
Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo

Abstract
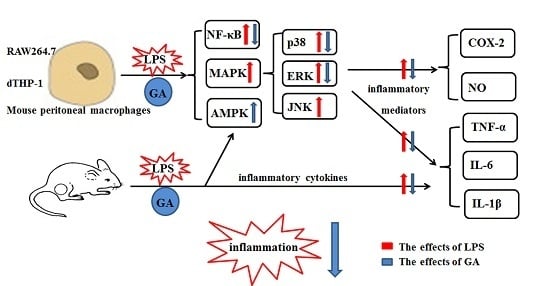
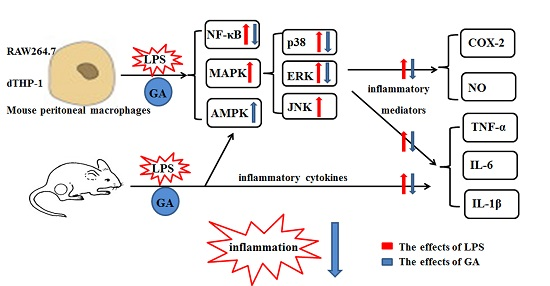
:
1. Introduction
2. Results
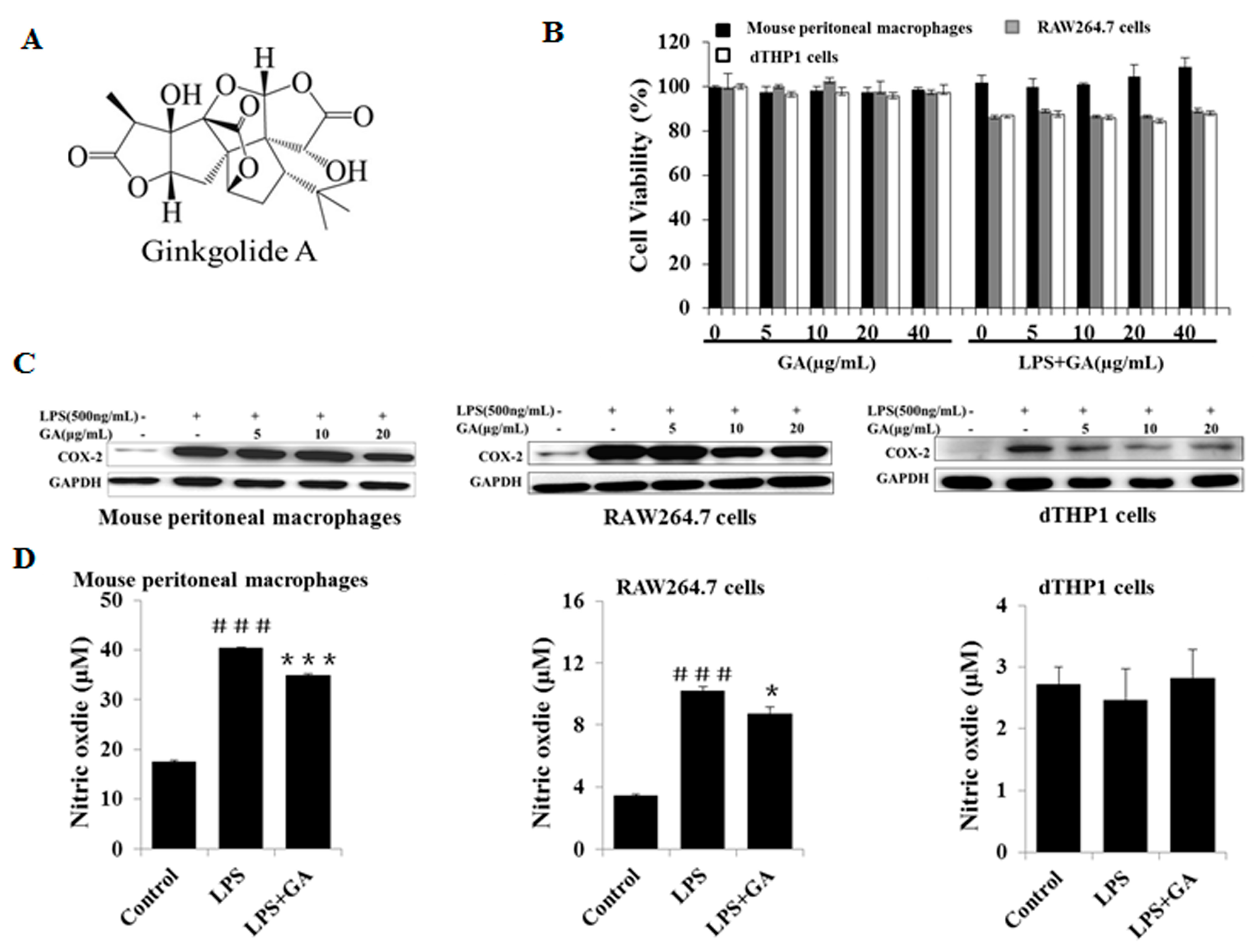
2.1. GA Inhibits LPS-Stimulated COX-2 Expression and NO Production
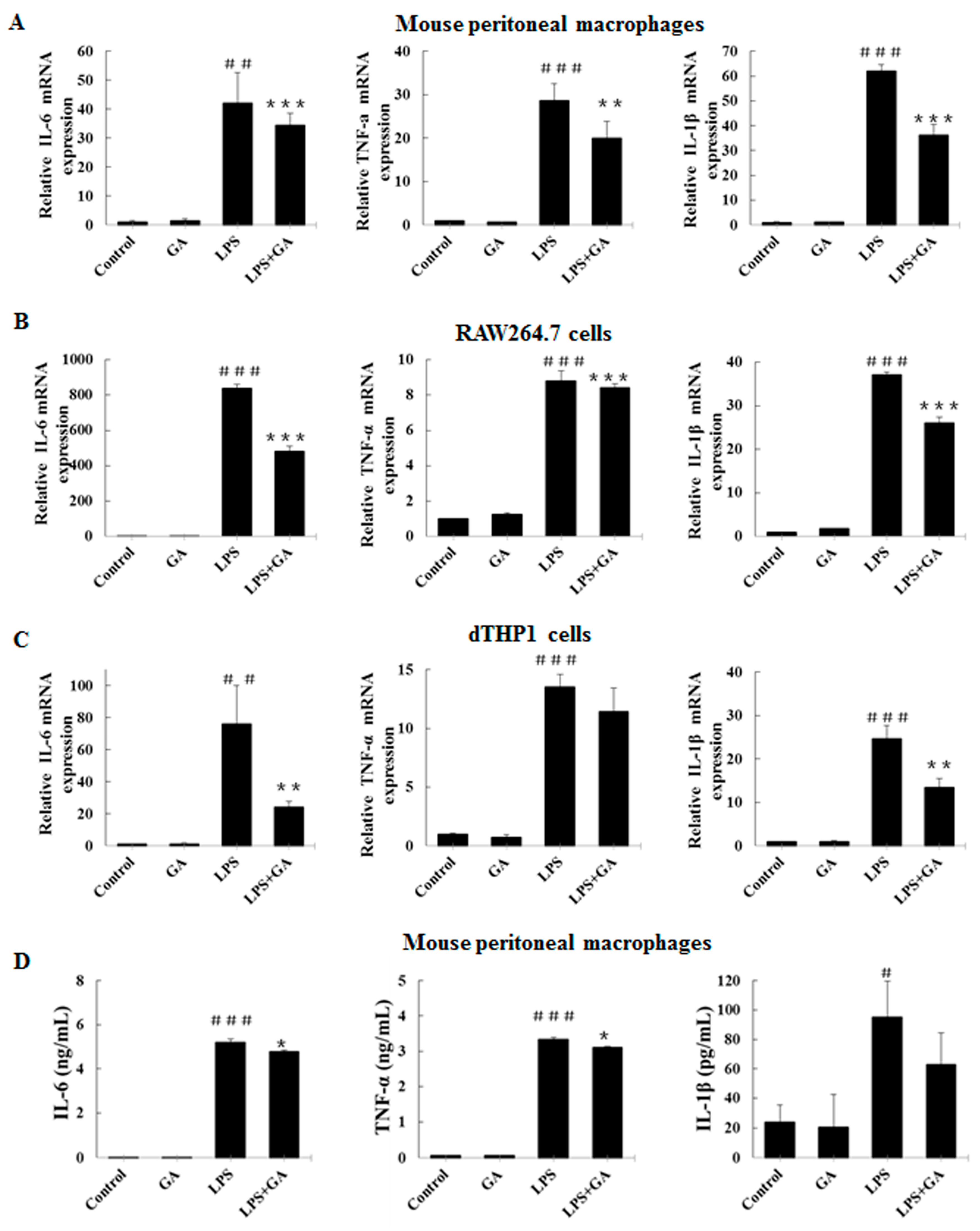
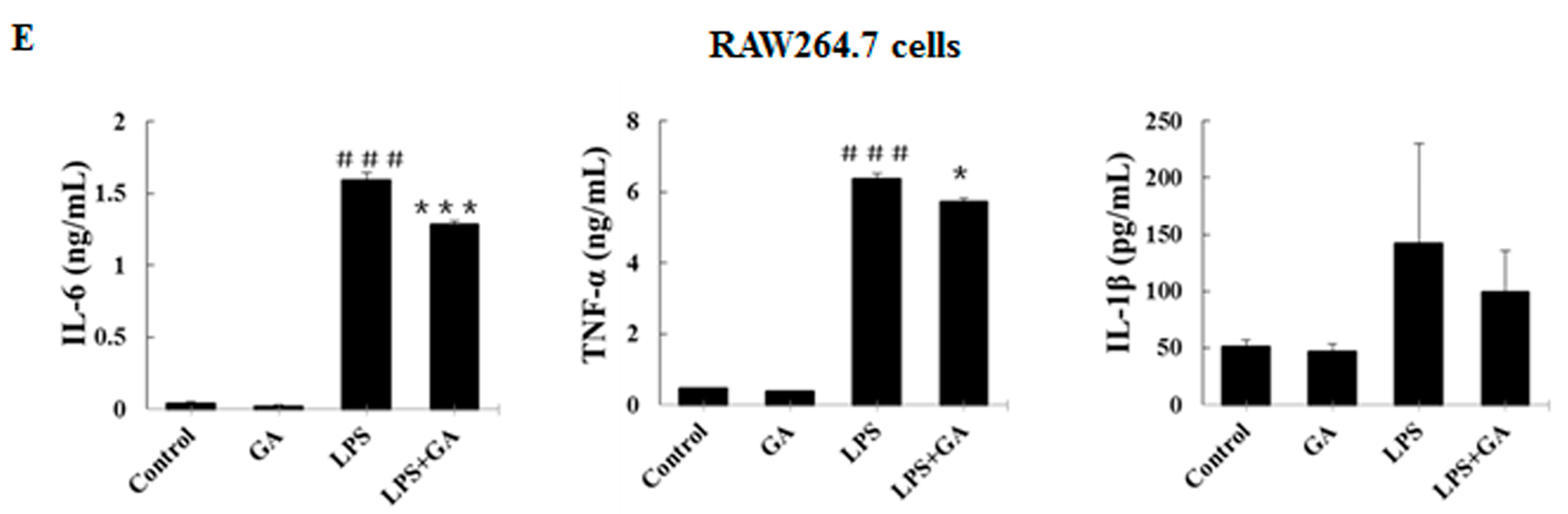
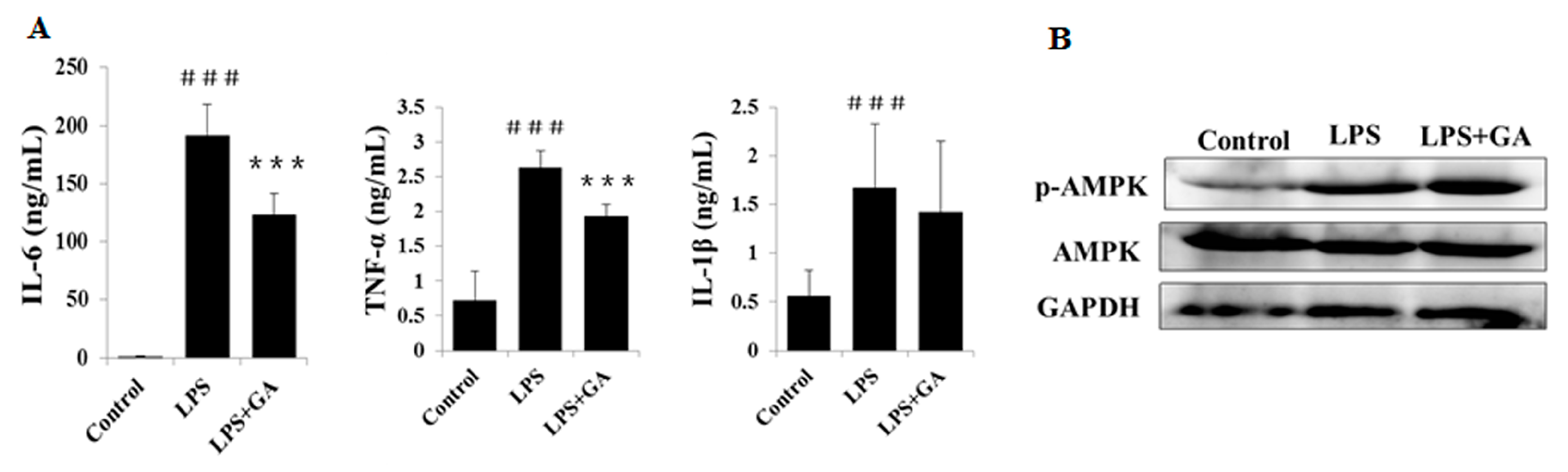
2.2. GA Inhibits LPS-Stimulated Expression and Production of Pro-Inflammatory Cytokines in Macrophages
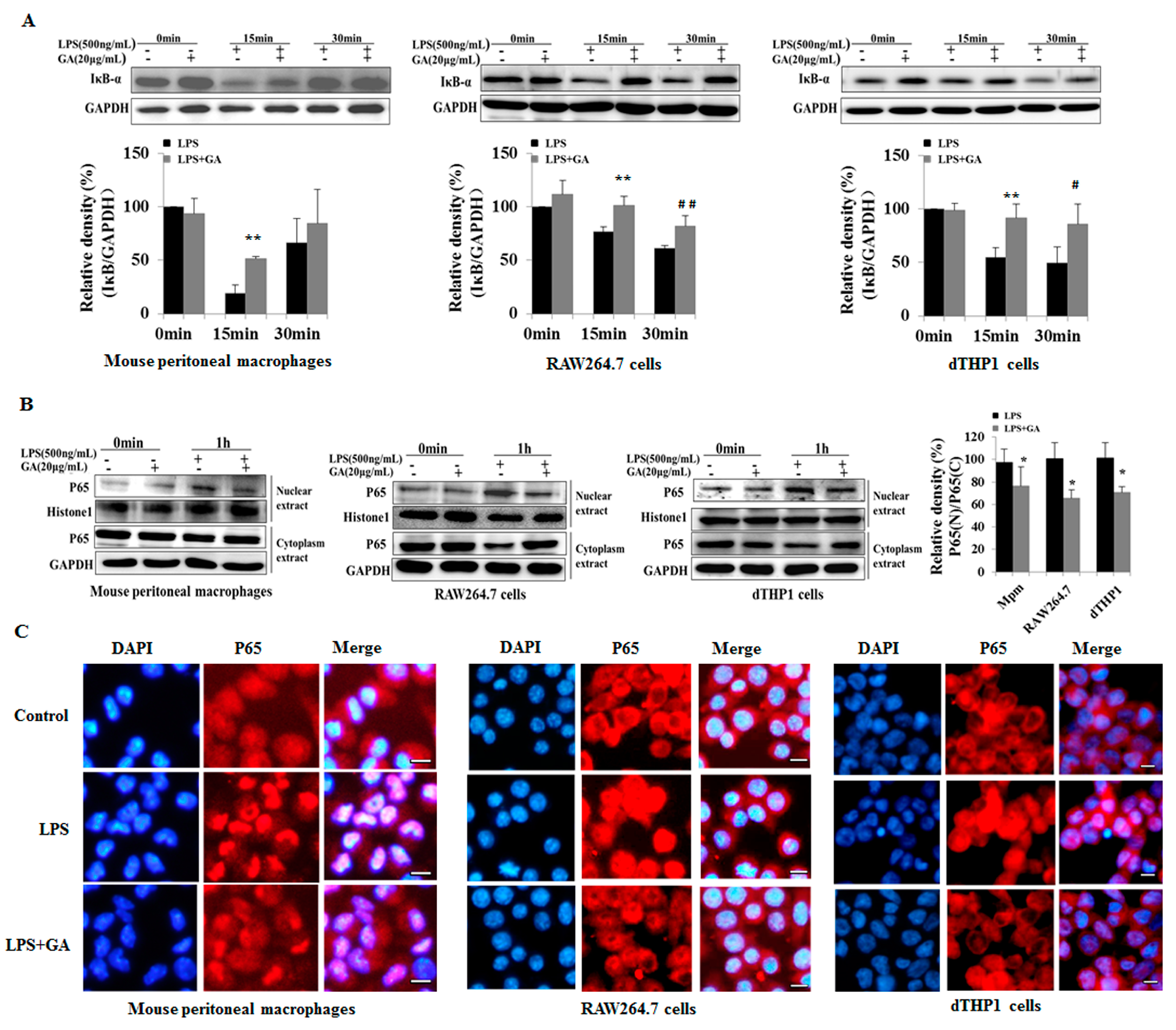
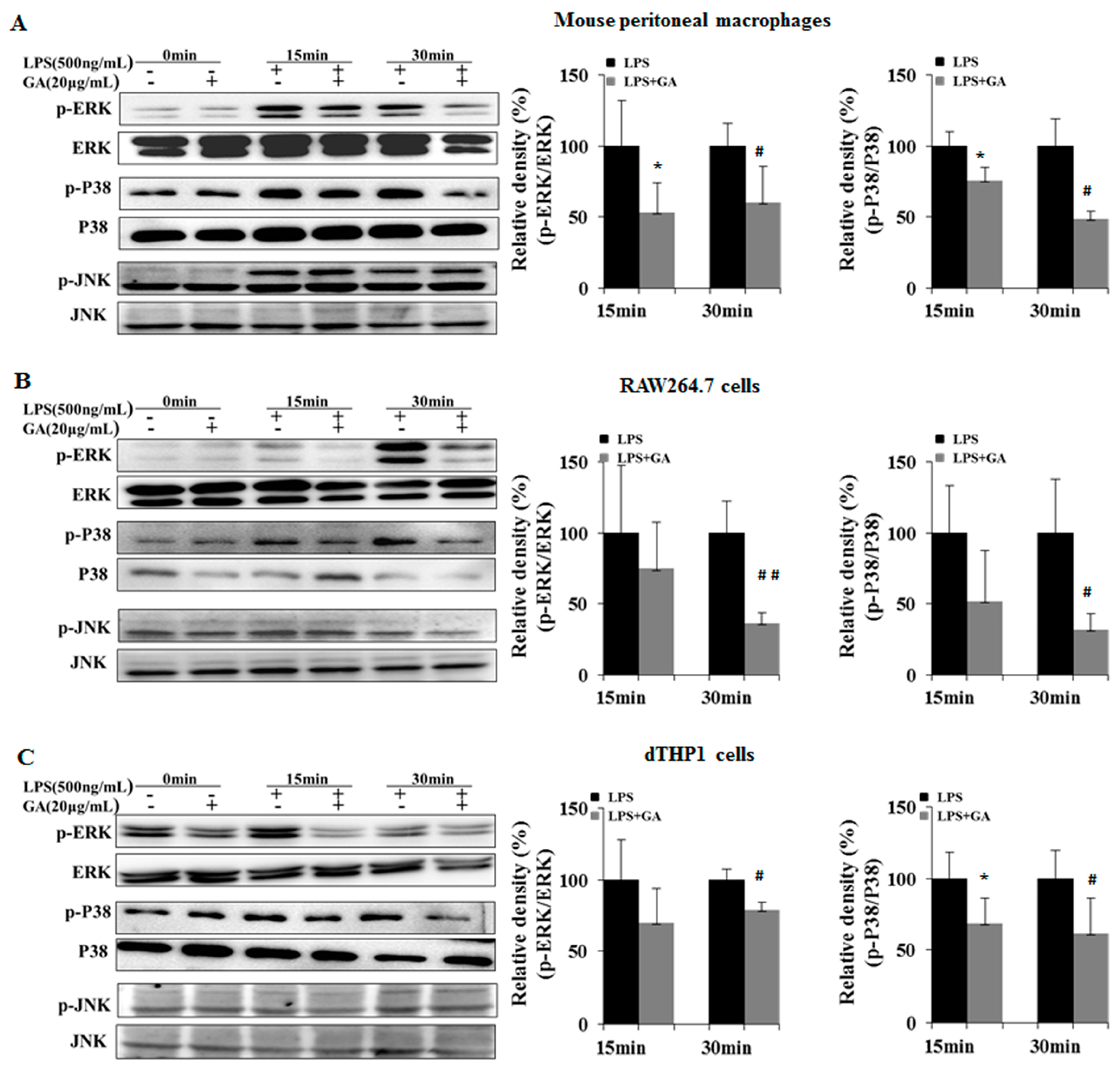
2.3. GA Suppresses LPS-Induced Activation of NF-κB and MAPK Signaling
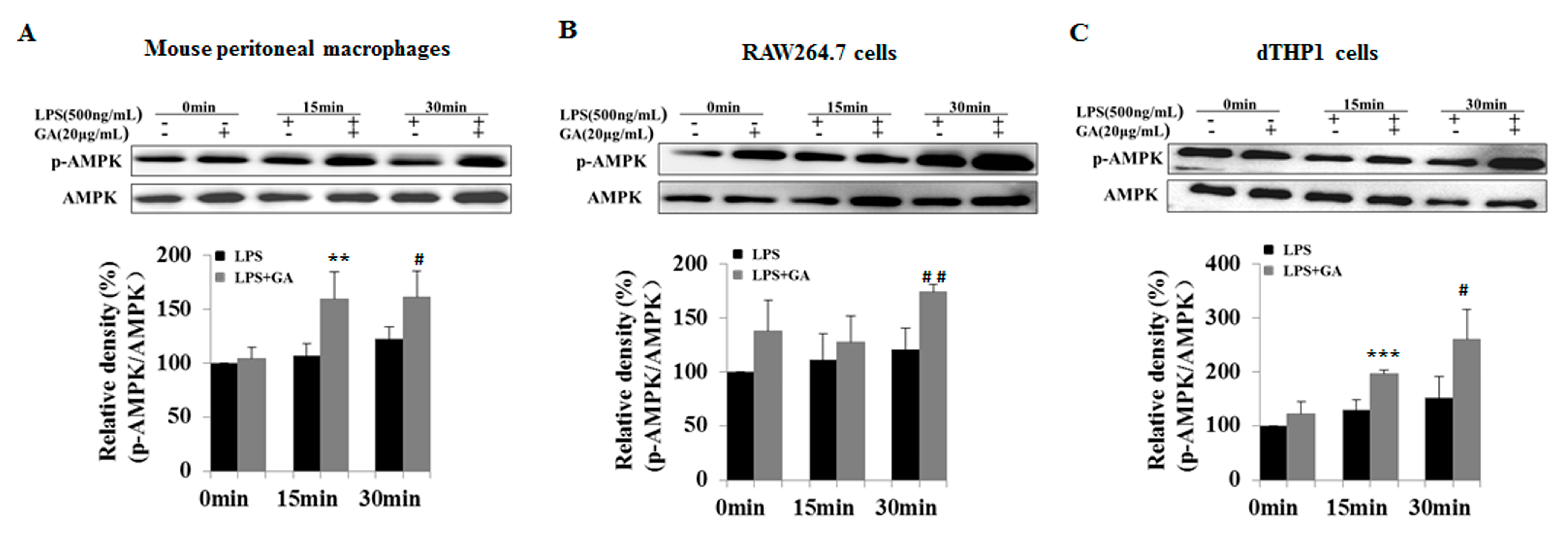
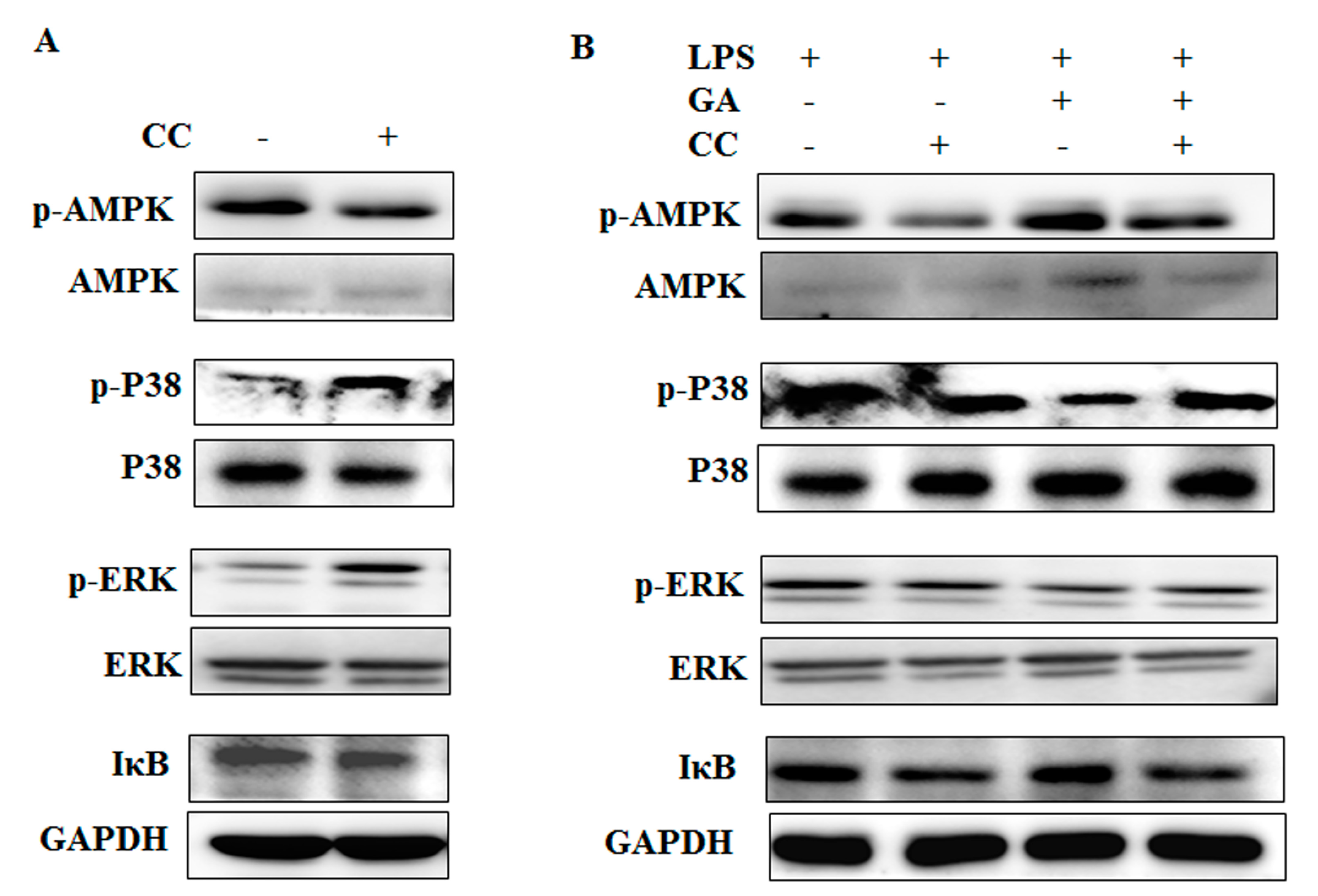
2.4. GA Upregulates AMPK Signaling
2.5. GA Inhibits NF-κB and MAPks Pathway through AMPK Activation
2.6. GA Attenuates the Inflammatory Response in LPS-Treated BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Reagents and Antibodies
4.3. Cell Culture
4.4. Cell Viability
4.5. RT-qPCR
4.6. ELISA
4.7. NO Assay
4.8. Western Blot Analysis
4.9. Fluorescence Microscopy for Visualization of NF-κB p65 Localization
4.10. LPS-Induced Mice
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| COX-2 | Cyclo-oxygenase-2 |
| CC | Compound C |
| ERK | Extracellular regulated kinases |
| GA | Ginkgolide A |
| JNK | c-Jun N-terminal kinases |
| LPS | Lipopolysaccharide |
| MAPKs | Mitogen-activated protein kinases |
| NO | Nitric oxide |
| NF-κB | Nuclear factor kappa-B |
References
- Arulselvan, P.; Tan, W.S.; Gothai, S.; Muniandy, K.; Fakurazi, S.; Esa, N.M.; Alarfaj, A.A.; Kumar, S.S. Anti-inflammatory potential of ethyl acetate fraction of moringa oleifera in downregulating the NF-kappaB signaling pathway in lipopolysaccharide-stimulated macrophages. Molecules 2016, 21, 1452. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Y.; Jeon, B.H.; Kim, Y.C.; Lee, S.H.; Sohn, D.H.; Seo, G.S. PF2401-SF, standardized fraction of Salvia miltiorrhiza shows anti-inflammatory activity in macrophages and acute arthritis in vivo. Int. Immunopharmacol. 2013, 16, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Wu, C.; Zhao, W.; Zhang, X.; Chen, X. Neocryptotanshinone inhibits lipopolysaccharide-induced inflammation in RAW264.7 macrophages by suppression of NF-κB and iNOS signaling pathways. Acta Pharm. Sin. B 2015, 5, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Guo, H.; Li, Y.; Meng, X.; Yan, L.; Zhang, D.; Wu, S.; Zhou, H.; Peng, L.; Xie, Q.; et al. Oleoylethanolamide exerts anti-inflammatory effects on LPS-induced THP-1 cells by enhancing PPARα signaling and inhibiting the NF-κB and ERK1/2/AP-1/STAT3 pathways. Sci. Rep. 2016, 6, 34611. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.H.; Oh, Y.C.; Cho, W.K.; Yim, N.H.; Ma, J.Y. Anti-Inflammatory effect of rhapontici radix ethanol extract via inhibition of NF-κB and MAPK and induction of HO-1 in macrophages. Mediat. Inflamm. 2016, 2016, 7216912. [Google Scholar] [CrossRef] [PubMed]
- Angus, D.C.; Linde-Zwirble, W.T.; Lidicker, J.; Clermont, G.; Carcillo, J.; Pinsky, M.R. Epidemiology of severe sepsis in the United States: Analysis of incidence, outcome, and associated costs of care. Crit. Care Med. 2001, 29, 1303–13010. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Chen, L.; Bi, C.; Zhang, M.; Jiao, W.; Yao, X. In vitro anti-inflammatory effect of picrasmalignan A by the inhibition of iNOS and COX2 expression in LPS activated macrophage RAW264.7 cells. Mol. Med. Rep. 2013, 8, 1575–1579. [Google Scholar] [PubMed]
- Shin, J.S.; Hong, Y.; Lee, H.H.; Ryu, B.; Cho, Y.W.; Kim, N.J.; Jang, D.S.; Lee, K.T. Fulgidic Acid isolated from the rhizomes of cyperus rotundus suppresses LPS-Induced iNOS, COX-2, TNF-α, and IL-6 expression by AP-1 inactivation in RAW264.7 macrophages. Biol. Pharm. Bull. 2015, 38, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.L.; Tong, Q.; Wang, W.Q.; Shi, C.Y.; Xiong, W.; Chen, J.; Liu, X.; Fang, J.G. Suppression of inflammatory responses by dihydromyricetin, a flavonoid from ampelopsis grossedentata, via inhibiting the activation of NF-κB and MAPK signaling pathways. J. Nat. Prod. 2015, 78, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Kuribara, H.; Weintraub, S.T.; Yoshihama, T.; Maruyama, Y. An anxiolytic-like effect of Ginkgo biloba extract and its constituent, ginkgolide-A, in mice. J. Nat. Prod. 2003, 66, 1333–1337. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.; Wang, H.; Hong, J.; Zhang, T.; Lin, C.; Meng, C. PXR mediated protection against liver inflammation by Ginkgolide A in tetrachloromethane treated mice. Biomol. Ther. 2016, 24, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Gao, C.; Cui, Z. Ginkgolide A reduces inflammatory response in high-glucose-stimulated human umbilical vein endothelial cells through STAT3-mediated pathway. Int. Immunopharmacol. 2015, 25, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.H.; Ge, J.B.; Li, M.; Wu, F.; Zhang, W.; Qin, Z.H. Inhibition of NF-κB activation is associated with anti-inflammatory and anti-apoptotic effects of Ginkgolide B in a mouse model of cerebral ischemia/reperfusion injury. Eur. J. Pharm. Sci. 2012, 47, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.L.; Liou, C.J.; Li, Z.Y.; Lai, X.Y.; Fang, L.W.; Huang, W.C. Sesamol suppresses the inflammatory response by inhibiting NF-κB/MAPK activation and upregulating AMP kinase signaling in RAW264.7 macrophages. Inflamm. Res. 2015, 64, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.; Kikuchi, K.; Ito, T.; Tokuda, M.; Matsuyama, T.; Noma, S.; Hashiguchi, T.; Torii, M.; Maruyama, I.; Kawahara, K. MK615 attenuates Porphyromonas gingivalis lipopolysaccharide-induced pro-inflammatory cytokine release via MAPK inactivation in murine macrophage-like RAW264.7 cells. Biochem. Biophys. Res. Commun. 2009, 389, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zmijewski, J.W.; Lorne, E.; Liu, G.; Park, Y.J.; Tsuruta, Y.; Abraham, E. Activation of AMPK attenuates neutrophil proinflammatory activity and decreases the severity of acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L497–L504. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jin, M.L.; Ko, M.J.; Park, G.; Choi, Y.W. Anti-neuroinflammatory effect of emodin in LPS-stimulated microglia: Involvement of AMPK/Nrf2 activation. Neurochem. Res. 2016, 41, 2981–2992. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.P.; Lin, C.H.; Chen, H.M.; Lin, J.T.; Cheng, Y.F.; Kao, S.H. AMPK activation inhibits expression of proinflammatory mediators through downregulation of PI3K/p38 MAPK and NF-κB signaling in murine macrophages. DNA Cell Biol. 2015, 34, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.K.; Raposo, C.; Rocha, S.W.; Barbosa, K.P.; Luna, R.L.; da Cruz-Hofling, M.A.; Peixoto, C.A. Involvement of AMPK, IKβα-NFκB and eNOS in the sildenafil anti-inflammatory mechanism in a demyelination model. Brain Res. 2015, 1627, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Ghandadi, M.; Sahebkar, A. Curcumin: An effective inhibitor of interleukin-6. Curr. Pharm. Des. 2016. [Google Scholar] [CrossRef]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A review: Inflammatory process in Alzheimer’s disease, role of cytokines. Sci. World J. 2012, 2012, 756357. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.A.; Chien, S.Y.; Chan, Y.L.; Lu, M.K.; Wu, C.H.; Kong, Z.L.; Wu, C.J. Inhibition of Lipopolysaccharide (LPS)-induced inflammatory responses by Sargassum hemiphyllum sulfated polysaccharide extract in RAW264.7 macrophage cells. J. Agric. Food Chem. 2011, 59, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Brooks, R.; Houskeeper, J.; Bremner, S.K.; Dunlop, J.; Viollet, B.; Logan, P.J.; Salt, I.P.; Ahmed, S.F.; Yarwood, S.J. Metformin suppresses adipogenesis through both AMP-activated protein kinase (AMPK)-dependent and AMPK-independent mechanisms. Mol. Cell Endocrinol. 2016, 440, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xin, H.; Tang, W.; Li, Y.; Zhang, Z.; Fan, L.; Miao, L.; Tan, B.; Wang, X.; Zhu, Y. AMPK serves as a therapeutic target against anemia of inflammation. Antioxid. Redox Signal. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.C.; Duke, R.K.; Hinton, T.; Johnston, G.A. GABAA receptor cysteinyl mutants and the ginkgo terpenoid lactones bilobalide and ginkgolides. Eur. J. Pharmacol. 2016, 777, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, G.; Yan, Y.; Bao, L.; Chen, B.; Qi, R. Ginkgolide B reduces atherogenesis and vascular inflammation in ApoE(-/-) mice. PLoS ONE 2012, 7, e36237. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhang, D.M.; Song, Z.B.; Hou, Y.Q.; Bao, Y.L.; Sun, L.G.; Yu, C.L.; Li, Y.X. Protumoral TSP50 regulates macrophage activities and polarization via production of TNF-α and IL-1β, and activation of the NF-κB signaling pathway. PLoS ONE 2015, 10, e0145095. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IL-6(M) | 5′-TCCAGTTGCCTTCTTGGGAC-3′ | 5′-GTGTAATTAAGCCTCCGACTTG-3′ |
| IL-6(H) | 5′-GTGAAAGCAGCAAAGAGGCACT-3′ | 5′-ACCAGGCAAGTCTCCTCATTGA-3′ |
| TNF-α(M) | 5′-GACCCTCACACTCAGATCATCTTCT-3′ | 5′-CCTCCACTTGGTGGTTTGCT-3′ |
| TNF-α(H) | 5′-TCAATCGGCCCGACTATCTC-3′ | 5′-CAGGGCAATGATCCCAAAGT-3′ |
| IL–1β(M) | 5′-TCGTGCTGTCGGACCCATAT-3′ | 5′-GTCGTTGCTTGGTTCTCCTTGT-3′ |
| IL–1β(H) | 5′-GCACGATGCACCTGTACGAT-3′ | 5′-AGACATCACCAAGCTTTTTTGCT-3′ |
| GAPDH(M) | 5′-AAATGGTGAAGGTCGGTGTG-3′ | 5′-TGAAGGGGTCGTTGATGG-3′ |
| β-actin(H) | 5′-CGTGCGTGACATTAAGGAGAAG-3′ | 5′-GGAAGGAAGGCTGGAAGAGTG-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wu, Y.; Yao, X.; Hao, F.; Yu, C.; Bao, Y.; Wu, Y.; Song, Z.; Sun, Y.; Zheng, L.; et al. Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 794. https://doi.org/10.3390/ijms18040794
Li Y, Wu Y, Yao X, Hao F, Yu C, Bao Y, Wu Y, Song Z, Sun Y, Zheng L, et al. Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo. International Journal of Molecular Sciences. 2017; 18(4):794. https://doi.org/10.3390/ijms18040794
Chicago/Turabian StyleLi, Yan, Yannan Wu, Xinlei Yao, Fang Hao, Chunlei Yu, Yongli Bao, Yin Wu, Zhenbo Song, Ying Sun, Lihua Zheng, and et al. 2017. "Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo" International Journal of Molecular Sciences 18, no. 4: 794. https://doi.org/10.3390/ijms18040794
APA StyleLi, Y., Wu, Y., Yao, X., Hao, F., Yu, C., Bao, Y., Wu, Y., Song, Z., Sun, Y., Zheng, L., Wang, G., Huang, Y., Sun, L., & Li, Y. (2017). Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo. International Journal of Molecular Sciences, 18(4), 794. https://doi.org/10.3390/ijms18040794