Unconventional Transport Routes of Soluble and Membrane Proteins and Their Role in Developmental Biology

 ,
, 
 and
and 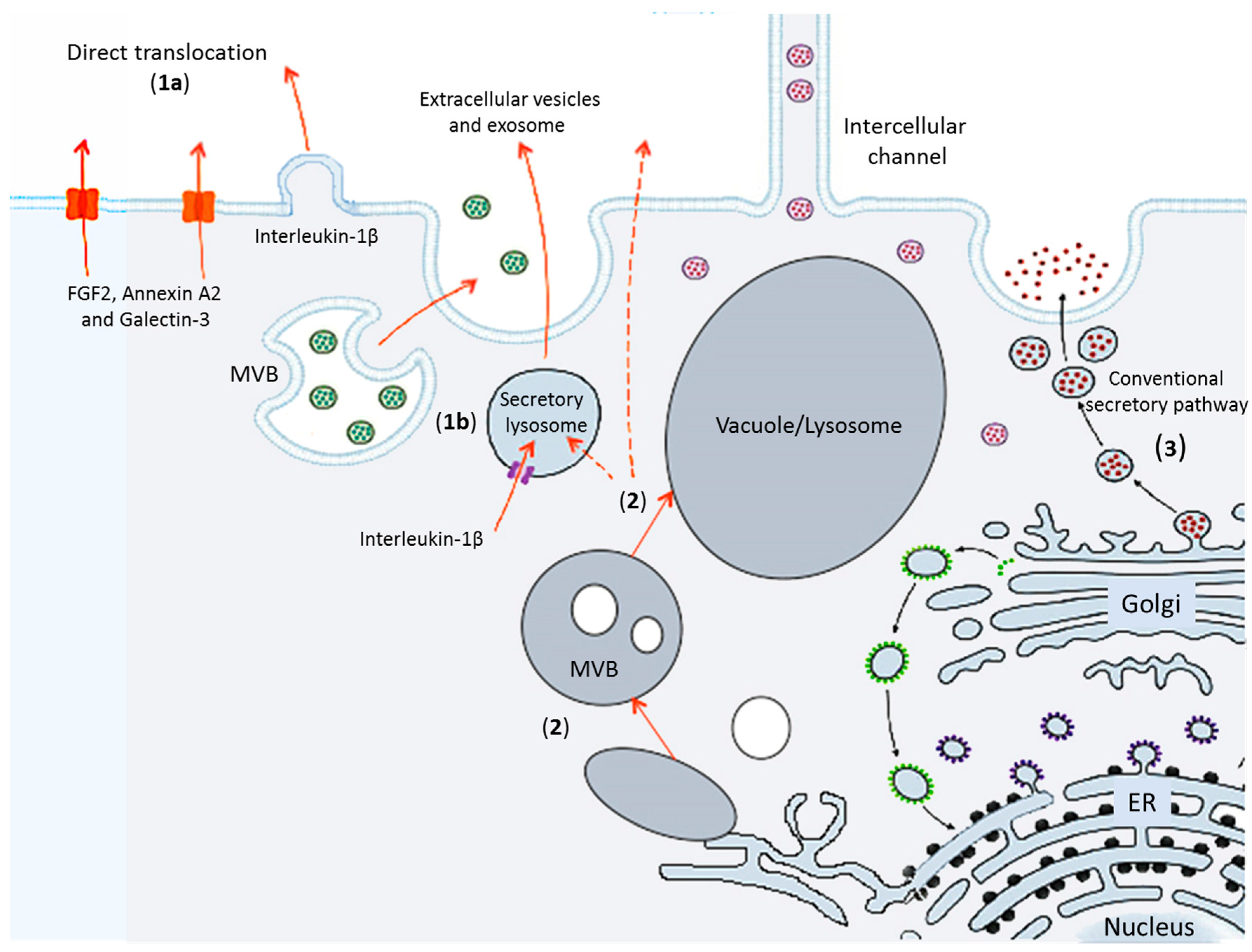{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Routes for Unconventional Secretion of Leaderless Proteins to the Extra Cellular Space
2.1. Pore Formation at the PM
2.2. Different Types of Secretory Vesicles for Leaderless Proteins Translocation
2.3. Other Examples of Unconventional Leaderless Proteins Secretion
3. Routes for Unconventional Secretion of Soluble or Transmembrane Proteins with ER Localization Subsequently Transported to the PM, to the Vacuole, or to the Extra Cellular Space
3.1. Involvement of Autophagy in Unconventional Secretion
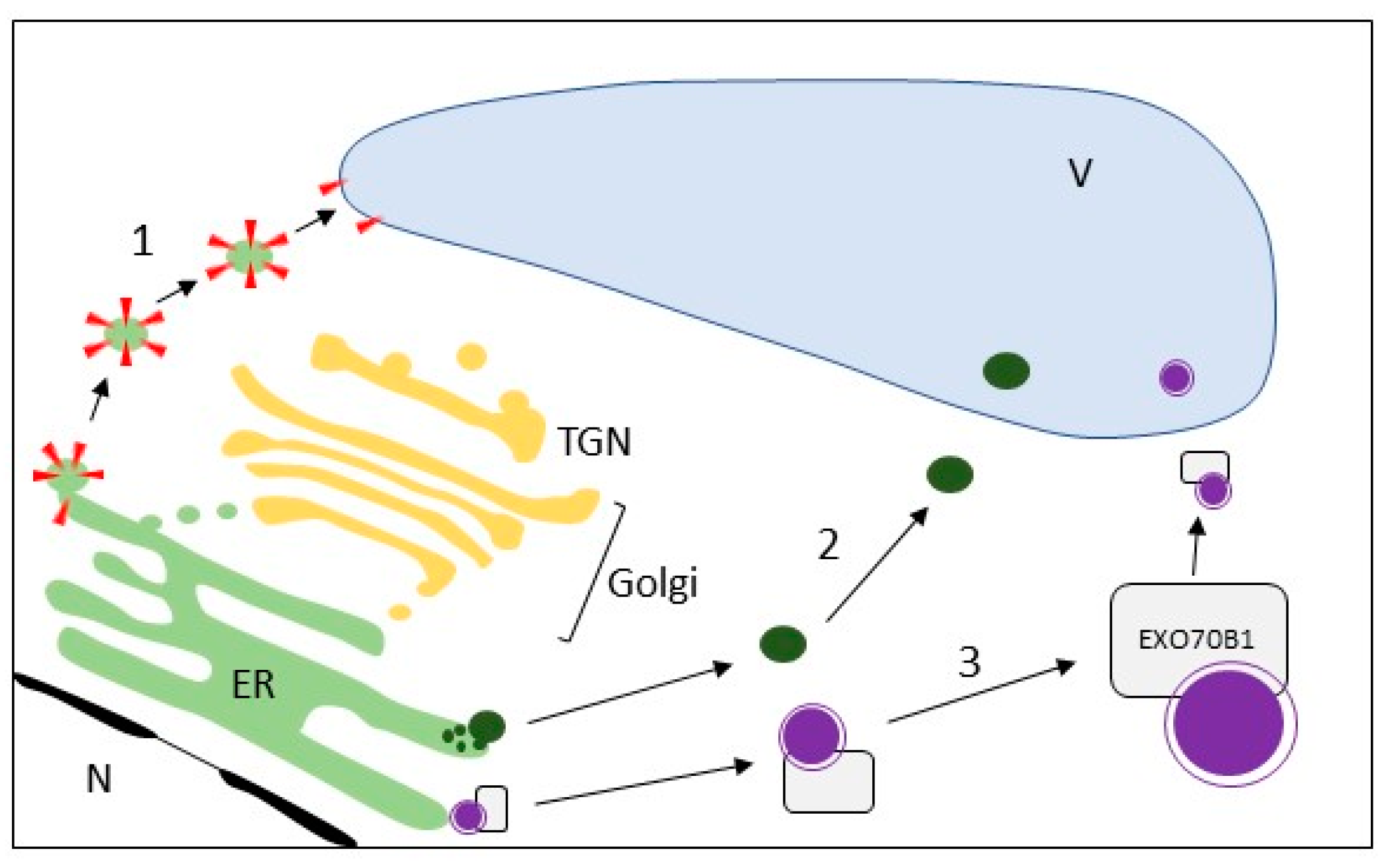
3.2. Direct Delivery from the ER to the Vacuole
3.3. Exosome Release
4. Intercellular Channels
5. Unconventional Role of Proteins Normally Functioning in Conventional Protein and Membrane Traffic
5.1. Exocyst
5.2. SNAP29
5.3. Interfering SNAREs
5.4. RAB Proteins
5.5. AP-2 Adaptor Complex
5.6. Invariant Chain
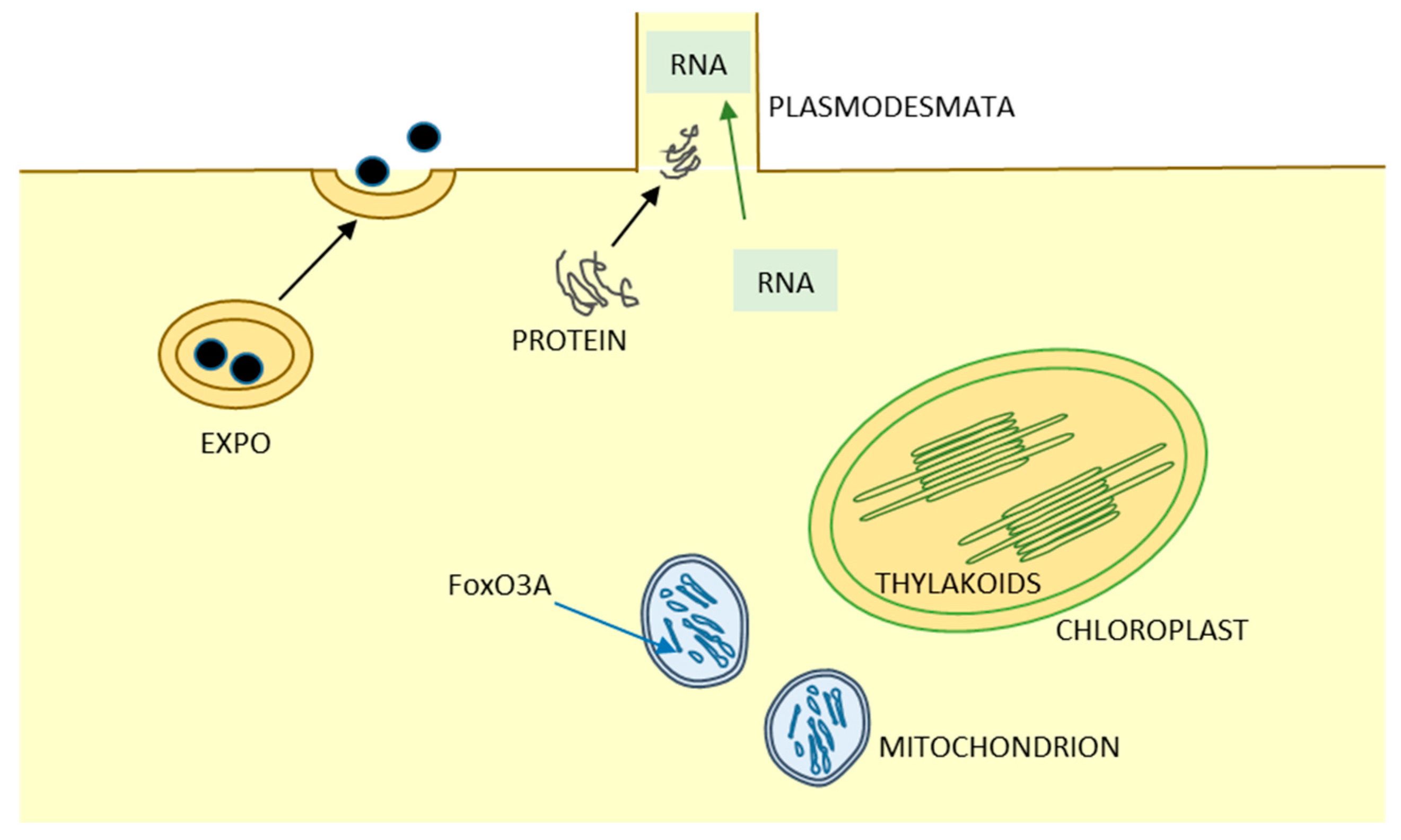
6. Unusual or Unexplored Intra- and Intercellular Pathways and Organelle Biogenesis
6.1. Plant Sexuality
6.2. EXPO
6.3. Mitochondrion
6.4. Chloroplast
6.5. RNA as Molecular Signals in Plant Biology
6.6. Other Examples of Unusual or Unexplored Intra- and Intercellular Pathways
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vitale, A.; Denecke, J. The Endoplasmic Reticulum—Gateway of the Secretory Pathway. Plant Cell 1999, 11, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Bonifacino, J.S.; Glick, B.S. The mechanisms of vesicle budding and fusion. Cell 2004, 116, 153–166. [Google Scholar] [CrossRef]
- Viotti, C. ER to Golgi-Dependent Protein Secretion: The Conventional Pathway. In Unconventional Protein Secretion, Methods in Molecular Biology; Pompa, A., De Marchis, F., Eds.; Humana Press Inc.: New York, NY, USA, 2016; Volume 1459, pp. 3–29. [Google Scholar]
- Robinson, D.G.; Ding, Y.; Jiang, L. Unconventional protein secretion in plants: A critical assessment. Protoplasma 2016, 253, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.J.; Kang, B.-H.; Heringer, A.S.; Wilkop, T.E.; Drakakaki, G. Unconventional Protein Secretion in Plants. In Unconventional Protein Secretion, Methods in Molecular Biology; Pompa, A., De Marchis, F., Eds.; Humana Press Inc.: New York, NY, USA, 2016; Volume 1459, pp. 47–63. [Google Scholar]
- Ng, F.; Tang, B.L. Unconventional Protein Secretion in Animal Cells. In Unconventional Protein Secretion, Methods in Molecular Biology; Pompa, A., De Marchis, F., Eds.; Humana Press Inc.: New York, NY, USA, 2016; Volume 1459, pp. 31–46. [Google Scholar]
- Bhattacharya, A.; Prakash, Y.S.; Eissa, N.T. Secretory function of autophagy in innate immune cells. Cell Microbiol. 2014, 16, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Nickel, W.; Rabouille, C. Mechanisms of regulated unconventional protein secretion. Nat. Rev. Mol. Cell Biol. 2009, 10, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Moss, S.E. Annexins: From structure to function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Valapala, M.; Maji, S.; Borejdo, J.; Vishwanatha, J.K. Cell surface translocation of annexin A2 facilitates glutamate-induced extracellular proteolysis. J. Biol. Chem. 2014, 289, 15915–15926. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Maji, S.; Borejdo, J.; Vishwanatha, J.K. Tyrosine 23 phosphorylation-dependent cell-surface localization of annexin A2 is required for invasion and metastases of pancreatic cancer. PLoS ONE 2011, 6, e19390. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.J.; Longbottom, R.E.; Evans, M.A.; Moss, S.E. Annexinopathies. Subcell. Biochem. 2007, 45, 1–28. [Google Scholar] [PubMed]
- Sidera, K.; Patsavoudi, E. Extracellular HSP90: Conquering the cell surface. Cell Cycle 2008, 7, 1564–1568. [Google Scholar] [CrossRef] [PubMed]
- Sheokand, N.; Malhotra, H.; Kumar, S.; Tillu, V.A.; Chauhan, A.S.; Raje, C.I.; Raje, M. Moonlighting cell-surface GAPDH recruits apotransferrin to effect iron egress from mammalian cells. J. Cell Sci. 2014, 127, 4279–4291. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Rubinstein, N.; Fainboim, L. Unlocking the secrets of galectins: A challenge at the frontier of glyco-immunology. J. Leukoc. Biol. 2002, 71, 741–752. [Google Scholar] [PubMed]
- Pinedo, M.; Regente, M.; Elizalde, M.; Quiroga, I.Y.; Pagnussat, L.A.; Jorrin-Novo, J.; Maldonado, A.; de la Canal, L. Extracellular sunflower proteins: Evidence on non-classical secretion of a jacalin-related lectin. Protein Pept. Lett. 2012, 19, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: Where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Kowala, J.; Arras, G.; Colombo, M.; Jouvea, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed]
- Prada, I.; Meldolesi, J. Binding and Fusion of Extracellular Vesicles to the Plasma Membrane of Their Cell Targets. Int. J. Mol. Sci. 2016, 17, E1296. [Google Scholar] [CrossRef] [PubMed]
- Lavieri, R.; Rubartelli, A.; Carta, S. Redox stress unbalances the inflammatory cytokine network: Role in autoinflammatory patients and healthy subjects. J. Leukoc. Biol. 2016, 99, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Rubartelli, A. Autoinflammatory diseases. Immunol. Lett. 2014, 161, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Piccioli, P.; Rubartelli, A. The secretion of IL-1β and options for release. Semin. Immunol. 2013, 25, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Zacherl, S.; La Venuta, G.; Muller, H.M.; Wegehingel, S.; Dimou, E.; Sehr, P.; Lewis, J.D.; Erfle, H.; Pepperkok, R.; Nickel, W. A direct role for ATP1A1 in unconventional secretion of fibroblast growth factor 2. J. Biol. Chem. 2015, 290, 3654–3665. [Google Scholar] [CrossRef] [PubMed]
- Steringer, J.P.; Bleicken, S.; Andreas, H.; Zacherl, S.; Laussmann, M.; Temmerman, K.; Contreras, F.X.; Bharat, T.A.M.; Lecgner, J.; Muller, H.M.; et al. Phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2)-dependent oligomerization of fibroblast growth factor 2 (FGF2) triggers the formation of a lipidic membrane pore implicated in unconventional secretion. J. Biol. Chem. 2012, 287, 27659–27669. [Google Scholar] [CrossRef]
- Muller, H.M.; Steringer, J.P.; Wegehingel, S.; Bleicken, S.; Münster, M.; Dimou, E.; Unger, S.; Weidmann, G.; Andreas, H.; García-Sáez, A.J.; et al. Formation of disulfide bridges drives oligomerization, membrane pore formation, and translocation of fibroblast growth factor 2 to cell surfaces. J. Biol. Chem. 2015, 290, 8925–8937. [Google Scholar] [CrossRef] [PubMed]
- Mandelkow, E.M.; Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb. Perspect. Med. 2012, 2, a006247. [Google Scholar] [CrossRef] [PubMed]
- De Torre-Minguela, C.; Barberà-Cremades, M.; Gómez, A.I.; Martín-Sánchez, F.; Pelegrín, P. Macrophage activation and polarization modify P2X7 receptor secretome influencing the inflammatory process. Sci. Rep. 2016, 6, 22586. [Google Scholar] [CrossRef] [PubMed]
- Martin-Sanchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.; Mortellaro, A.; et al. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Bianco, F.; Perrotta, C.; Novellino, L.; Francolini, M.; Riganti, L.; Menna, E.; Saglietti, L.; Schuchman, E.H.; Furlan, R.; Clementi, E.; et al. Acid sphingomyelinase activity triggers microparticle release from glial cells. EMBO J. 2009, 28, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, F.; Turola, E.; Riganti, L.; Caleo, M.; Gabrielli, M.; Perrotta, C.; Novellino, L.; Clementi, E.; Giussani, P.; Viani, P.; et al. Microvesicles released from microglia stimulate synaptic activity via enhanced sphingolipid metabolism. EMBO J. 2012, 31, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Verderio, C.; Muzio, L.; Turola, E.; Bergami, A.; Novellino, L.; Ruffini, F.; Riganti, L.; Corradini, I.; Francolini, M.; Garzetti, L.; et al. Myeloid microvesicles are a marker and therapeutic target for neuroinflammation. Ann. Neurol. 2012, 72, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.; Turola, E.; Ruiz, A.; Bergami, A.; Libera, D.D.; Benussi, L.; Giussani, P.; Magnani, G.; Comi, G.; Legname, G.; et al. Microglia convert aggregated amyloid-β into neurotoxic forms through the shedding of microvesicles. Cell Death Differ. 2014, 21, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Agosta, F.M.; Dalla Libera, D.; Spinelli, E.G.; Finardi, A.; Canu, E.; Bergami, A.; Bocchio Chiavetto, L.; Baronio, M.; Comi, G.; Martino, G.; et al. Myeloid microvescicles in cerebrospinal fluid are associated with myelin damage and neuronal loss in mild cognitive impairment and Alzheimer disease. Ann. Neurol. 2014, 76, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.L.; Jackson, W.T. Intracellular Vesicle Acidification Promotes Maturation of Infectious Poliovirus Particles. PLoS Pathog. 2012, 8, e1003046. [Google Scholar] [CrossRef] [PubMed]
- Curwin, A.J.; Brouwers, N.; Alonso, Y.; Adell, M.; Teis, D.; Turacchio, G.; Parashuraman, S.; Ronchi, P.; Malhotra, V. ESCRT-III drives the final stages of CUPS maturation for unconventional protein secretion. Elife 2016, 26, e16299. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, D.; Curwin, A.J.; Popoff, J.F.; Bruns, C.; Duran, J.M.; Malhotra, V. Remodeling of secretory compartments creates CUPS during nutrient starvation. J. Cell Biol. 2014, 207, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Langner, T.; Özturk, M.; Hartmann, S.; Cord-Landwehr, S.; Moerschbacher, B.; Walton, J.D.; Göhre, V. Chitinases are essential for cell separation in Ustilago maydis. Eukaryot. Cell 2015, 14, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Stock, J.; Terfrüchte, M.; Schipper, K. A Reporter System to Study Unconventional Secretion of Proteins Avoiding N-glycosylation in Ustilago maydis. In Unconventional Protein Secretion, Methods in Molecular Biology; Pompa, A., de Marchis, F., Eds.; Humana Press Inc.: New York, NY, USA, 2016; Volume 1459, pp. 149–160. [Google Scholar]
- Stock, J.; Sarkari, P.; Kreibich, S.; Brefort, T.; Feldbrügge, M.; Schipper, K. Applying unconventional secretion of the endochitinase Cts1 to export heterologous proteins in Ustilago maydis. J. Biotechnol. 2012, 161, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Pallotta, M.T.; Orabona, C.; Volpi, C.; Vacca, C.; Belladonna, M.L.; Bianchi, R.; Servillo, G.; Brunacci, C.; Calvitti, M.; Bicciato, S.; et al. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat. Immunol. 2011, 12, 870–878. [Google Scholar] [CrossRef] [PubMed]
- De Marchis, F.; Bellucci, M.; Pompa, A. Unconventional pathways of secretory plant proteins from the endoplasmic reticulum to the vacuole bypassing the Golgi complex. Plant Signal. Behav. 2013, 8, e25129. [Google Scholar] [CrossRef] [PubMed]
- Viotti, C. ER and vacuoles: Never been closer. Front. Plant. Sci. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Jennelle, L.; Hunegnaw, R.; Dubrovsky, L.; Pushkarsky, T.; Fitzgerald, M.L.; Sviridov, D.; Popratiloff, A.; Brichacek, B.; Bukrinsky, M. HIV-1 protein Nef inhibits activity of ATP-binding cassette transporter A1 by targeting Endoplasmic Reticulum chaperone Calnexin. J. Biol. Chem. 2014, 289, 28870–28884. [Google Scholar] [CrossRef]
- Kulich, I.; Pečenková, T.; Sekereš, J.; Smetana, O.; Fendrych, M.; Foissner, I.; Höftberger, M.; Zárský, V. Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole. Traffic 2013, 14, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Pedrazzini, E.; Caprera, A.; Fojadelli, I.; Stella, A.; Rocchetti, A.; Bassin, B.; Martinoia, E.; Vitale, A. The Arabidopsis tonoplast is almost devoid of glycoproteins with complex N-glycans, unlike the rat lysosomal membrane. J. Exp. Bot. 2016, 67, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Pedrazzini, E.; Komarova, N.Y.; Rentsch, D.; Vitale, A. Traffic routes and signals for the tonoplast. Traffic 2013, 14, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Stigliano, E.; Faraco, M.; Neuhaus, J.M.; Montefusco, A.; Dalessandro, G.; Piro, G.; Di Sansebastiano, G.P. Two glycosylated vacuolar GFPs are new markers for ER-to-vacuole sorting. Plant Physiol. Biochem. 2013, 73, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Occhialini, A.; Gouzerh, G.; Di Sansebastiano, G.P.; Neuhaus, J.M. Dimerization of the Vacuolar Receptors AtRMR1 and -2 from Arabidopsis thaliana Contributes to Their Localization in the trans-Golgi Network. Int. J. Mol. Sci. 2016, 17, E1661. [Google Scholar] [CrossRef] [PubMed]
- Staudt, C.; Puissant, E.; Boonen, M. Subcellular Trafficking of Mammalian Lysosomal Proteins: An Extended View. Int. J. Mol. Sci. 2017, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- De Marcos Lousa, C.; Denecke, J. Lysosomal and vacuolar sorting: Not so different after all! Biochem. Soc. Trans. 2016, 44, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Hilscher, J.; Kapusi, E.; Stoger, E.; Ibl, V. Cell layer-specific distribution of transiently expressed barley ESCRT-III component HvVPS60 in developing barley endosperm. Protoplasma 2016, 253, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Pajonk, S.; Micali, C.; O’Connell, R.; Schulze-Lefert, P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J. 2009, 57, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Richter, S.; Kientz, M.; Brumm, S.; Nielsen, M.E.; Park, M.; Gavidia, R.; Krause, C.; Voss, U.; Beckmann, H.; Mayer, U.; et al. Delivery of endocytosed proteins to the cell-division plane requires change of pathway from recycling to secretion. Elife 2014, 8, e02131. [Google Scholar] [CrossRef] [PubMed]
- Otero, S.; Helariutta, Y.; Benitez-Alfonso, Y. Symplastic communication in organ formation and tissue patterning. Curr. Opin. Plant Biol. 2016, 29, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Benitez-Alfonso, Y. Roles and regulation of plant cell walls surrounding plasmodesmata. Curr. Opin. Plant Biol. 2014, 22, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, H.H.; Rustom, A.; Wang, X. Tunneling nanotubes, an emerging intercellular communication route in development. Mech. Dev. 2013, 130, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Knox, K.; Wang, P.; Kriechbaumer, V.; Tilsner, J.; Frigerio, L.; Sparkes, I.; Hawes, C.; Oparka, K. Putting the Squeeze on Plasmodesmata: A Role for Reticulons in Primary Plasmodesmata Formation. Plant Physiol. 2015, 168, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Kriechbaumer, V.; Botchway, S.W.; Slade, S.E.; Knox, K.; Frigerio, L.; Oparka, K.; Hawes, C. Reticulomics: Protein-Protein Interaction Studies with Two Plasmodesmata-Localized Reticulon Family Proteins Identify Binding Partners Enriched at Plasmodesmata, Endoplasmic Reticulum, and the Plasma Membrane. Plant Physiol. 2015, 169, 1933–1945. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, M. Plasmodesmata: Channels for viruses on the move. Methods Mol. Biol. 2015, 1217, 25–52. [Google Scholar] [PubMed]
- Amari, K.; Di Donato, M.; Dolja, V.V.; Heinlein, M. Myosins VIII and XI play distinct roles in reproduction and transport of tobacco mosaic virus. PLoS Pathog. 2014, 10, e1004448. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, J.; Ursache, R.; Campilho, A.; Miyashima, S.; Belevich, I.; O’Regan, S.; Mullendore, D.L.; Yadav, S.R.; Lanz, C.; Beverina, L.; et al. CHOLINE TRANSPORTER-LIKE1 is required for sieve plate development to mediate long-distance cell-to-cell communication. Nat. Commun. 2014, 5, 4276. [Google Scholar] [CrossRef] [PubMed]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R.; et al. Callose biosynthesis regulates symplastic trafficking during root development. Dev. Cell. 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.K.; Lappalainen, P. Filopodia: Molecular architecture and cellular functions. Nat. Rev. Mol. Cell Biol. 2008, 9, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Delage, E.; Cervantes, D.C.; Pénard, E.; Schmitt, C.; Syan, S.; Disanza, A.; Scita, G.; Zurzolo, C. Differential identity of Filopodia and Tunneling Nanotubes revealed by the opposite functions of actin regulatory complexes. Sci. Rep. 2016, 6, 39632. [Google Scholar] [CrossRef] [PubMed]
- Abounit, S.; Bousset, L.; Loria, F.; Zhu, S.; de Chaumont, F.; Pieri, L.; Olivo-Marin, J.C.; Melki, R.; Zurzolo, C. Tunneling nanotubes spread fibrillar α-synuclein by intercellular trafficking of lysosomes. EMBO J. 2016, 35, 2120–2138. [Google Scholar] [CrossRef] [PubMed]
- Bodemann, B.O.; Orvedahl, A.; Chenget, T.; Ram, R.R.; Ou, Y.H.; Formstecher, E.; Maiti, M.; Hazelett, C.C.; Wauson, E.M.; Balakireva, M.; et al. RalB and the exocyst mediate the cellular starvation response by direct activation of autophagosome assembly. Cell 2011, 144, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Pecenková, T.; Hála, M.; Kulich, I.; Kocourková, D.; Drdová, E.; Fendrych, M.; Toupalová, H.; Žársky, V. The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction. J. Exp. Bot. 2011, 62, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.; Raikhel, N.V.; Hicks, G.R. Powerful partners: Arabidopsis and chemical genomics. Arabidopsis Book 2009, 7, e0109. [Google Scholar] [CrossRef]
- Zhang, C.; Brown, M.Q.; van de Ven, W.; Zhang, Z.M.; Wu, B.; Young, M.C.; Synek, L.; Borchardt, D.; Harrison, R.; Pan, S.; et al. Endosidin2 targets conserved exocyst complex subunit EXO70 to inhibit exocytosis. Proc. Natl. Acad. Sci. USA 2016, 113, E41–E50. [Google Scholar] [CrossRef] [PubMed]
- Di Sansebastiano, G.P. Defining new SNARE functions: The i-SNARE. Front. Plant Sci. 2013, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Ginefra, P.; Mastrodonato, V.; Beznoussenko, G.V.; Rusten, T.E.; Bilder, D.; Stenmark, H.; Mironov, A.A.; Vaccari, T. Multiple functions of the SNARE protein Snap29 in autophagy, endocytic, and exocytic trafficking during epithelial formation in Drosophila. Autophagy 2014, 10, 2251–2268. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Mastrodonato, V.; Beznoussenko, G.V.; Mironov, A.A.; Tognon, E.; Vaccari, T. An essential step of kinetochore formation controlled by the SNARE protein Snap29. EMBO J. 2016, 35, 2223–2237. [Google Scholar] [CrossRef] [PubMed]
- Faraco, M.; Latino, A.; de Benedictis, M.; Piro, G.; Di Sansebastiano, G.P. Atsyp51 and Atsyp52 differently affect sorting of Gfpchi and Aleugfp vacuolar markers. J. Plant Biochem. Physiol. 2013, 1, e105. [Google Scholar]
- Borg, M.; Bakke, O.; Progida, C. A novel interaction between Rab7b and actomyosin reveals a dual role in intracellular transport and cell migration. J. Cell Sci. 2014, 127, 4927–4939. [Google Scholar] [CrossRef] [PubMed]
- Margiotta, A.; Progida, C.; Bakke, O.; Bucci, C. Rab7a regulates cell migration through Rac1 and vimentin. Biochim. Biophys. Acta 2016, 1864, 367–381. [Google Scholar] [CrossRef]
- Heard, W.; Sklenar, J.; Tome, D.F.; Robatzek, S.; Jones, A.M. Identification of regulatory and cargo proteins of endosomal and secretory pathways in Arabidopsis thaliana by proteomic dissection. Mol. Cell. Prot. 2015, 14, 1796–1813. [Google Scholar] [CrossRef] [PubMed]
- Sechet, J.; Frey, A.; Effroy-Cuzzi, D.; Berger, A.; Perreau, F.; Cueff, G.; Charif, D.; Rajjou, L.; Mouille, G.; North, H.M.; et al. Xyloglucan metabolism differentially impacts the cell wall characteristics of the endosperm and embryo during Arabidopsis seed germination. Plant Phys. 2016, 170, 1367–1380. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, C.P.; Dilokpimol, A.; Mouille, G.; Burow, M.; Geshi, N. Arabinogalactan glycosyltransferases target to a unique subcellular compartment that may function in unconventional secretion in plants. Traffic 2014, 15, 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Landsverk, O.J.; Barois, N.; Gregers, T.F.; Bakke, O. Invariant chain increases the half-life of MHC II by delaying endosomal maturation. Immunol. Cell Biol. 2011, 89, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Safavian, D.; Zayed, Y.; Indriolo, E.; Chapman, L.; Ahmed, A.; Goring, D.R. RNA Silencing of Exocyst Genes in the Stigma Impairs the Acceptance of Compatible Pollen in Arabidopsis. Plant. Phys. 2015, 169, 2526–2538. [Google Scholar] [CrossRef] [PubMed]
- Doucet, J.; Lee, H.K.; Goring, D.R. Pollen acceptance or rejection: A tale of two pathways. Trends Plant Sci. 2016, 21, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Hafidh, S.; Potěšil, D.; Fíla, J.; Čapková, V.; Zdráhal, Z.; Honys, D. Quantitative proteomics of the tobacco pollen tube secretome identifies novel pollen tube guidance proteins important for fertilization. Genome Biol. 2016, 17, 81. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.P.; Zeng, Y.; Jiang, L. COPII paralogs in plants—Functional redundancy or diversity? Trends Plant Sci. 2016, 21, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Shen, J.; Gao, C.; Zhuang, X.; Wang, J.; Jiang, L. Biogenesis of Plant Prevacuolar Multivesicular Bodies. Mol. Plant 2016, 9, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Bolter, B.; Soll, J. Once upon a time—Chloroplast protein import research from infancy to future challenges. Mol. Plant 2016, 9, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Vothknecht, U.C.; Soll, J. Chloroplast membrane transport: Interplay of prokaryotic and eukaryotic traits. Gene 2005, 354, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Cabanillas, D.G.; Zheng, H.; Laliberté, J.F. Turnip mosaic virus moves systemically through both phloem and xylem as membrane-associated complexes. Plant Physiol. 2015, 167, 1374–1388. [Google Scholar] [CrossRef] [PubMed]
- Saplaoura, E.; Kragler, F. Mobile Transcripts and Intercellular Communication in Plants. Enzymes 2016, 40, 1–29. [Google Scholar] [PubMed]
- Crippa, V.; D’Agostino, V.G.; Cristofani, R.; Rusmini, P.; Cicardi, M.E.; Messi, E.; Loffredo, R.; Pancher, M.; Piccolella, M.; Galbiati, M.; et al. Transcriptional induction of the heat shock protein B8 mediates the clearance of misfolded proteins responsible for motor neuron diseases. Sci. Rep. 2016, 10, 22827. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Shinohara, H.; Sakagami, Y.; Matsubayashi, Y. Arabidopsis CLV3 peptide directly binds CLV1 ectodomain. Science 2008, 319, 294. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Cornish, K.; Xie, W. Natural rubber biosynthesis in plants: Rubber transferase. Methods Enzymol. 2012, 515, 63–82. [Google Scholar] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompa, A.; De Marchis, F.; Pallotta, M.T.; Benitez-Alfonso, Y.; Jones, A.; Schipper, K.; Moreau, K.; Žárský, V.; Di Sansebastiano, G.P.; Bellucci, M. Unconventional Transport Routes of Soluble and Membrane Proteins and Their Role in Developmental Biology. Int. J. Mol. Sci. 2017, 18, 703. https://doi.org/10.3390/ijms18040703
Pompa A, De Marchis F, Pallotta MT, Benitez-Alfonso Y, Jones A, Schipper K, Moreau K, Žárský V, Di Sansebastiano GP, Bellucci M. Unconventional Transport Routes of Soluble and Membrane Proteins and Their Role in Developmental Biology. International Journal of Molecular Sciences. 2017; 18(4):703. https://doi.org/10.3390/ijms18040703
Chicago/Turabian StylePompa, Andrea, Francesca De Marchis, Maria Teresa Pallotta, Yoselin Benitez-Alfonso, Alexandra Jones, Kerstin Schipper, Kevin Moreau, Viktor Žárský, Gian Pietro Di Sansebastiano, and Michele Bellucci. 2017. "Unconventional Transport Routes of Soluble and Membrane Proteins and Their Role in Developmental Biology" International Journal of Molecular Sciences 18, no. 4: 703. https://doi.org/10.3390/ijms18040703
APA StylePompa, A., De Marchis, F., Pallotta, M. T., Benitez-Alfonso, Y., Jones, A., Schipper, K., Moreau, K., Žárský, V., Di Sansebastiano, G. P., & Bellucci, M. (2017). Unconventional Transport Routes of Soluble and Membrane Proteins and Their Role in Developmental Biology. International Journal of Molecular Sciences, 18(4), 703. https://doi.org/10.3390/ijms18040703