439–2449. [Google Scholar] [CrossRef] [PubMed]
- Bosnali, M.; Edenhofer, F. Generation of transducible versions of transcription factors Oct4 and Sox2. Biol. Chem. 2008, 389, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wu, S.; Joo, J.Y.; Zhu, S.; Han, D.W.; Lin, T.; Trauger, S.; Bien, G.; Yao, S.; Zhu, Y.; et al. Generation of induced pluripotent stem cells using recombinant proteins. Cell Stem Cell 2009, 4, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, C.H.; Moon, J.I.; Chung, Y.G.; Chang, M.Y.; Han, B.S.; Ko, S.; Yang, E.; Cha, K.Y.; Lanza, R.; et al. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 2009, 4, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.H.; Ko, J.Y.; Chang, M.Y.; Yi, S.H.; Kim, D.; Kim, C.H.; Shim, J.W.; Jo, A.Y.; Kim, B.W.; Lee, H.; et al. Protein-based human iPS cells efficiently generate functional dopamine neurons and can treat a rat model of Parkinson disease. J. Clin. Investig. 2011, 121, 2326–2335. [Google Scholar] [CrossRef] [PubMed]
- McWhirter, S.M.; Fitzgerald, K.A.; Rosains, J.; Rowe, D.C.; Golenbock, D.T.; Maniatis, T. IFN-regulatory factor 3-dependent gene expression is defective in Tbk1-deficient mouse embryonic fibroblasts. Proc. Natl. Acad. Sci. USA 2004, 101, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Sayed, N.; Hunter, A.; Au, K.F.; Wong, W.H.; Mocarski, E.S.; Pera, R.R.; Yakubov, E.; Cooke, J.P. Activation of innate immunity is required for efficient nuclear reprogramming. Cell 2012, 151, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Thier, M.; Munst, B.; Edenhofer, F. Exploring refined conditions for reprogramming cells by recombinant Oct4 protein. Int. J. Dev. Biol. 2010, 54, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Thier, M.; Munst, B.; Mielke, S.; Edenhofer, F. Cellular reprogramming employing recombinant Sox2 protein. Stem Cells Int. 2012, 2012, 549846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, Y.; Gu, J.; Liao, B.; Li, J.; Wong, J.; Jin, Y. Reprogramming of somatic cells via TAT-mediated protein transduction of recombinant factors. Biomaterials 2012, 33, 5047–5055. [Google Scholar] [CrossRef] [PubMed]
- Islas, J.F.; Liu, Y.; Weng, K.C.; Robertson, M.J.; Zhang, S.; Prejusa, A.; Harger, J.; Tikhomirova, D.; Chopra, M.; Iyer, D.; et al. Transcription factors ETS2 and MESP1 transdifferentiate human dermal fibroblasts into cardiac progenitors. Proc. Natl. Acad. Sci. USA 2012, 109, 13016–13021. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Guo, Y.; Wang, C.; Liu, Q.; Yang, Y.; Li, S.; Guo, X.; Lian, R.; Yu, R.; Liu, H.; et al. Non-genetic direct reprogramming and biomimetic platforms in a preliminary study for adipose-derived stem cells into corneal endothelia-like cells. PLoS ONE 2014, 9, e109856. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Chen, R.; Teesalu, T.; Ruoslahti, E.; Clegg, D.O. Reprogramming human retinal pigmented epithelial cells to neurons using recombinant proteins. Stem Cells Transl. Med. 2014, 3, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, Q.; Jiang, L.; Deng, C.; Liu, Z.; Fu, Y.; Zhang, M.; Tan, H.; Feng, Y.; Shan, Z.; et al. Generation of Functional Human Cardiac Progenitor Cells by High-Efficiency Protein Transduction. Stem Cells Transl. Med. 2015, 4, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Choi, H.W.; Cho, J.; Jung, S.; Seo, H.G.; Do, J.T. Activation of pluripotency genes by a nanotube-mediated protein delivery system. Mol. Reprod. Dev. 2013, 80, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Kim, J.; Kang, J.; Jo, D. Partial somatic to stem cell transformations induced by cell-permeable reprogramming factors. Sci. Rep. 2014, 4, 4361. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.M.; Eroglu, E.; Bawage, S.S.; Vig, K.; Miller, M.E.; Pillai, S.; Dennis, V.A.; Singh, S.R. Enhanced intracellular translocation and biodistribution of gold nanoparticles functionalized with a cell-penetrating peptide (VG-21) from vesicular stomatitis virus. Biomaterials 2014, 35, 9484–9494. [Google Scholar] [CrossRef] [PubMed]
- Erazo-Oliveras, A.; Muthukrishnan, N.; Baker, R.; Wang, T.Y.; Pellois, J.P. Improving the endosomal escape of cell-penetrating peptides and their cargos: Strategies and challenges. Pharmaceuticals 2012, 5, 1177–1209. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Source of Somatic Cells | Reprogramming Factors | Type of Protocol | Assessment of Pluripotency | Efficiency | Reference |
|---|---|---|---|---|---|
| 293T or NIH3T3 fibroblasts | Extract of human NCCIT ECCs or mouse ESCs | Exposed to cell extract | Pluripotency | [26] | |
| 293T | Extract of mouse ESCs | [3] | |||
| Mouse cFBs | Extract of mouse ESCs | [28] | |||
| MEFs | OKSM with VPA | 11R-fused CPPs | 0.001% | [37] | |
| HNFs | OKSM | 9R-fused CPPs | 0.001% | [38] | |
| Human fibroblast | OS with PolyI:C | 11R-fused CPPs | [41] | ||
| MEFs | Oct4 with Serum replacement, Sucrose | TAT-fused CPPs | [42] | ||
| MEFs | Sox2 with Serum replacement | TAT-fused CPPs | [43] | ||
| HAF cells HFFs NIH3T3 fibroblasts | OKSMN with VPA | TAT-fused CPPs | 0.012% | [44] | |
| HAF cells HFFs NIH3T3 fibroblasts | OKS | 11R-fused CPPs | [44] | ||
| mNSCs | OKSMN | TiO2 Nanotube-mediated protein delivery | 0.005%–0.01% | [49] | |
| HDFs | OKSML with Vitamin C | MTDs | Partially reprogrammed | 0.34% | [50] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, B.J.; Hong, Y.J.; Do, J.T. Cellular Reprogramming Using Protein and Cell-Penetrating Peptides. Int. J. Mol. Sci. 2017, 18, 552. https://doi.org/10.3390/ijms18030552
Seo BJ, Hong YJ, Do JT. Cellular Reprogramming Using Protein and Cell-Penetrating Peptides. International Journal of Molecular Sciences. 2017; 18(3):552. https://doi.org/10.3390/ijms18030552
Chicago/Turabian StyleSeo, Bong Jong, Yean Ju Hong, and Jeong Tae Do. 2017. "Cellular Reprogramming Using Protein and Cell-Penetrating Peptides" International Journal of Molecular Sciences 18, no. 3: 552. https://doi.org/10.3390/ijms18030552
APA StyleSeo, B. J., Hong, Y. J., & Do, J. T. (2017). Cellular Reprogramming Using Protein and Cell-Penetrating Peptides. International Journal of Molecular Sciences, 18(3), 552. https://doi.org/10.3390/ijms18030552