
Inhibition of NLRP3 Inflammasome Pathway by Butyrate Improves Corneal Wound Healing in Corneal Alkali Burn



Abstract
:1. Introduction
2. Results
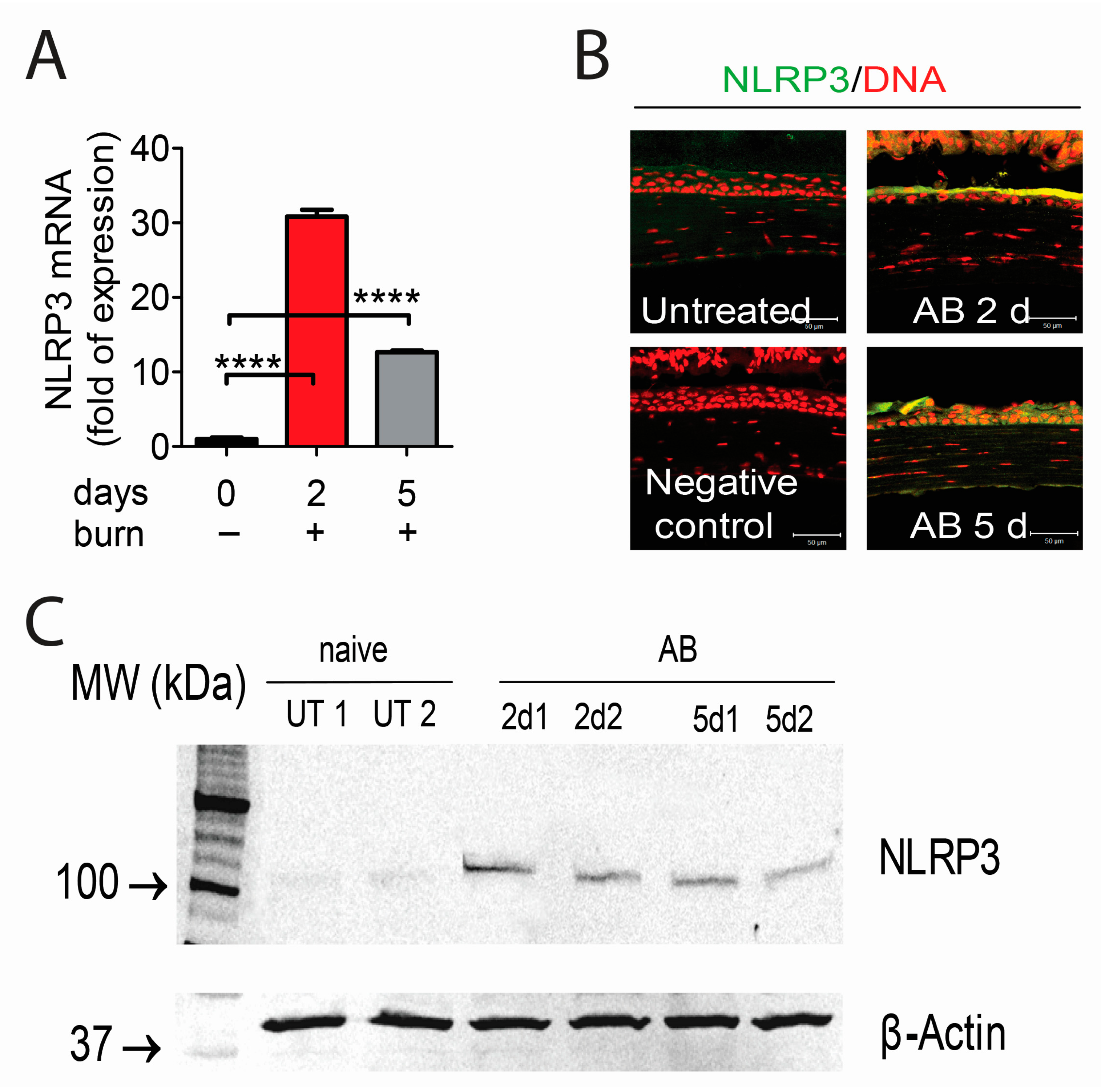
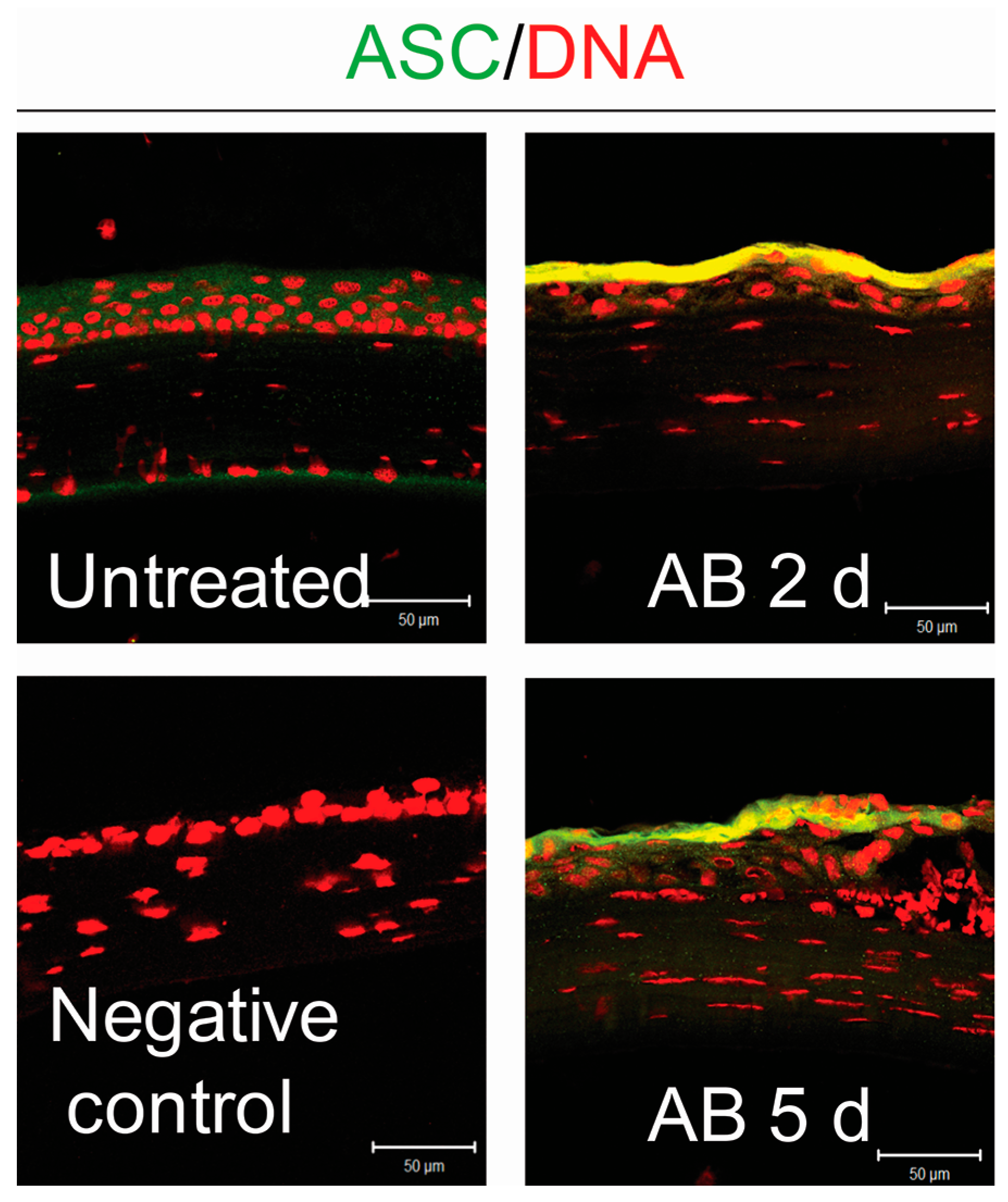
2.1. Corneal Alkali Burn Upregulates NLRP3 and Adaptor Protein ASC in Corneal Epithelium
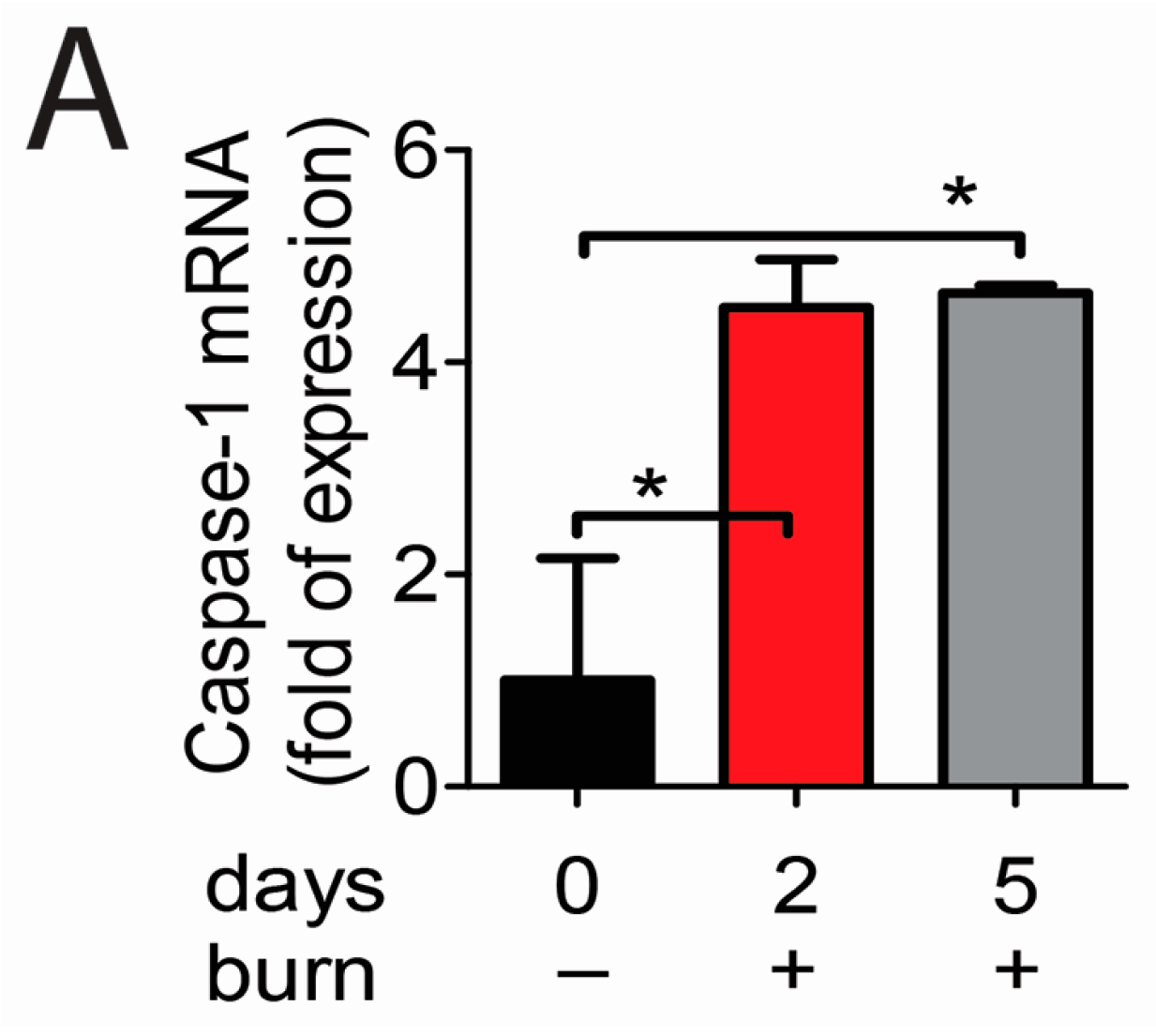
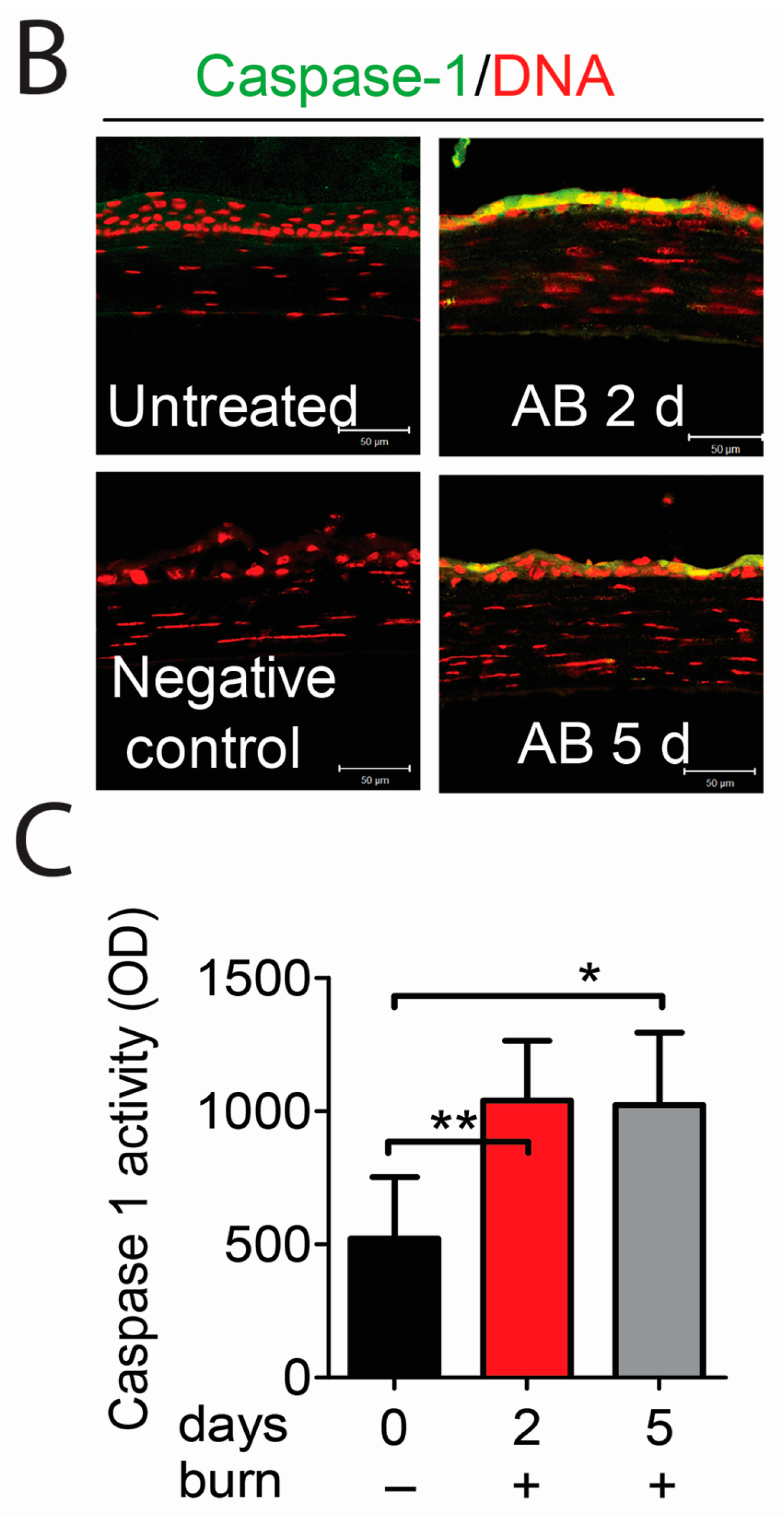
2.2. Caspase-1 Activation in Alkali-Injured Corneas
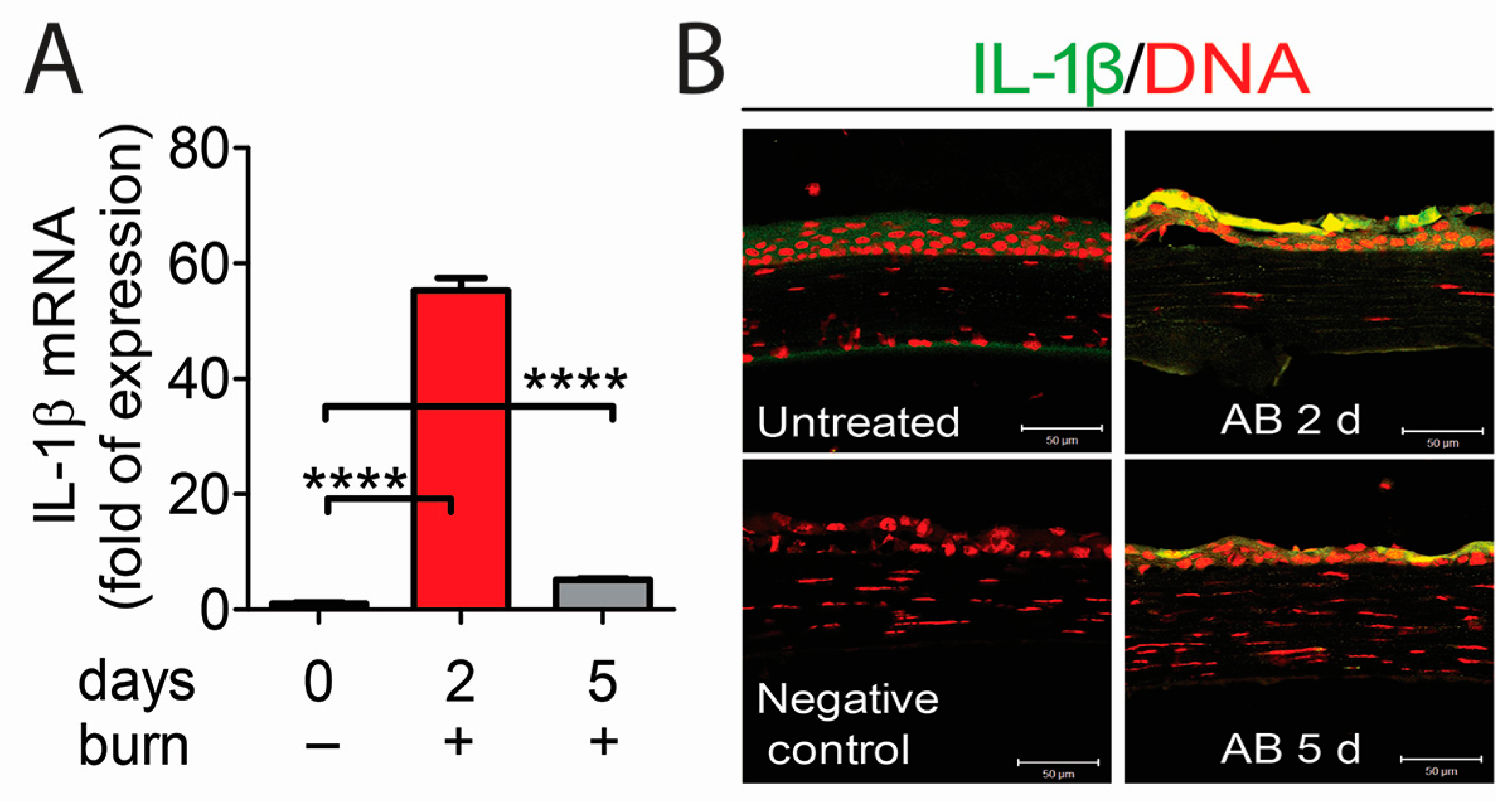
2.3. The NLRP3–ASC–Caspase-1 Inflammasome Directs IL-1β Secretion in Alkali-Injured Corneas
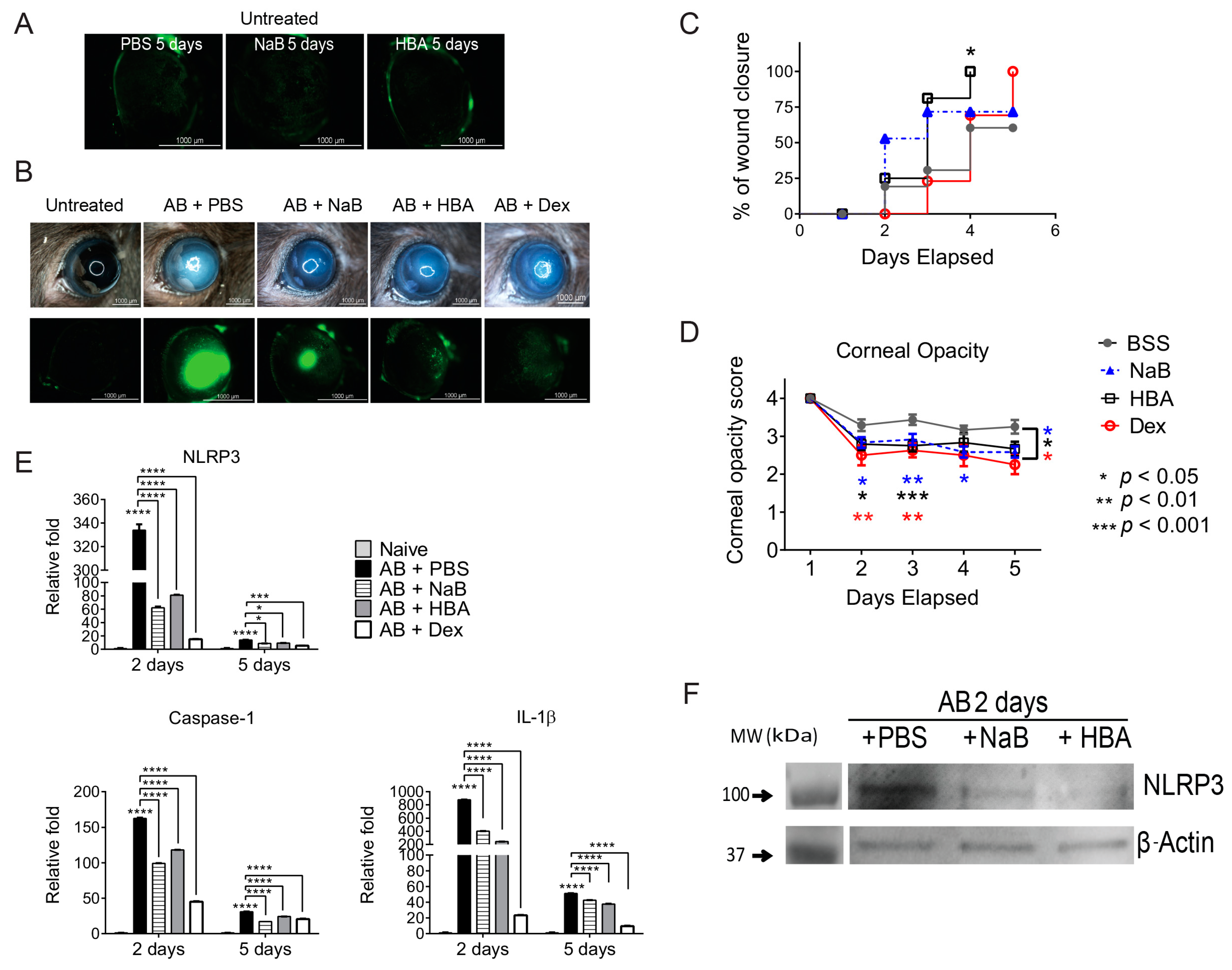
2.4. Sodium Butyrate and β-Hydroxybutyric Acid Topical Treatment Inhibits Activation of NLRP3 Inflammasome in Alkali-Injured Corneas
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Unilateral Alkali Burn
4.3. Histology and Immunostaining
4.4. RNA Isolation and Quantitative PCR
4.5. Caspase-1 Activation Fluorometric Assays
4.6. Western Blot
4.7. Treatment Regimen
4.8. Clinical Findings: Opacity Score
4.9. Measurement of Corneal Epithelial Defect
4.10. Number of Animals and Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AB | alkali burn |
| d | days |
| UT | untreated |
| IL | interleukin |
| NaB | Sodium butyrate |
| HBA | β-hydroxybutyric acid |
| BSS | balanced salt solution |
| ASC | apoptosis-associated speck-like protein |
| B2M | beta-2-microglobulin |
| BCA | bicinchoninic acid |
| Dex | dexamethasone |
| NOD | nucleotide-binding oligomerization domain-containing protein |
| NLR | NOD-like receptor |
| PAMPs | pathogen-associated molecular patterns |
| TLR | Toll like receptor |
| DAMPs | damage-associated molecular patterns |
| ANOVA | one-way analysis of variance |
| CARD | caspase activation and recruitment domain |
| QID | quater in die (in Latin, four times a day) |
References
- Clare, G.; Suleman, H.; Bunce, C.; Dua, H. Amniotic membrane transplantation for acute ocular burns. Cochrane Database Syst. Rev. 2012, 9, CD009379. [Google Scholar]
- Shao, L.; Kamalu, O.; Mayer, L. Non-classical MHC class i molecules on intestinal epithelial cells: Mediators of mucosal crosstalk. Immunol. Rev. 2005, 206, 160–176. [Google Scholar] [CrossRef] [PubMed]
- Yoshikai, Y. The interaction of intestinal epithelial cells and intraepithelial lymphocytes in host defense. Immunol. Res. 1999, 20, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, J.; Xin, Z.; Wu, X. Aspergillus fumigatus triggers innate immune response via NOD1 signaling in human corneal epithelial cells. Exp. Eye Res. 2014, 127, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Bals, R.; Hiemstra, P.S. Innate immunity in the lung: How epithelial cells fight against respiratory pathogens. Eur. Respir. J. 2004, 23, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Rajalakshmy, A.R.; Malathi, J.; Madhavan, H.N. HCV core and NS3 proteins mediate toll like receptor induced innate immune response in corneal epithelium. Exp. Eye Res. 2014, 128, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, K.; Ambati, B.; Yu, F.S. Toll-like receptor 5-mediated corneal epithelial inflammatory responses to Pseudomonas aeruginosa flagellin. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4247–4254. [Google Scholar] [CrossRef]
- Eslani, M.; Movahedan, A.; Afsharkhamseh, N.; Sroussi, H.; Djalilian, A.R. The role of toll-like receptor 4 in corneal epithelial wound healing. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6108–6115. [Google Scholar] [CrossRef] [PubMed]
- Edye, M.E.; Lopez-Castejon, G.; Allan, S.M.; Brough, D. Acidosis drives damage-associated molecular pattern (damp)-induced interleukin-1 secretion via a caspase-1-independent pathway. J. Biol. Chem. 2013, 288, 30485–30494. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Pulskens, W.P.; Sadler, J.J.; Butter, L.M.; Teske, G.J.; Ulland, T.K.; Eisenbarth, S.C.; Florquin, S.; Flavell, R.A.; Leemans, J.C.; et al. Necrotic cells trigger a sterile inflammatory response through the NLRP3 inflammasome. Proc. Natl. Acad. Sci. USA 2009, 106, 20388–20393. [Google Scholar] [CrossRef] [PubMed]
- Pajak, B.; Orzechowski, A.; Gajkowska, B. Molecular basis of sodium butyrate-dependent proapoptotic activity in cancer cells. Adv. Med. Sci. 2007, 52, 83–88. [Google Scholar] [PubMed]
- Wang, X.; He, G.; Peng, Y.; Zhong, W.; Wang, Y.; Zhang, B. Sodium butyrate alleviates adipocyte inflammation by inhibiting NRLP3 pathway. Sci. Rep. 2015, 5, 12676. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Shibuya, M.; Saito, Y.; Nakashima, H.; Saito, F.; Higuchi, A.; Tsubota, K. Protective effect of d-β-hydroxybutyrate on corneal epithelia in dry eye conditions through suppression of apoptosis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4682–4688. [Google Scholar] [CrossRef]
- Franchi, L.; Munoz-Planillo, R.; Nunez, G. Sensing and reacting to microbes through the inflammasomes. Nat. Immunol. 2012, 13, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed]
- Negash, A.A.; Ramos, H.J.; Crochet, N.; Lau, D.T.; Doehle, B.; Papic, N.; Delker, D.A.; Jo, J.; Bertoletti, A.; Hagedorn, C.H.; et al. IL-1β production through the NRLP3 inflammasome by hepatic macrophages links hepatitis C virus infection with liver inflammation and disease. PLoS Pathog. 2013, 9, e1003330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannitti, R.G.; Napolioni, V.; Oikonomou, V.; de Luca, A.; Galosi, C.; Pariano, M.; Massi-Benedetti, C.; Borghi, M.; Puccetti, M.; Lucidi, V.; et al. IL-1 receptor antagonist ameliorates inflammasome-dependent inflammation in murine and human cystic fibrosis. Nat. Commun. 2016, 7, 10791. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A. Inflammasome inhibition: Putting out the fire. Cell Metab. 2015, 21, 513–514. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Pelegrino, F.S.; Tukler Henriksson, J.; Pflugfelder, S.C.; Volpe, E.A.; Li, D.Q.; de Paiva, C.S. Differential effects of dexamethasone and doxycycline on inflammation and MMP production in alkali-burned corneas associated with dry eye. Ocular Surface 2016, 14, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Nunez, G. Sterile inflammation: Sensing and reacting to damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, E.; Campbell, M.; Doyle, S.L. Targeting the NRLP3 inflammasome in chronic inflammatory diseases: Current perspectives. J. Inflamm. Res. 2015, 8, 15–27. [Google Scholar] [PubMed]
- Ghiringhelli, F.; Apetoh, L.; Tesniere, A.; Aymeric, L.; Ma, Y.; Ortiz, C.; Vermaelen, K.; Panaretakis, T.; Mignot, G.; Ullrich, E.; et al. Activation of the NRLP3 inflammasome in dendritic cells induces IL-1β-dependent adaptive immunity against tumors. Nat. Med. 2009, 15, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Puyang, Z.; Feng, L.; Chen, H.; Liang, P.; Troy, J.B.; Liu, X. Retinal ganglion cell loss is delayed following optic nerve crush in NRLP3 knockout mice. Sci. Rep. 2016, 6, 20998. [Google Scholar] [CrossRef] [PubMed]
- Mulay, S.R.; Kulkarni, O.P.; Rupanagudi, K.V.; Migliorini, A.; Darisipudi, M.N.; Vilaysane, A.; Muruve, D.; Shi, Y.; Munro, F.; Liapis, H.; et al. Calcium oxalate crystals induce renal inflammation by NRLP3-mediated IL-1β secretion. J. Clin. Investig. 2013, 123, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; She, Y.; Xiao, B.; Hua, S.; Qu, J.; Chen, W. Reactive oxygen species activated NRLP3 inflammasomes prime environment-induced murine dry eye. Exp. Eye Res. 2014, 125, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chi, W.; Hua, X.; Bian, F.; Yuan, X.; Deng, R.; Zhang, Z.; de Paiva, C.S.; Pflugfelder, S.C.; Li, D.-Q. Caspase-8 mediated activation of NLRP3 and NLRC4 inflammasomes in experimental dry eye mouse model and human corneal epithelial cells exposed to hyperosmolarity. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4878. [Google Scholar]
- McGilligan, V.E.; Gregory-Ksander, M.S.; Li, D.; Moore, J.E.; Hodges, R.R.; Gilmore, M.S.; Moore, T.C.; Dartt, D.A. Staphylococcus aureus activates the NRLP3 inflammasome in human and rat conjunctival goblet cells. PLoS ONE 2013, 8, e74010. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hanus, J.W.; Abu-Asab, M.S.; Shen, D.; Ogilvy, A.; Ou, J.; Chu, X.K.; Shi, G.; Li, W.; Wang, S.; et al. NLRP3 upregulation in retinal pigment epithelium in age-related macular degeneration. Int. J. Mol. Sci. 2016, 17, 73. [Google Scholar] [CrossRef] [PubMed]
- Corrales, R.M.; Stern, M.E.; de Paiva, C.S.; Welch, J.; Li, D.Q.; Pflugfelder, S.C. Desiccating stress stimulates expression of matrix metalloproteinases by the corneal epithelium. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3293–3302. [Google Scholar] [CrossRef] [PubMed]
- Thermo Fisher Scientific. Available online: www.lifetechnologies.com (accessed on 28 February 2017).
- De Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D., III; Fang, B.; Zheng, X.; Ma, P.; Pangelinan, W.J.; Siemasko, K.S.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Shibuya, M.; Nakashima, H.; Imagawa, T.; Uehara, M.; Tsubota, K. d-β-Hydroxybutyrate protects against corneal epithelial disorders in a rat dry eye model with jogging board. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2379–2387. [Google Scholar] [CrossRef] [PubMed]
- Yoeruek, E.; Ziemssen, F.; Henke-Fahle, S.; Tatar, O.; Tura, A.; Grisanti, S.; Bartz-Schmidt, K.U.; Szurman, P.; Tübingen Bevacizumab Study Group. Safety, penetration and efficacy of topically applied bevacizumab: Evaluation of eyedrops in corneal neovascularization after chemical burn. Acta Ophthalmol. 2008, 86, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Pelegrino, F.S.; Pflugfelder, S.C.; Volpe, E.A.; Li, D.Q.; de Paiva, C.S. Desiccating stress-induced MMP production and activity worsens wound healing in alkali-burned corneas. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4908–4918. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Symbol | Assay ID * |
|---|---|---|
| β-2-microglobulin | B2M | Mm00437762 |
| Caspase-1 | Caspase-1 | Mm00438023 |
| Interleukin-1β | IL-1β | Mm00434228 |
| NLR family, pyrin domain containing 3 | NLRP3 | Mm00840904 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, F.; Xiao, Y.; Zaheer, M.; Volpe, E.A.; Pflugfelder, S.C.; Li, D.-Q.; De Paiva, C.S. Inhibition of NLRP3 Inflammasome Pathway by Butyrate Improves Corneal Wound Healing in Corneal Alkali Burn. Int. J. Mol. Sci. 2017, 18, 562. https://doi.org/10.3390/ijms18030562
Bian F, Xiao Y, Zaheer M, Volpe EA, Pflugfelder SC, Li D-Q, De Paiva CS. Inhibition of NLRP3 Inflammasome Pathway by Butyrate Improves Corneal Wound Healing in Corneal Alkali Burn. International Journal of Molecular Sciences. 2017; 18(3):562. https://doi.org/10.3390/ijms18030562
Chicago/Turabian StyleBian, Fang, Yangyan Xiao, Mahira Zaheer, Eugene A. Volpe, Stephen C. Pflugfelder, De-Quan Li, and Cintia S. De Paiva. 2017. "Inhibition of NLRP3 Inflammasome Pathway by Butyrate Improves Corneal Wound Healing in Corneal Alkali Burn" International Journal of Molecular Sciences 18, no. 3: 562. https://doi.org/10.3390/ijms18030562
APA StyleBian, F., Xiao, Y., Zaheer, M., Volpe, E. A., Pflugfelder, S. C., Li, D. -Q., & De Paiva, C. S. (2017). Inhibition of NLRP3 Inflammasome Pathway by Butyrate Improves Corneal Wound Healing in Corneal Alkali Burn. International Journal of Molecular Sciences, 18(3), 562. https://doi.org/10.3390/ijms18030562