
miR-103 Promotes Proliferation and Metastasis by Targeting KLF4 in Gastric Cancer

Abstract
:1. Introduction
2. Results
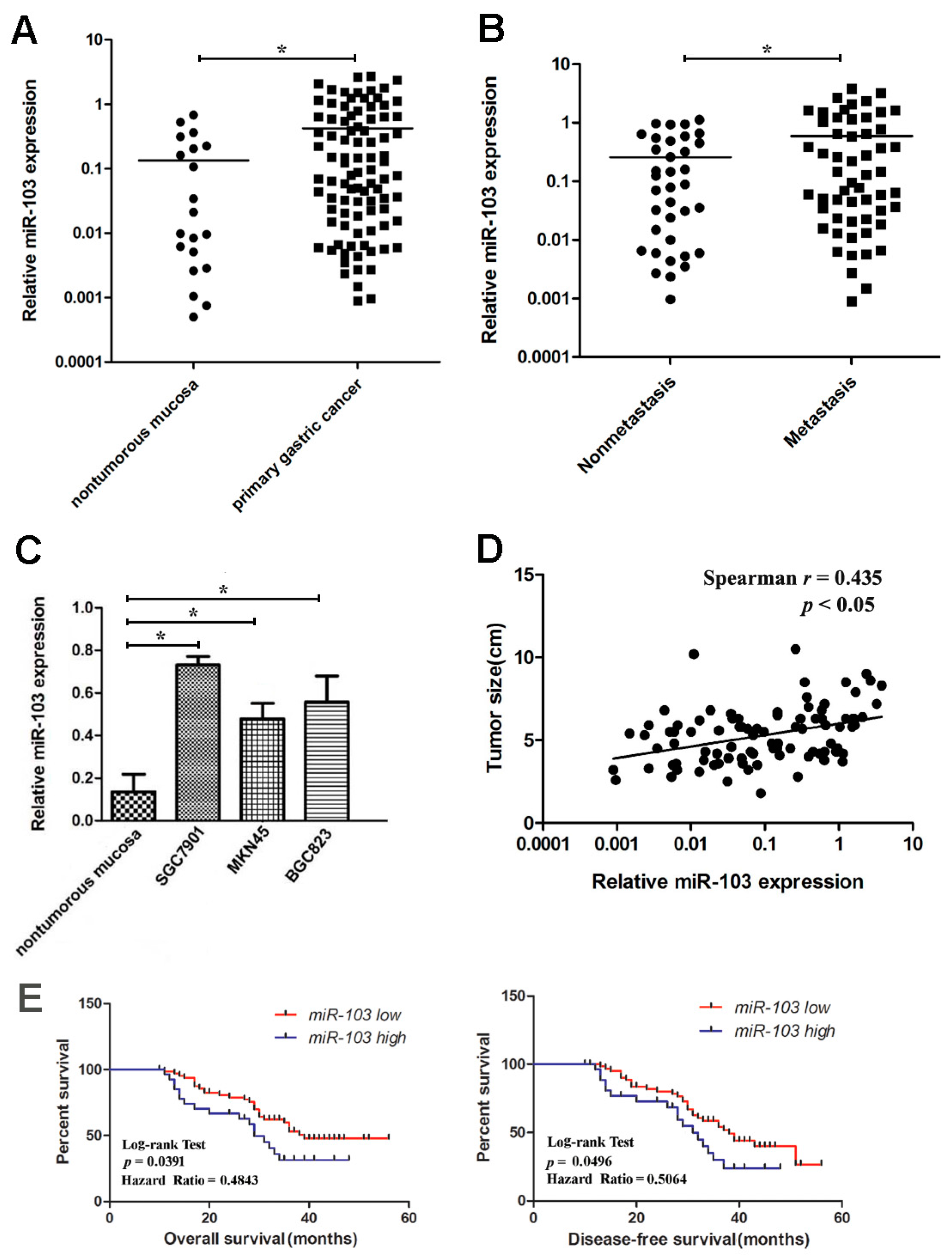
2.1. Upregulation of miR-103 in Gastric Cancer Tissues and Cell Lines
2.2. Overexpression of miR-103 Is Correlated with Poor Prognosis for Gastric Cancer
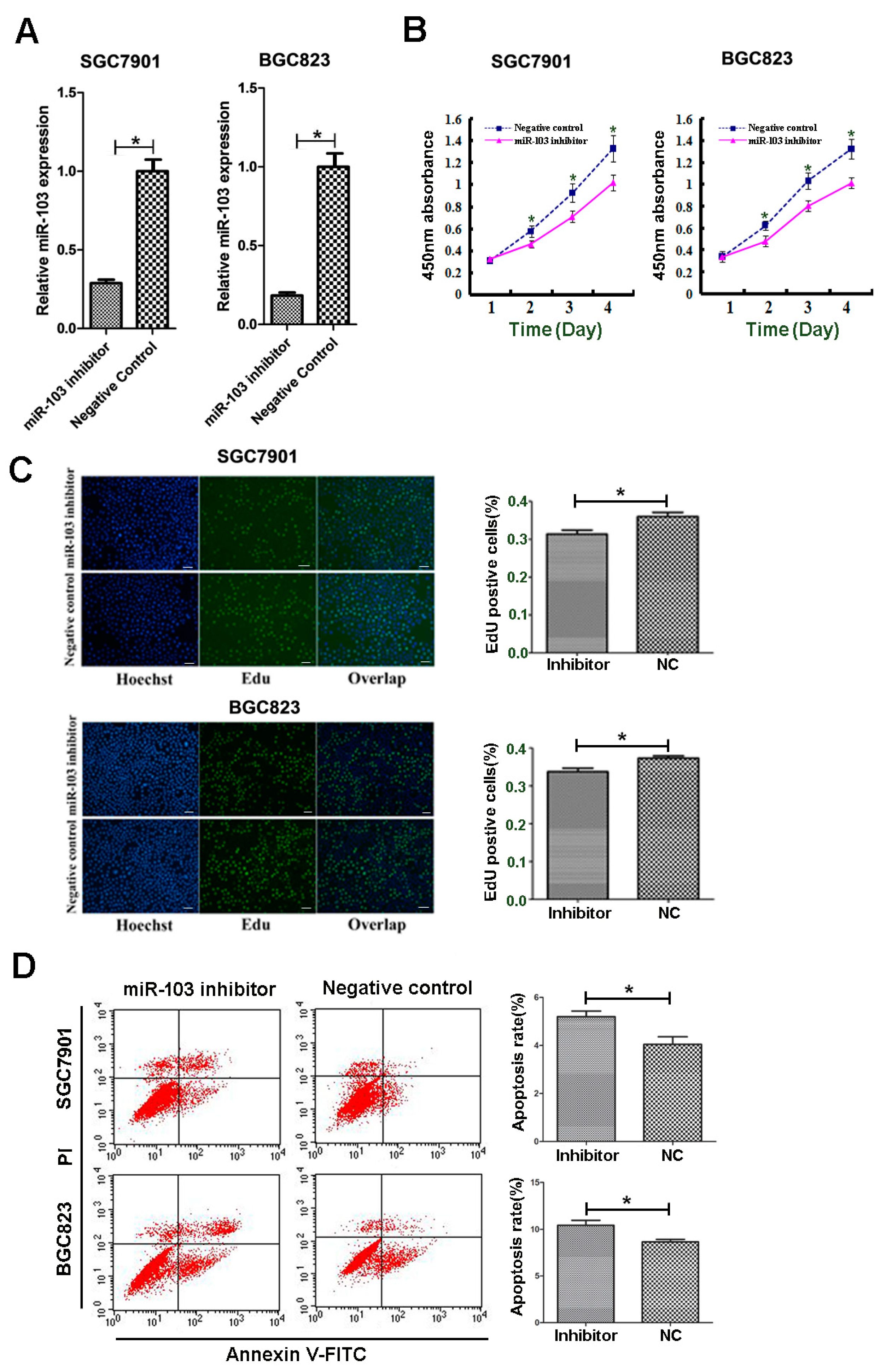
2.3. Downregulation of miR-103 Impaired Proliferation and Induced Apoptosis of SGC7901 and BGC823 Cells
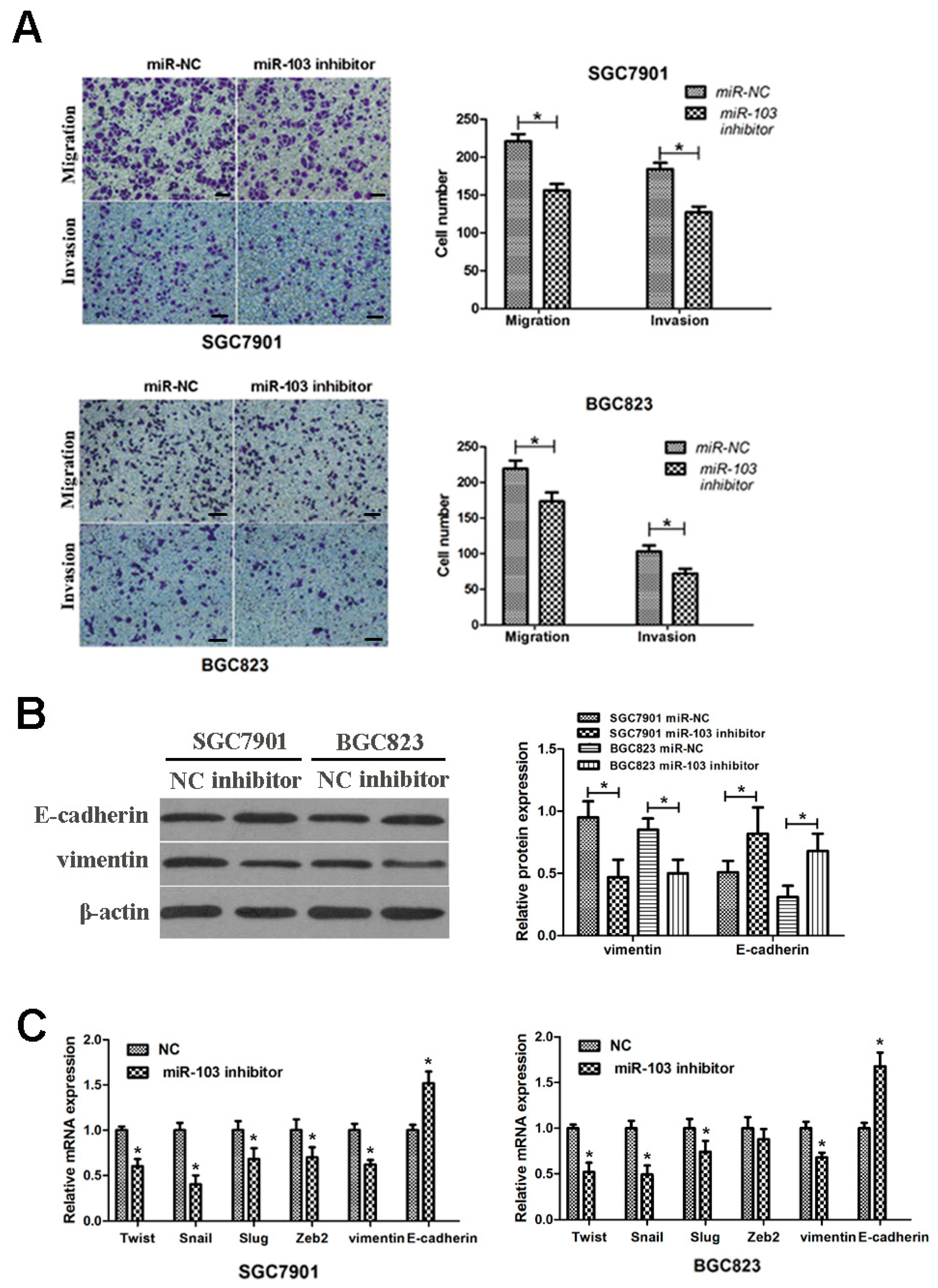
2.4. Reduction of miR-103 Inhibited GC Cells Migration and Invasion
2.5. Knockdown of miR-103 Suppressed the Epithelial–Mesenchymal Transition (EMT) Process of GC Cells
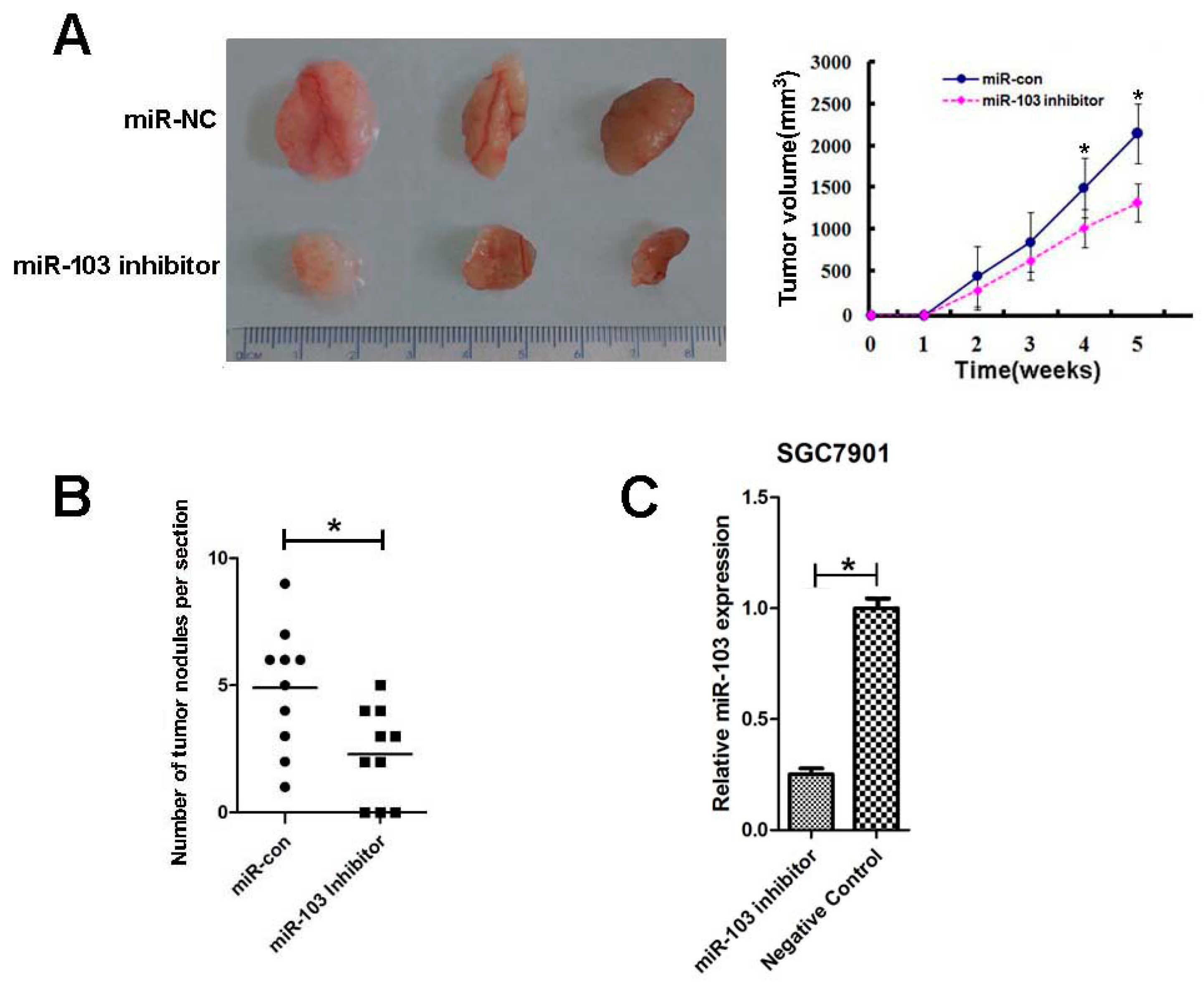
2.6. Downregulation of miR-103 Inhibited GC Growth and Lung Metastasis In Vivo
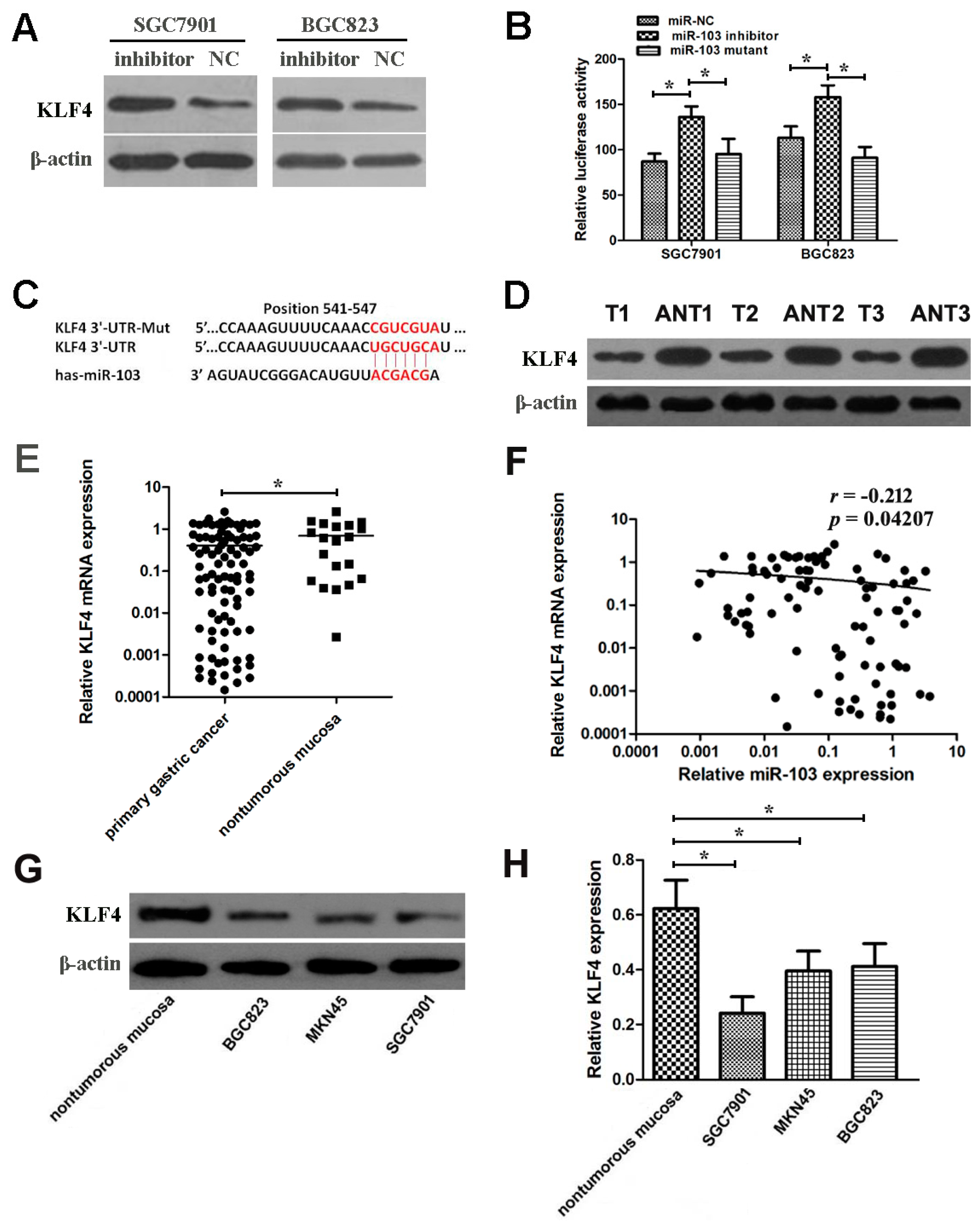
2.7. miR-103 Directly Targets and Down-Regulates KLF4 in GC Cells
2.8. miR-103 and KLF4 Are Clinically Relevant in Human GC Cell and Tissues
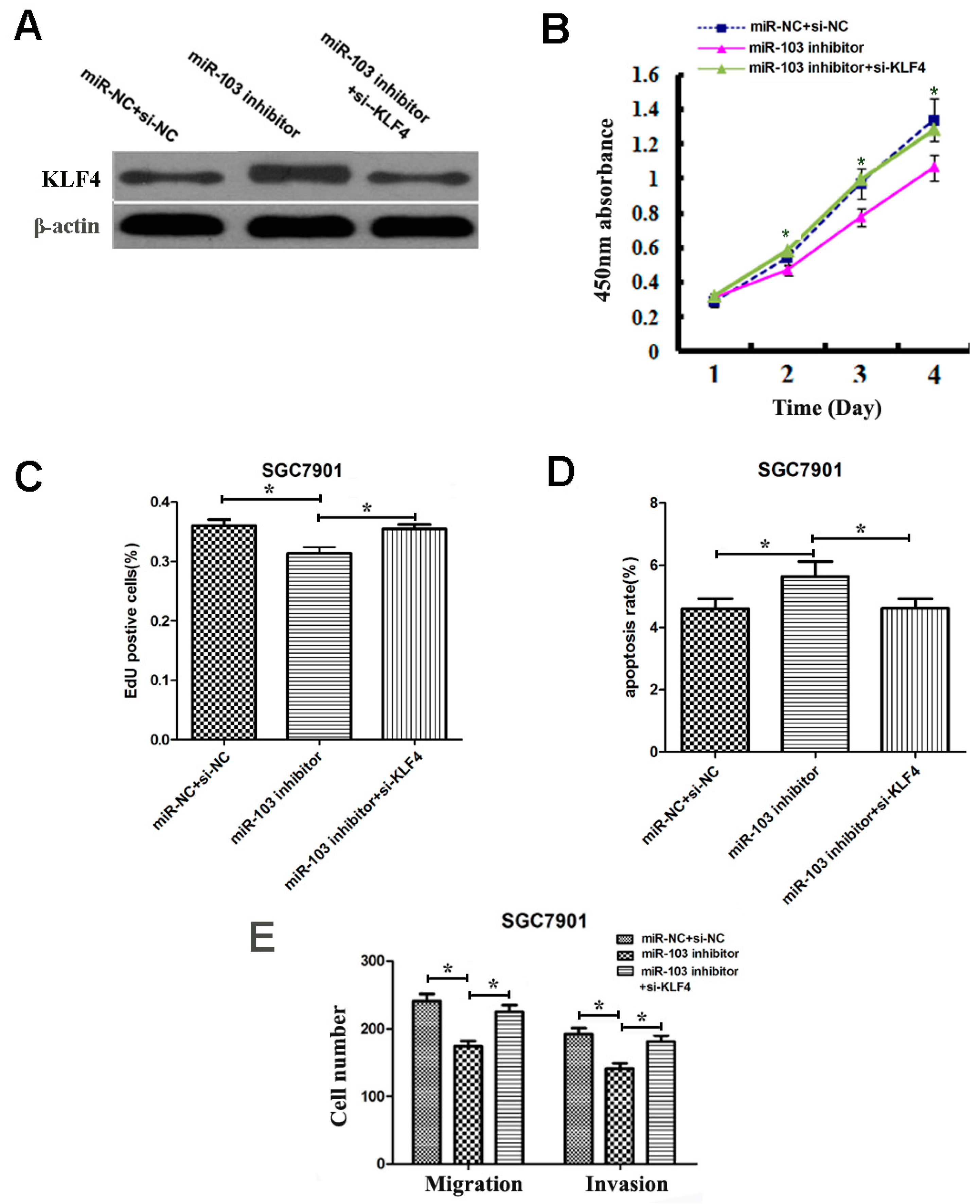
2.9. Overexpression of KLF4 Could Rescue the Oncogenic Effects of miR-103 in GC
3. Discussion
4. Materials and Methods
4.1. Tissue Specimens
4.2. Cell Culture and Transfection
4.3. RNA Extraction and Real-Time PCR (qRT-PCR)
4.4. Cell Proliferation Assay
4.5. Flow Cytometry Apoptosis Assay
4.6. Cell Migration and Invasion Assay
4.7. Vector Construction and Luciferase Reporter Assay
4.8. Western Blot Analysis
4.9. Xenograft Model Experiment
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Sun, B.; Gao, B.; Wang, Z.; Quan, C.; Wei, F.; Fang, X.D. MicroRNA-103 promotes colorectal cancer by targeting tumor suppressor DICER and PTEN. Int. J. Mol. Sci. 2014, 15, 8458–8472. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhou, H.; Xun, Q.; Xu, X.; Ling, J.; Hu, Y. MicroRNA-103 regulates the growth and invasion of endometrial cancer cells through the downregulation of tissue inhibitor of metalloproteinase 3. Oncol. Lett. 2012, 3, 1221–1226. [Google Scholar] [PubMed]
- Fu, X.; Zhang, W.; Su, Y.; Lu, L.; Wang, D.; Wang, H. MicroRNA-103 suppresses tumor cell proliferation by targeting PDCD10 in prostate cancer. Prostate 2016, 76, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Volinia, S.; Okumura, H.; Shimizu, M.; Taccioli, C.; Rossi, S.; Alder, H.; Liu, C.G.; Oue, N.; Yasui, W.; et al. Relation between microRNA expression and progression and prognosis of gastric cancer: A microRNA expression analysis. Lancet Oncol. 2010, 11, 136–146. [Google Scholar] [CrossRef]
- Tchernitsa, O.; Kasajima, A.; Schafer, R.; Kuban, R.J.; Ungethum, U.; Gyorffy, B.; Neumann, U.; Simon, E.; Weichert, W.; Ebert, M.P.; et al. Systematic evaluation of the miRNA-ome and its downstream effects on mRNA expression identifies gastric cancer progression. J. Pathol. 2010, 222, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Hong, S.W.; Kim, A.; Choi, S.H.; Yoon, S.O. Prognostic implications for high expression of oncogenic microRNAs in advanced gastric carcinoma. J. Surg. Oncol. 2013, 107, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Rotkrua, P.; Shimada, S.; Mogushi, K.; Akiyama, Y.; Tanaka, H.; Yuasa, Y. Circulating microRNAs as biomarkers for early detection of diffuse-type gastric cancer using a mouse model. Br. J. Cancer 2013, 108, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, P.; Zhou, Z.; Natarelli, L.; Wei, Y.; Nazari-Jahantigh, M.; Zhu, M.; Grommes, J.; Steffens, S.; Weber, C.; Schober, A. Endothelial Dicer promotes atherosclerosis and vascular inflammation by miRNA-103-mediated suppression of KLF4. Nat. Commun. 2016, 7, 10521. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Christy, R.J.; Yang, V.W. Identification and characterization of a gene encoding a gut-enriched Kruppel-like factor expressed during growth arrest. J. Biol. Chem. 1996, 271, 20009–20017. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Sinha, L.A.; Eberspaecher, H.; Seldin, M.F.; de Crombrugghe, B. A gene for a novel zinc-finger protein expressed in differentiated epithelial cells and transiently in certain mesenchymal cells. J. Biol. Chem. 1996, 271, 31384–31390. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Tian, C.; Ding, Y.; Ye, Q.; Gao, Y.; Yang, N.; Li, Q. Down-regulation of Kruppel-like factor-4 by microRNA-135a-5p promotes proliferation and metastasis in hepatocellular carcinoma by transforming growth factor-β1. Oncotargt 2016, 7, 42566–42578. [Google Scholar]
- Hu, W.; Jia, Y.; Xiao, X.; Lv, K.; Chen, Y.; Wang, L.; Luo, X.; Liu, T.; Li, W.; Li, Y.; et al. KLF4 downregulates hTERT expression and telomerase activity to inhibit lung carcinoma growth. Oncotarget 2016, 7, 52870–52887. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Hisamuddin, I.M.; Nandan, M.O.; Babbin, B.A.; Lamb, N.E.; Yang, V.W. Identification of Kruppel-like factor 4 as a potential tumor suppressor gene in colorectal cancer. Oncogene 2004, 23, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.W.; Ren, S.; Louro, I.D.; Lobo-Ruppert, S.M.; Mckie-Bell, P.; Grizzle, W.; Hayes, M.R.; Broker, T.R.; Chow, L.T.; Ruppert, J.M. Oncogene expression cloning by retroviral transduction of adenovirus E1A-immortalized rat kidney RK3E cells: Transformation of a host with epithelial features by c-MYC and the zinc finger protein GKLF. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1999, 10, 423–434. [Google Scholar]
- Wei, D.; Gong, W.; Kanai, M.; Schlunk, C.; Wang, L.; Yao, J.C.; Wu, T.T.; Huang, S.; Xie, K. Drastic down-regulation of Kruppel-like factor 4 expression is critical in human gastric cancer development and progression. Cancer Res. 2005, 65, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, J.; Shuai, L.; Zha, L.; He, M.; Huang, Z.; Wang, Z. Kruppel-like factor 4 negatively regulates β-catenin expression and inhibits the proliferation, invasion and metastasis of gastric cancer. Int. J. Oncol. 2012, 40, 2038–2048. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Number | Median | p Value |
|---|---|---|---|
| Age (years) | 92 | ||
| ≥60 | 52 | 0.6181 | 0.1022 |
| <60 | 40 | 0.3569 | |
| Gender | |||
| Male | 54 | 0.5837 | 0.2351 |
| Female | 38 | 0.3920 | |
| Tumor size (cm) | |||
| <5 | 44 | 0.3377 | 0.0429 * |
| ≥5 | 48 | 0.6575 | |
| Lauren’s classification | |||
| Intestinal | 52 | 0.3247 | 0.0088 * |
| Diffuse | 40 | 0.7383 | |
| Differentiation status | |||
| Well/moderate | 45 | 0.6385 | 0.0979 |
| Poor | 47 | 0.3762 | |
| TNM stage | |||
| I + II | 32 | 0.3635 | 0.1948 |
| III + IV | 60 | 0.5797 | |
| Lymph node metastasis | |||
| Negative | 36 | 0.3098 | 0.048 * |
| Positive | 56 | 0.6297 |
| Human Gene | Primer Sequence 5′ to 3′ | |
|---|---|---|
| Forward | Reverse | |
| E-cadherin | CGAGAGCTACACGTTCACGG | GGGTGTCGAGGGAAAAATAGG |
| Vimentin | GACGCCATCAACACCGAGT | CTTTGTCGTTGGTTAGCTGGT |
| Twist | GTCCGCAGTCTTACGAGGAG | GCTTGAGGGTCTGAATCTTGCT |
| Snail | TCGGAAGCCTAACTACAGCGA | AGATGAGCATTGGCAGCGAG |
| Slug | CAACGCCTCCAAAAAGCCAA | ACTCACTCGCCCCAAAGATG |
| KLF4 | TCGGACCACCTCGCCTTACA | CTGGGCTCCTTCCCTCATCG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Liu, Y.; Qiao, Y.; Zhang, L.; Lu, S. miR-103 Promotes Proliferation and Metastasis by Targeting KLF4 in Gastric Cancer. Int. J. Mol. Sci. 2017, 18, 910. https://doi.org/10.3390/ijms18050910
Zheng J, Liu Y, Qiao Y, Zhang L, Lu S. miR-103 Promotes Proliferation and Metastasis by Targeting KLF4 in Gastric Cancer. International Journal of Molecular Sciences. 2017; 18(5):910. https://doi.org/10.3390/ijms18050910
Chicago/Turabian StyleZheng, Jie, Yuzhen Liu, Yanchun Qiao, Liying Zhang, and Shijun Lu. 2017. "miR-103 Promotes Proliferation and Metastasis by Targeting KLF4 in Gastric Cancer" International Journal of Molecular Sciences 18, no. 5: 910. https://doi.org/10.3390/ijms18050910
APA StyleZheng, J., Liu, Y., Qiao, Y., Zhang, L., & Lu, S. (2017). miR-103 Promotes Proliferation and Metastasis by Targeting KLF4 in Gastric Cancer. International Journal of Molecular Sciences, 18(5), 910. https://doi.org/10.3390/ijms18050910