
Zampanolide, a Microtubule-Stabilizing Agent, Is Active in Resistant Cancer Cells and Inhibits Cell Migration

 , and
, and
Abstract
:
1. Introduction
2. Results
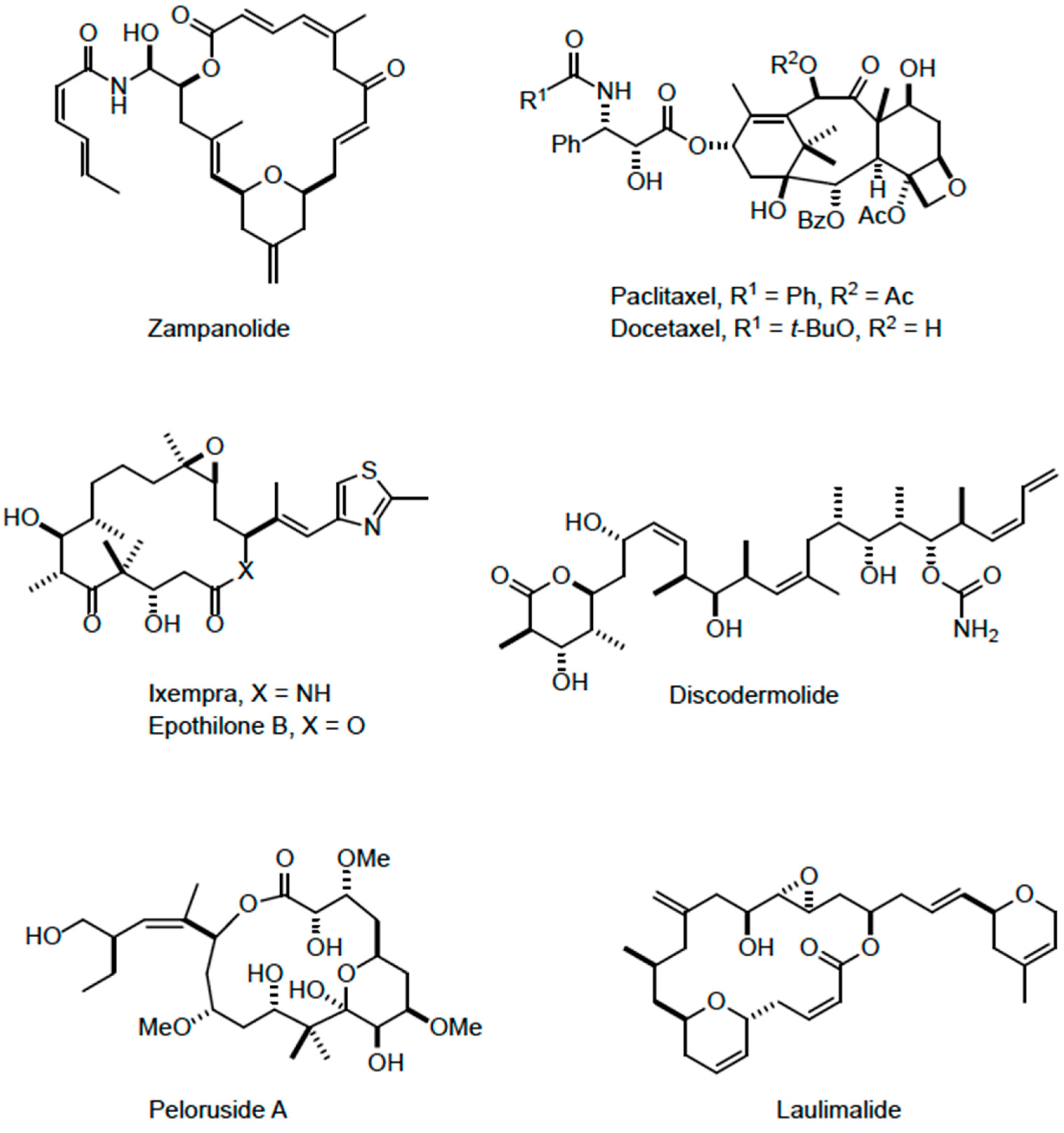
2.1. Growth Inhibition by Zampanolide and Other Microtubule-Stabilizing Agents in Different Cell Lines
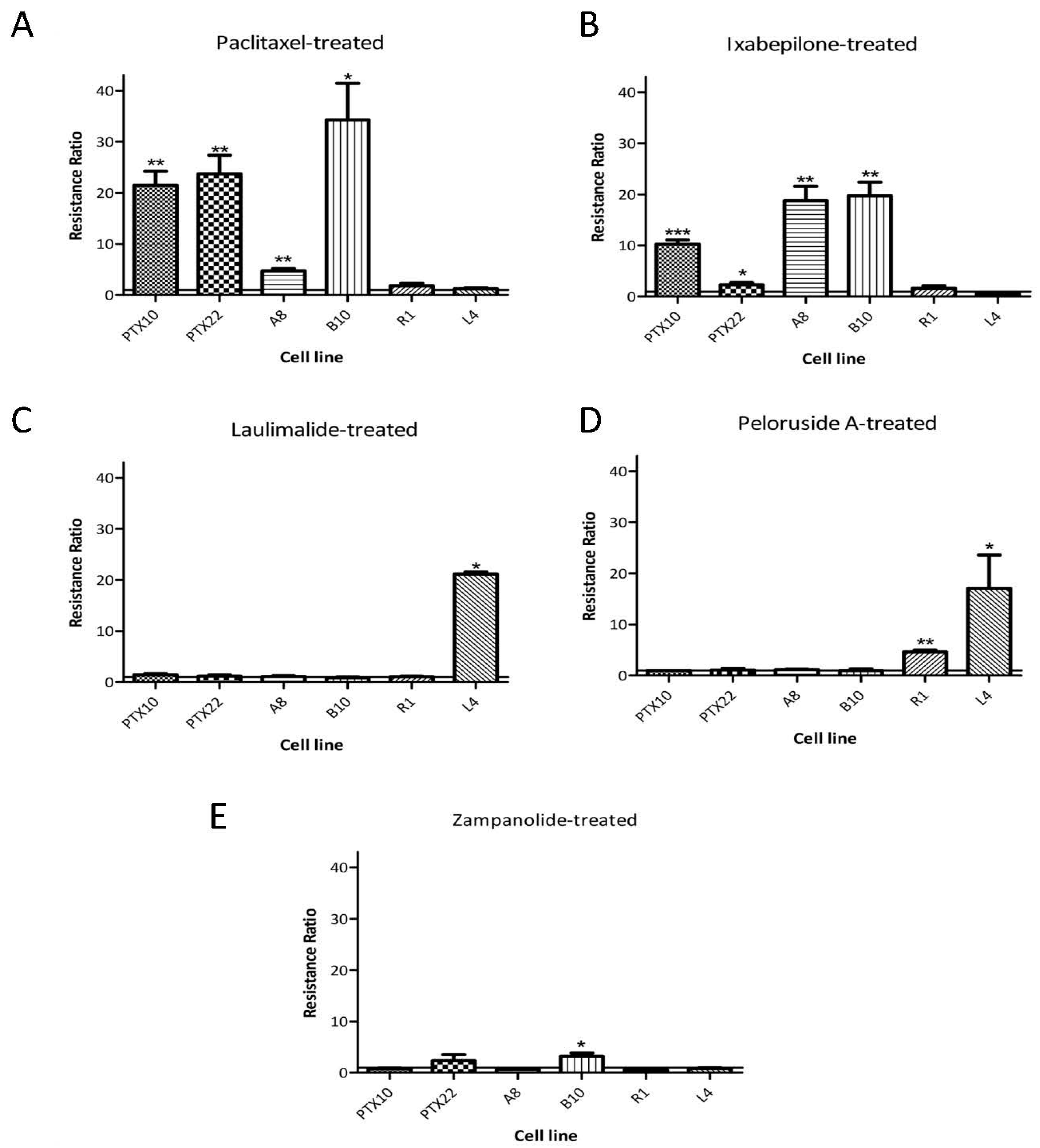
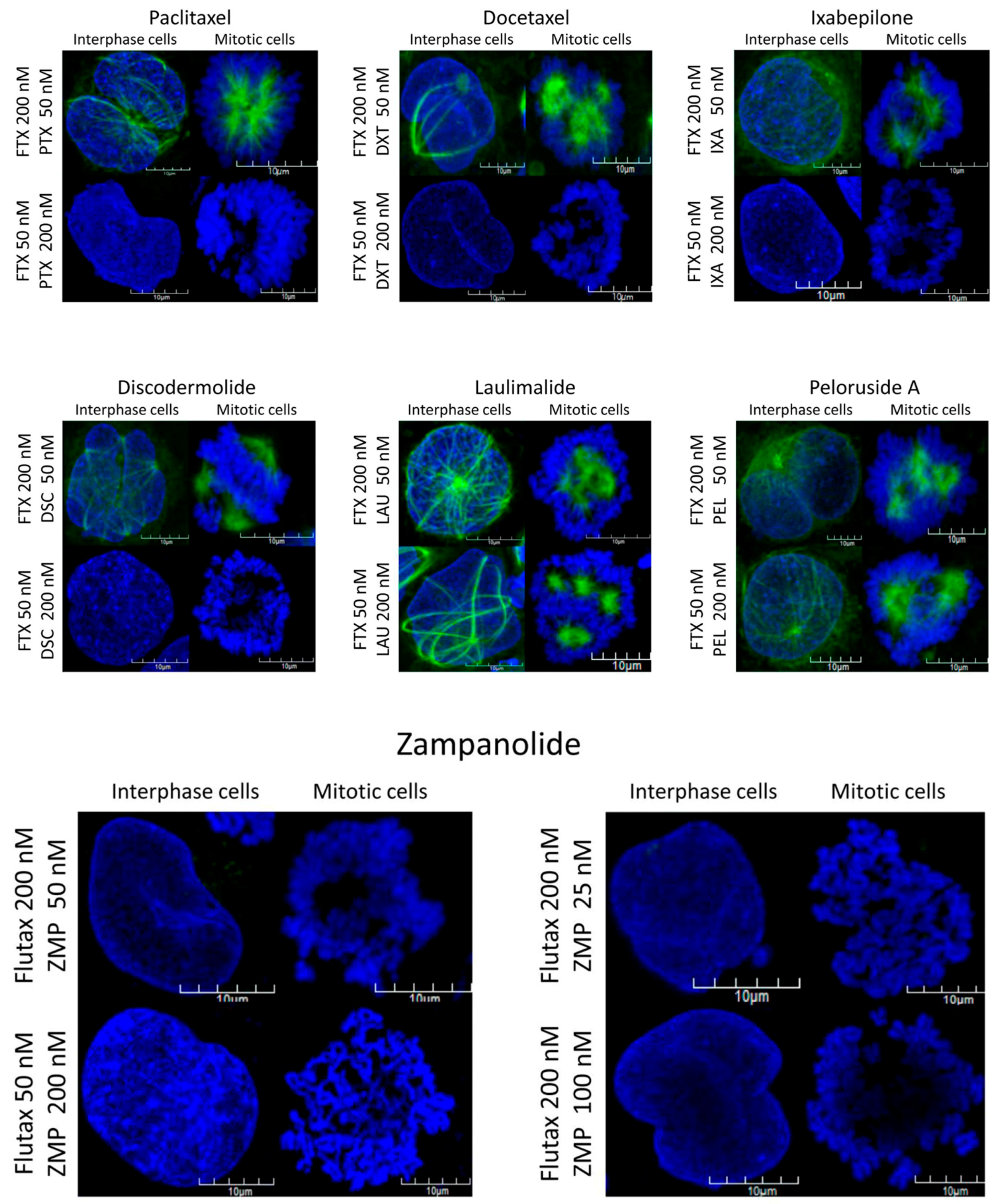
2.2. Action of Zampanolide on Cells with β-Tubulin Mutations
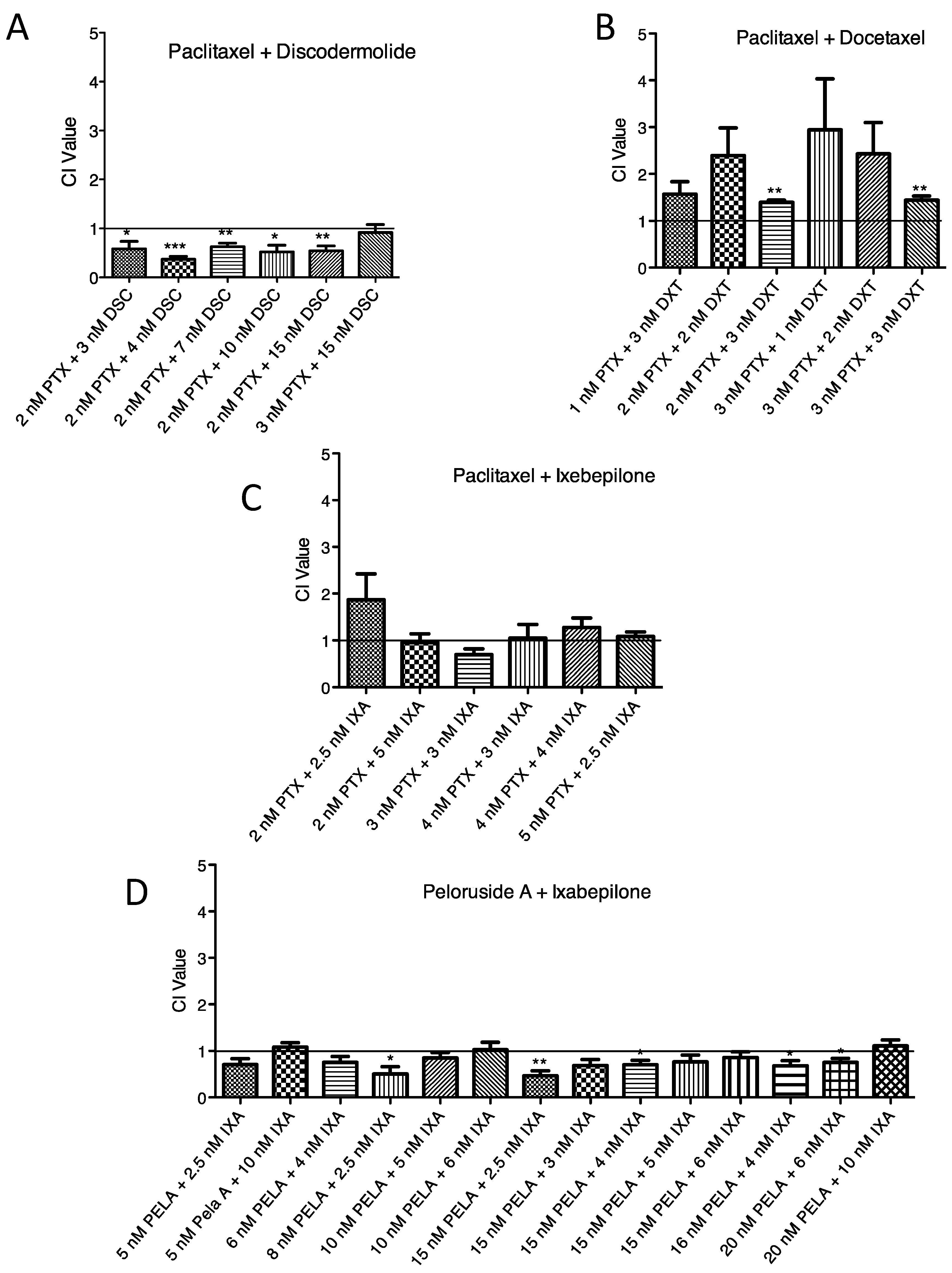
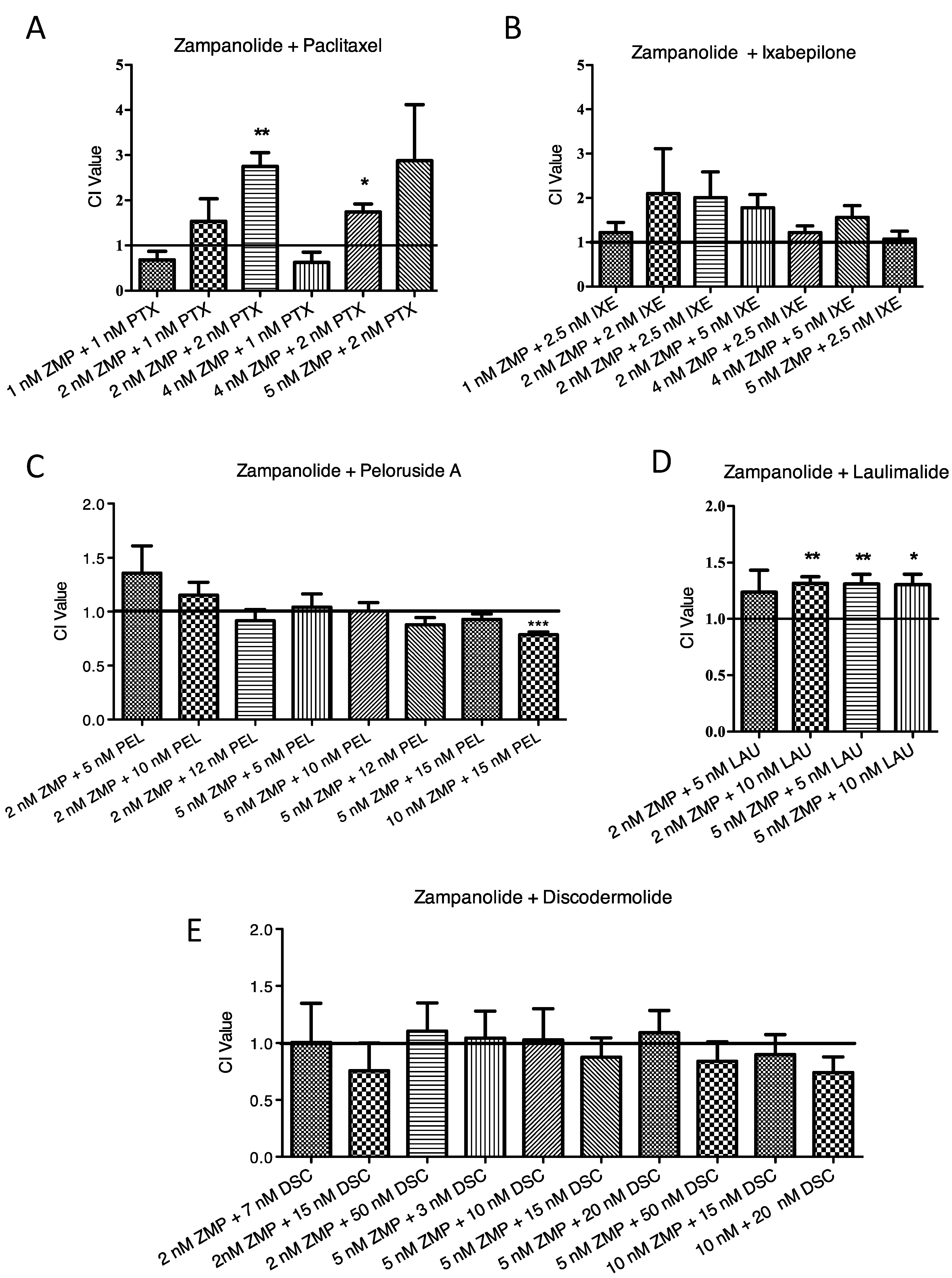
2.3. Lack of Synergistic Interactions between ZMP and Other MSAs
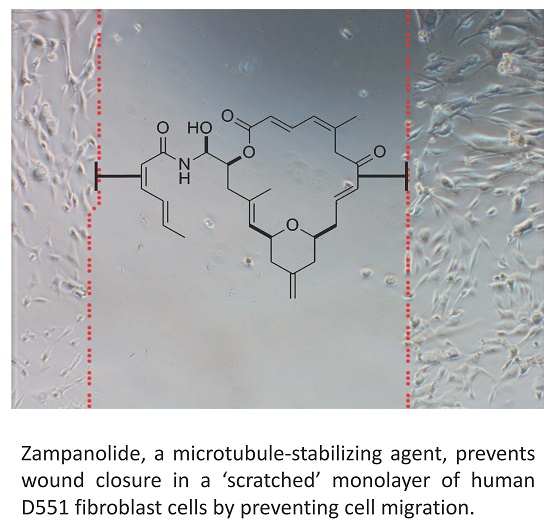
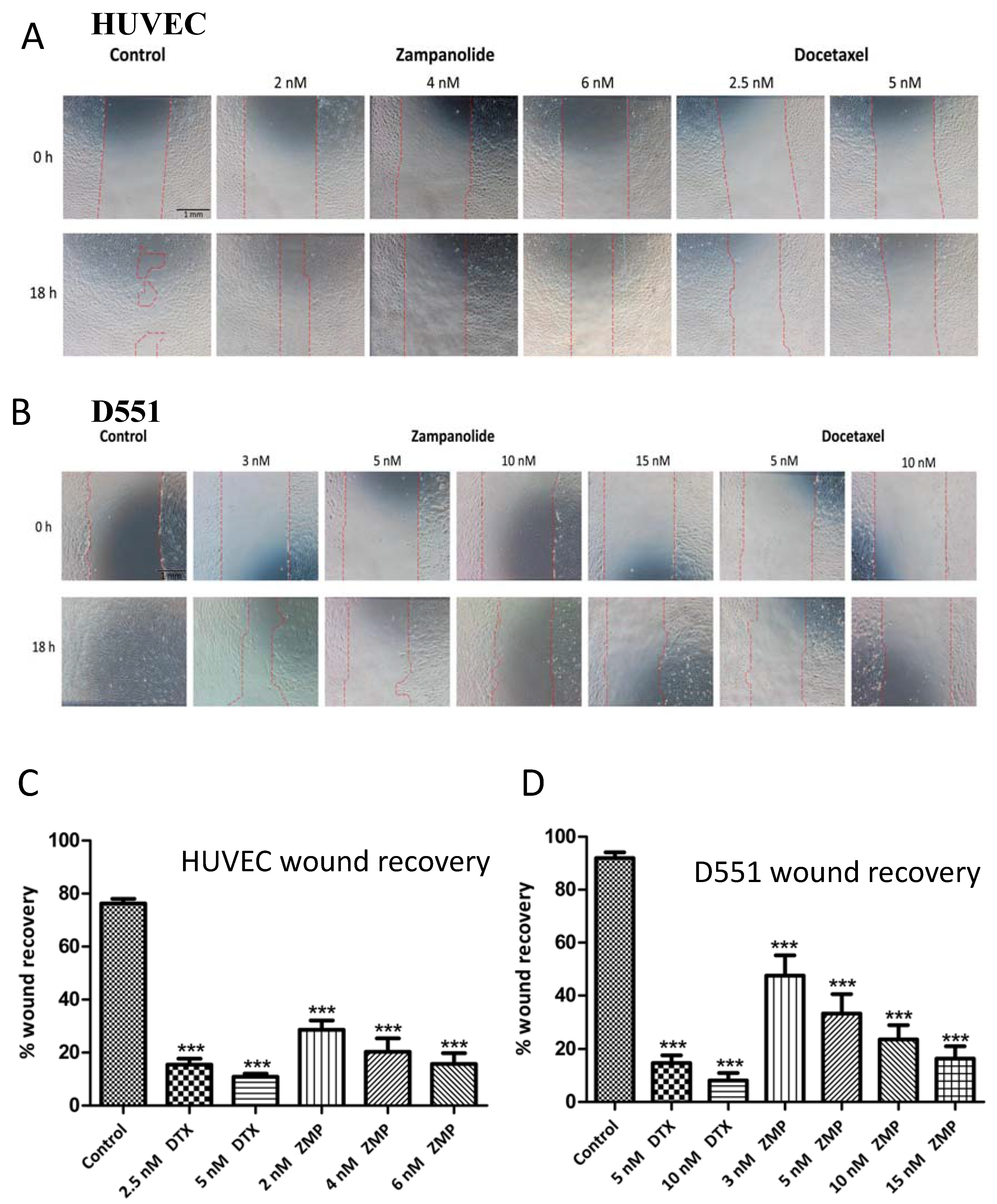
2.4. Zampanolide Effect on Cell Migration in Culture
3. Discussion
3.1. ZMP Cytotoxicity and ZMP Activity in Resistant Cells
3.2. Action of ZMP on Cells with β-Tubulin Mutations
3.3. Lack of Synergy with ZMP
3.4. ZMP Effect on Cell Migration
4. Materials and Methods
4.1. Compounds
4.2. Cell Lines, Cell Culture, and the MTT Cell Proliferation Assay
4.3. The Combination Index (CI Value) for Synergy Determinations in 1A9 Cells
4.4. Wound Scratch Healing Assay with Human Umbilical Vein Endothelial Cells (HUVEC) and D551 Cells
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CI | Combination index |
| MT | Microtubule |
| MTA | Microtubule-targeting agent |
| MSA | Microtubule-stabilizing agent |
| PTX | Paclitaxel |
| P-gp | p-glycoprotein |
| ZMP | Zampanolide |
References
- Field, J.J.; Singh, A.J.; Kanakkanthara, A.; Halafihi, T.; Northcote, P.T.; Miller, J.H. Microtubule-stabilizing activity of zampanolide, a potent macrolide isolated from the Tongan marine sponge Cacospongia mycofijiensis. J. Med. Chem. 2009, 52, 7328–7332. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Pera, B.; Calvo, E.; Canales, A.; Zurwerra, D.; Trigili, C.; Rodríguez-Salarichs, J.; Matesanz, R.; Kanakkanthara, A.; Wakefield, S.J.; et al. Zampanolide, a potent new microtubule-stabilizing agent, covalently reacts with the taxane luminal site in tubulin α,β-heterodimers and microtubules. Chem. Biol. 2012, 19, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Buey, R.M.; Calvo, E.; Barasoain, I.; Pineda, O.; Edler, M.C.; Matesanz, R.; Cerezo, G.; Vanderwal, C.D.; Day, B.W.; Sorensen, E.J.; et al. Cyclostreptin binds covalently to microtubule pores and lumenal taxoid binding sites. Nat. Chem. Biol. 2007, 3, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Risinger, A.L.; Li, J.; Bennett, M.J.; Rohena, C.C.; Schriemer, D.C.; Mooberry, S.L. Taccalonolide binding to tubulin imparts microtubule stability and potent in vivo activity. Cancer Res. 2013, 73, 6780–6792. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Petter, R.C.; Baillie, T.A.; Whitty, A. The resurgence of covalent drugs. Nat. Rev. Drug Discov. 2011, 10, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Cabral, F.; Abraham, I.; Gottesman, M.M. Isolation of a taxol-resistant Chinese hamster ovary cell mutant that has an alteration in α-tubulin. Proc. Natl. Acad. Sci. USA 1981, 78, 4388–4391. [Google Scholar] [CrossRef] [PubMed]
- Schibler, M.J.; Cabral, F. Taxol-dependent mutants of Chinese hamster ovary cells with alterations in α- and β-tubulin. J. Cell Biol. 1986, 102, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Bhattacharya, R.; Cabral, F. Human mutations that confer paclitaxel resistance. Mol. Cancer Ther. 2010, 9, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Giannakakou, P.; Sackett, D.L.; Kang, Y.K.; Zhan, Z.; Buters, J.T.; Fojo, T.; Poruchynsky, M.S. Paclitaxel-resistant human ovarian cancer cells have mutant β-tubulins that exhibit impaired paclitaxel-driven polymerization. J. Biol. Chem. 1997, 272, 17118–17125. [Google Scholar] [CrossRef] [PubMed]
- Giannakakou, P.; Gussio, R.; Nogales, E.; Downing, K.H.; Zaharevitz, D.; Bollbuck, B.; Poy, G.; Sackett, D.; Nicolaou, K.C.; Fojo, T. A common pharmacophore for epothilone and taxanes: Molecular basis for drug resistance conferred by tubulin mutations in human cancer cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2904–2909. [Google Scholar] [CrossRef] [PubMed]
- Kanakkanthara, A.; Wilmes, A.; O'Brate, A.; Escuin, D.; Chan, A.; Gjyrezi, A.; Crawford, J.; Rawson, P.; Kivell, B.; Northcote, P.T.; et al. Peloruside- and laulimalide-resistant human ovarian carcinoma cells have βI-tubulin mutations and altered expression of βII- and βIII-tubulin isotypes. Mol. Cancer Ther. 2011, 10, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Begaye, A.; Trostel, S.; Zhao, Z.; Taylor, R.E.; Schriemer, D.C.; Sackett, D.L. Mutations in the β-tubulin binding site for peloruside A confer resistance by targeting a cleft significant in side chain binding. Cell Cycle 2011, 10, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.H.; Zeng, C.Q.; Hari, M.; Cabral, F. Random mutagenesis of β-tubulin defines a set of dispersed mutations that confer paclitaxel resistance. Pharm. Res. 2012, 29, 2994–3006. [Google Scholar] [CrossRef] [PubMed]
- Basciano, P.A.; Matakas, J.; Pecci, A.; Civaschi, E.; Cagioni, C.; Bompiani, N.; Burger, P.; Christos, P.; Snyder, J.P.; Bussel, J.; et al. β-1 tubulin R307H SNP alters microtubule dynamics and affects severity of a hereditary thrombocytopenia. J. Thromb. Haemost. 2015, 13, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Zurwerra, D.; Field, J.J.; Díaz, J.F.; Altmann, K.-H.; Steinmetz, M.O. Molecular mechanism of action of microtubule-stabilizing anticancer agents. Science 2013, 339, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Northcote, P.T.; Marsh, M.; Altmann, K.-H.; Miller, J.H.; Díaz, J.F.; Steinmetz, M.O. Structural basis of microtubule stabilization by laulimalide and peloruside A. Angew. Chem. Int. Ed. Engl. 2014, 53, 1621–1625. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.A.; Hills, P.M.; Davidson, B.S.; Wender, P.A.; Mooberry, S.L. Laulimalide and synthetic laulimalide analogues are synergistic with paclitaxel and 2-methoxyestradiol. Mol. Pharm. 2006, 3, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Wilmes, A.; Bargh, K.; Kelly, C.; Northcote, P.T.; Miller, J.H. Peloruside A synergizes with other microtubule stabilizing agents in cultured cancer cell lines. Mol. Pharm. 2007, 4, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Wilmes, A.; O’Sullivan, D.; Chan, A.; Chandrahasen, C.; Paterson, I.; Northcote, P.T.; La Flamme, A.C.; Miller, J.H. Synergistic interactions between peloruside A and other microtubule-stabilizing and destabilizing agents in cultured human ovarian carcinoma cells and murine T cells. Cancer Chemother. Pharmacol. 2011, 68, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Giannakakou, P.; Fojo, T. Discodermolide: Just another microtubule-stabilizing agent? No! A lesson in synergy. Clin. Cancer Res. 2000, 6, 1613–1615. [Google Scholar] [PubMed]
- Martello, L.A.; McDaid, H.M.; Regl, D.L.; Yang, C.P.; Meng, D.; Pettus, T.R.; Kaufman, M.D.; Arimoto, H.; Danishefsky, S.J.; Smith, A.B., 3rd; et al. Taxol and discodermolide represent a synergistic drug combination in human carcinoma cell lines. Clin. Cancer Res. 2000, 6, 1978–1987. [Google Scholar] [PubMed]
- Honore, S.; Kamath, K.; Braguer, D.; Horwitz, S.B.; Wilson, L.; Briand, C.; Jordan, M.A. Synergistic suppression of microtubule dynamics by discodermolide and paclitaxel in non-small cell lung carcinoma cells. Cancer Res. 2004, 64, 4957–4964. [Google Scholar] [CrossRef] [PubMed]
- Khrapunovich-Baine, M.; Menon, V.; Verdier-Pinard, P.; Smith, A.B., 3rd; Angeletti, R.H.; Fiser, A.; Horwitz, S.B.; Xiao, H. Distinct pose of discodermolide in taxol binding pocket drives a complementary mode of microtubule stabilization. Biochemistry 2009, 48, 11664–11677. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Bargsten, K.; Redondo, M.; Smith, A.B., 3rd; Yang, C.H.; McDaid, H.M.; Paterson, I.; Horwitz, S.B.; Díaz, J.F.; Steinmetz, M.O. Structural basis of microtubule stabilization by discodermolide. ChemBioChem 2017. [Google Scholar] [CrossRef]
- Brooks, S.A.; Lomax-Browne, H.J.; Carter, T.M.; Kinch, C.E.; Hall, D.M.S. Molecular interactions in cancer cell metastasis. Acta Histochem. 2010, 112, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Tozer, G.M.; Kanthou, C.; Baguley, B.C. Disrupting tumour blood vessels. Nat. Rev. Cancer 2005, 5, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Falasca, M.; Raimondi, C.; Maffucci, T. Boyden chamber. Methods Mol. Biol. 2011, 769, 87–95. [Google Scholar] [PubMed]
- Hall, D.M.; Brooks, S.A. In vitro invasion assay using Matrigel™: A reconstituted basement membrane preparation. Methods Mol. Biol. 2014, 1070, 1–11. [Google Scholar] [PubMed]
- Belotti, D.; Vergani, V.; Drudis, T.; Borsotti, P.; Pitelli, M.R.; Viale, G.; Giavazzi, R.; Taraboletti, G. The microtubule-affecting drug paclitaxel has antiangiogenic activity. Clin. Cancer Res. 1996, 2, 1843–1849. [Google Scholar] [PubMed]
- Lu, H.; Murtagh, J.; Schwartz, E.L. The microtubule binding drug laulimalide inhibits vascular endothelial growth factor-induced human endothelial cell migration and is synergistic when combined with docetaxel (Taxotere). Mol. Pharmacol. 2006, 69, 1207–1215. [Google Scholar] [CrossRef]
- Yang, H.; Ganguly, A.; Cabral, F. Inhibition of cell migration and cell division correlates with distinct effects of microtubule inhibiting drugs. J. Biol. Chem. 2010, 285, 32242–32250. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Singh, A.J.; Northcote, P.T.; Miller, J.H. Inhibition of human vascular endothelial cell migration and capillary-like tube formation by the microtubule-stabilizing agent peloruside A. Investig. New Drugs 2015, 33, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Kamath, K.; Smiyun, G.; Wilson, L.; Jordan, M.A. Mechanisms of inhibition of endothelial cell migration by taxanes. Cytoskeleton 2014, 71, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Kaverina, I.; Straube, A. Regulation of cell migration by dynamic microtubules. Semin. Cell. Dev. Biol. 2011, 22, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Yang, H.; Sharma, R.; Patel, K.D.; Cabral, F. The role of microtubules and their dynamics in cell migration. J. Biol. Chem. 2012, 287, 43359–43369. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Yang, H.; Zhang, H.; Cabral, F.; Patel, K.D. Microtubule dynamics control tail retraction in migrating vascular endothelial cells. Mol. Cancer Ther. 2013, 12, 2837–2846. [Google Scholar] [CrossRef] [PubMed]
- Zurwerra, D.; Glaus, F.; Betschart, L.; Schuster, J.; Gertsch, J.; Ganci, W.; Altmann, K.-H. Total synthesis of (−)-zampanolide and structure-activity relationship studies on (−)-dactylolide derivatives. Chemistry 2012, 18, 16868–16883. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Calvo, E.; Northcote, P.T.; Miller, J.H.; Altmann, K.-H.; Díaz, J.F. Methods for studying microtubule binding site interactions: Zampanolide as a covalent binding agent. In Methods in Cell Biology, Microtubules in Vitro, 2nd ed.; Wilson, L., Correia, J.J., Eds.; Academic Press: Burlington, NJ, USA, 2013; Volume 115, pp. 303–325. [Google Scholar]
- Gaitanos, T.N.; Buey, R.M.; Díaz, J.F.; Northcote, P.T.; Teesdale-Spittle, P.; Andreu, J.M.; Miller, J.H. Peloruside A does not bind to the taxoid site on β-tubulin and retains its activity in multidrug-resistant cell lines. Cancer Res. 2004, 64, 5063–5067. [Google Scholar] [CrossRef] [PubMed]
- Dumontet, C.; Jordan, M.A. Microtubule-binding agents: A dynamic field of cancer therapeutics. Nat. Rev. Drug Discov. 2010, 9, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Risinger, A.L.; Jackson, E.M.; Polin, L.A.; Helms, G.L.; LeBoeuf, D.A.; Joe, P.A.; Hopper-Borge, E.; Ludueña, R.F.; Kruh, G.D.; Mooberry, S.L. The taccalonolides: Microtubule stabilizers that circumvent clinically relevant taxane resistance mechanisms. Cancer Res. 2008, 68, 8881–8888. [Google Scholar] [CrossRef] [PubMed]
- Kanakkanthara, A.; Teesdale-Spittle, P.H.; Miller, J.H. Cytoskeletal alterations that confer resistance to anti-tubulin chemotherapeutics. Anticancer Agents Med. Chem. 2013, 13, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Díaz, J.F.; Miller, J.H. The binding sites of microtubule-stabilizing agents. Chem. Biol. 2013, 20, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Kanakkanthara, A.; Eras, J.; Northcote, P.T.; Cabral, F.; Miller, J.H. Resistance to peloruside A and laulimalide: Functional significance of acquired βI-tubulin mutations at sites important for drug-tubulin binding. Curr. Cancer Drug Targets. 2014, 14, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Gapud, E.J.; Bai, R.; Ghosh, A.K.; Hamel, E. Laulimalide and paclitaxel: A comparison of their effects on tubulin assembly and their synergistic action when present simultaneously. Mol. Pharmacol. 2004, 66, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Hamel, E.; Day, B.W.; Miller, J.H.; Jung, M.K.; Northcote, P.T.; Ghosh, A.K.; Curran, D.P.; Cushman, M.; Nicolaou, K.C.; Paterson, I.; et al. Synergistic effects of peloruside A and laulimalide with taxoid site drugs, but not with each other, on tubulin assembly. Mol. Pharmacol. 2006, 70, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Photiou, A.; Shah, P.; Leong, L.K.; Moss, J.; Retsas, S. In vitro synergy of paclitaxel (Taxol) and vinorelbine (navelbine) against human melanoma cell lines. Eur. J. Cancer 1997, 33, 463–470. [Google Scholar] [CrossRef]
- Giannakakou, P.; Villalba, L.; Li, H.; Poruchynsky, M.; Fojo, T. Combinations of paclitaxel and vinblastine and their effects on tubulin polymerization and cellular cytotoxicity: Characterization of a synergistic schedule. Int. J. Cancer 1998, 75, 57–63. [Google Scholar] [CrossRef]
- Ricker, J.L.; Chen, Z.; Yang, X.P.; Pribluda, V.S.; Swartz, G.M.; van Waes, C. 2-methoxyestradiol inhibits hypoxia-inducible factor 1α, tumor growth, and angiogenesis and augments paclitaxel efficacy in head and neck squamous cell carcinoma. Clin. Cancer Res. 2004, 10, 8665–8673. [Google Scholar] [CrossRef] [PubMed]
- Han, G.Z.; Liu, Z.J.; Shimoi, K.; Zhu, B.T. Synergism between the anticancer actions of 2-methoxyestradiol and microtubule-disrupting agents in human breast cancer. Cancer Res. 2005, 65, 387–393. [Google Scholar] [PubMed]
- Huang, G.S.; Lopez-Barcons, L.; Freeze, B.S.; Smith, A.B., 3rd; Goldberg, G.L.; Horwitz, S.B.; McDaid, H.M. Potentiation of taxol efficacy by discodermolide in ovarian carcinoma xenograft-bearing mice. Clin. Cancer Res. 2006, 12, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Khrapunovich-Baine, M.; Menon, V.; Yang, C.P.; Northcote, P.T.; Miller, J.H.; Angeletti, R.H.; Fiser, A.; Horwitz, S.B.; Xiao, H. Hallmarks of molecular action of microtubule stabilizing agents: Effects of epothilone B, ixabepilone, peloruside A, and laulimalide on microtubule conformation. J. Biol. Chem. 2011, 286, 11765–11778. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Cabral, F.; Yang, H.; Patel, K. Peloruside A is a microtubule-stabilizing agent with exceptional anti-migratory properties in human endothelial cells. Oncoscience 2015, 2, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Paterson, I.; Florence, G.J.; Gerlach, K.; Scott, J.P. Total synthesis of the antimicrotubule agent (+)-discodermolide using boron-mediated aldol reactions of chiral ketones. Angew. Chem. Int. Ed. Engl. 2000, 39, 377–380. [Google Scholar] [CrossRef]
- West, L.M.; Northcote, P.T.; Battershill, C.N. Peloruside A: A potent cytotoxic macrolide isolated from the New Zealand marine sponge Mycale sp. J. Org. Chem. 2000, 65, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Hood, K.A.; West, L.M.; Northcote, P.T.; Berridge, M.V.; Miller, J.H. Induction of apoptosis by the marine sponge (Mycale) metabolites, mycalamide A and pateamine. Apoptosis 2001, 6, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Berenbaum, M.C. The expected effect of a combination of agents: The general solution. J. Theor. Biol. 1985, 114, 413–431. [Google Scholar] [CrossRef]
- Tanaka, J.; Higa, T. Zampanolide, a new cytotoxic macrolide from a marine sponge. Tetrahedron Lett. 1996, 37, 5535–5538. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 ± SEM (nM) |
|---|---|
| Taxoid site ligands | |
| Zampanolide | 9.54 ± 0.85 |
| Paclitaxel | 3.71 ± 0.30 |
| Docetaxel | 3.55 ± 0.43 |
| Ixabepilone | 6.65 ± 0.33 |
| Discodermolide | 138 ± 12 |
| Laulimalide/Peloruside site ligands | |
| Peloruside A | 23.3 ± 1.1 |
| Laulimalide | 9.71 ± 0.28 |
| Cell Line | Source of ZMP | IC50 ± SEM (nM) | Duration (h) |
|---|---|---|---|
| 1A9 | natural | 8.2 ± 0.1 | 72 |
| 1A9 | synthetic | 4.6 ± 1.3 | 72 |
| 1A9 [1] | natural | 14.3 ± 2.4 | 72 |
| HL-60 [1] | natural | 4.3 ± 1.1 | 48 |
| D551 | synthetic | 7.3 ± 1.2 | 72 |
| HUVEC | synthetic | 0.6 ± 0.1 | 72 |
| HUVEC | synthetic | 1.0 ± 0.4 | 120 |
| Cell Line | Resistance to | Paclitaxel | Ixabepilone | Zampanolide | Peloruside A | Laulimalide |
|---|---|---|---|---|---|---|
| 1A9 | 4.2 ± 0.3 | 7.3 ± 0.6 | 8.2 ± 1.0 | 20.1 ± 0.9 | 8.3 ± 0.5 | |
| PTX10 | PTX and EPO | 91.7 ±8.2 | 54.9 ± 9.6 | 2.3 ± 0.9 | 17.5 ± 1.2 | 11.0 ± 1.0 |
| PTX22 | PTX | 100 ± 14.1 | 11.4 ± 1.7 | 9.2 ± 3.9 | 19.4 ± 5.4 | 10.6 ± 2.7 |
| A8 | EPO and PTX | 94.4 ± 5.6 | 99.8 ± 0.6 | 14.9 ± 4.6 | 14.0 ± 2.4 | 7.2 ± 1.1 |
| B10 | EPO | 17.2 ± 4.3 | 106 ± 6.5 | 8.6 ± 3.2 | 24.9 ± 1.9 | 10.8 ± 1.0 |
| 1A9-R1 | PLA | 8.8 ± 2.5 | 14.7 ± 3.3 | 5.9 ± 1.6 | 90.9 ± 8.5 | 9.8 ± 1.5 |
| 1A9-L4 | LAU and PLA | 4.2 ± 0.1 | 4.4 ± 0.4 | 4.7 ± 1.0 | 351 ± 126 | 344 ± 150 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Field, J.J.; Northcote, P.T.; Paterson, I.; Altmann, K.-H.; Díaz, J.F.; Miller, J.H. Zampanolide, a Microtubule-Stabilizing Agent, Is Active in Resistant Cancer Cells and Inhibits Cell Migration. Int. J. Mol. Sci. 2017, 18, 971. https://doi.org/10.3390/ijms18050971
Field JJ, Northcote PT, Paterson I, Altmann K-H, Díaz JF, Miller JH. Zampanolide, a Microtubule-Stabilizing Agent, Is Active in Resistant Cancer Cells and Inhibits Cell Migration. International Journal of Molecular Sciences. 2017; 18(5):971. https://doi.org/10.3390/ijms18050971
Chicago/Turabian StyleField, Jessica J., Peter T. Northcote, Ian Paterson, Karl-Heinz Altmann, J. Fernando Díaz, and John H. Miller. 2017. "Zampanolide, a Microtubule-Stabilizing Agent, Is Active in Resistant Cancer Cells and Inhibits Cell Migration" International Journal of Molecular Sciences 18, no. 5: 971. https://doi.org/10.3390/ijms18050971
APA StyleField, J. J., Northcote, P. T., Paterson, I., Altmann, K. -H., Díaz, J. F., & Miller, J. H. (2017). Zampanolide, a Microtubule-Stabilizing Agent, Is Active in Resistant Cancer Cells and Inhibits Cell Migration. International Journal of Molecular Sciences, 18(5), 971. https://doi.org/10.3390/ijms18050971