Leaf Physiological and Proteomic Analysis to Elucidate Silicon Induced Adaptive Response under Salt Stress in Rosa hybrida ‘Rock Fire’





Abstract
:
1. Introduction
2. Results
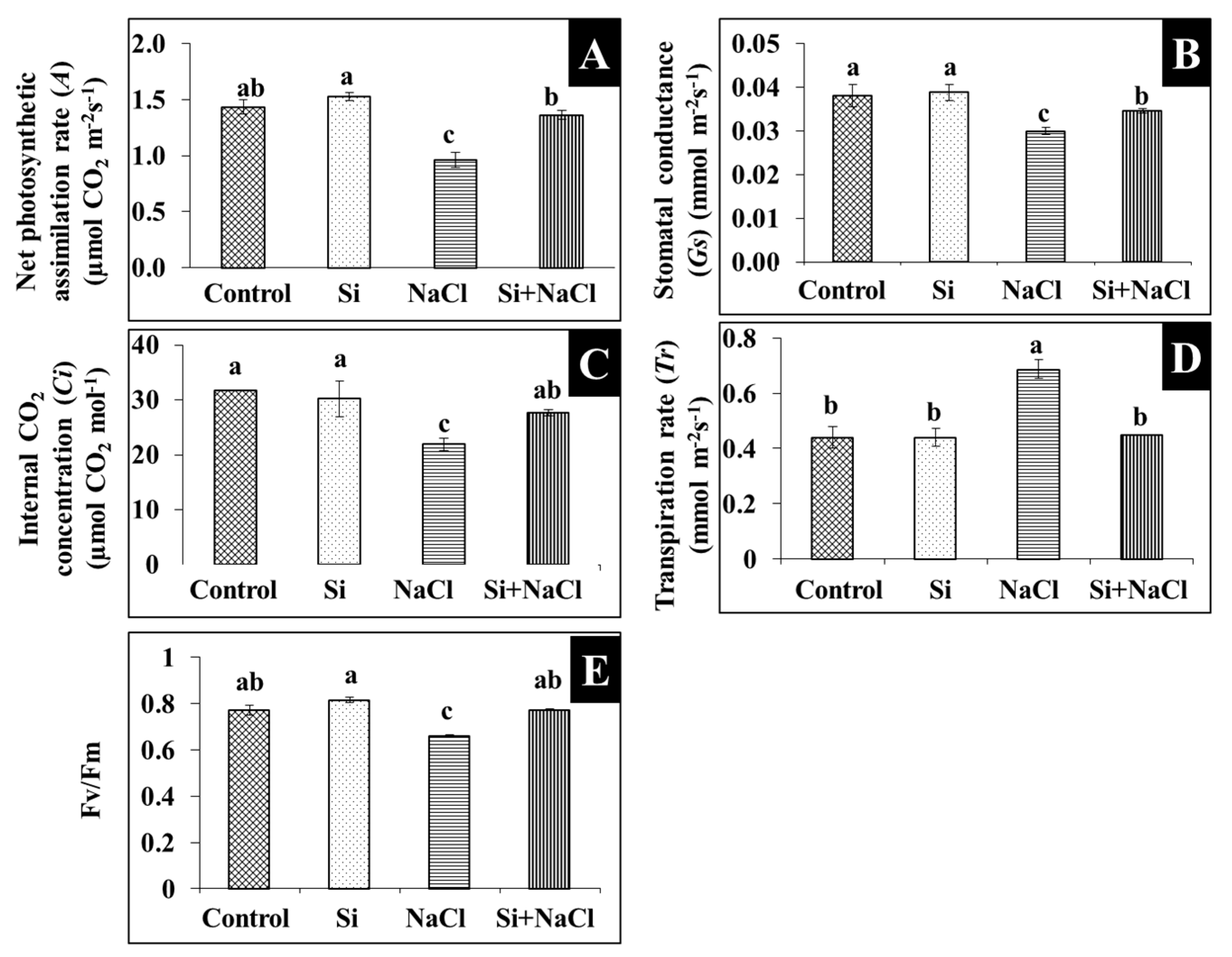
2.1. Physiology and Photosynthesis
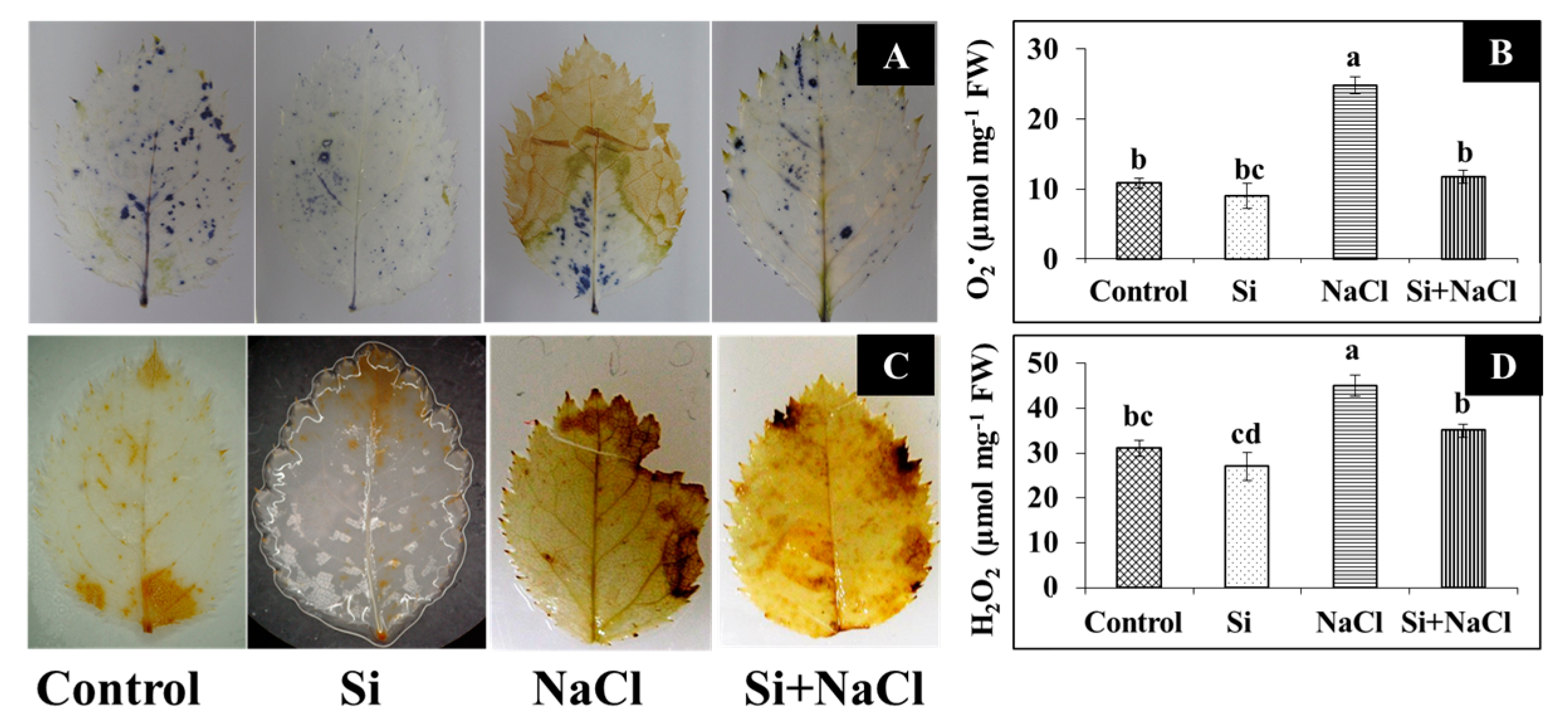
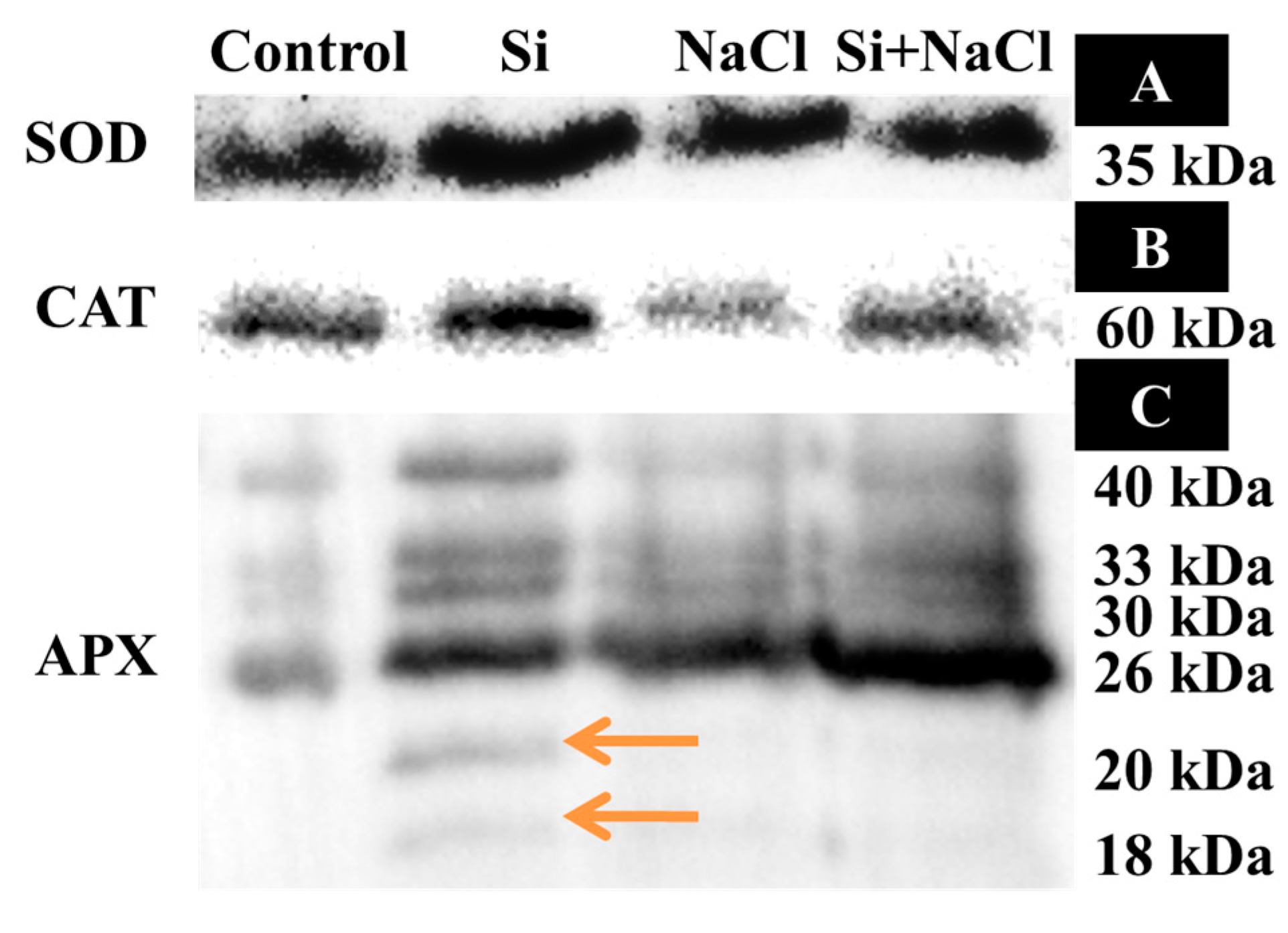
2.2. Oxidative Stress and Analysis of Antioxidant Enzymes
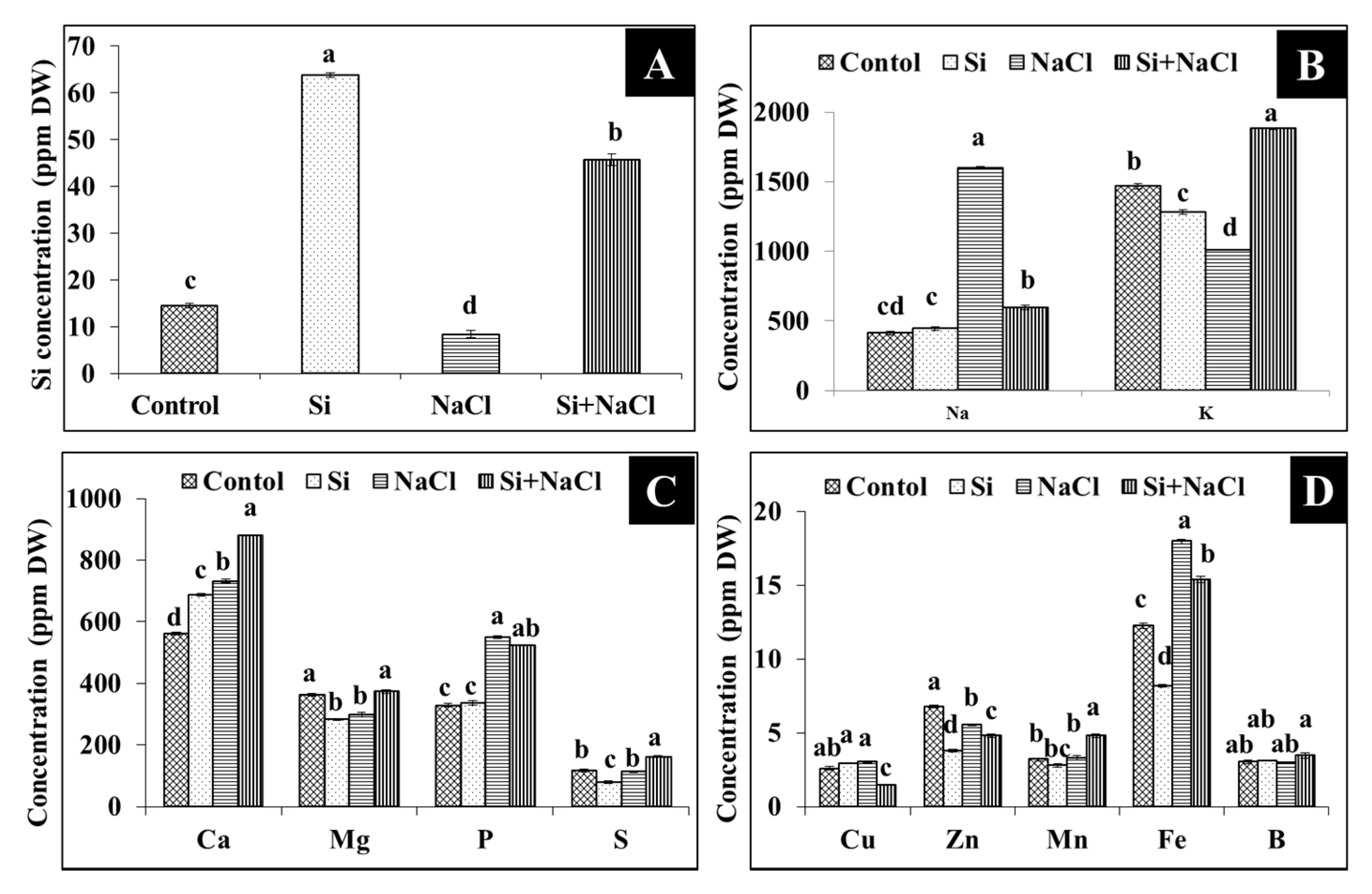
2.3. Uptake of Silicon, Sodium, and Potassium
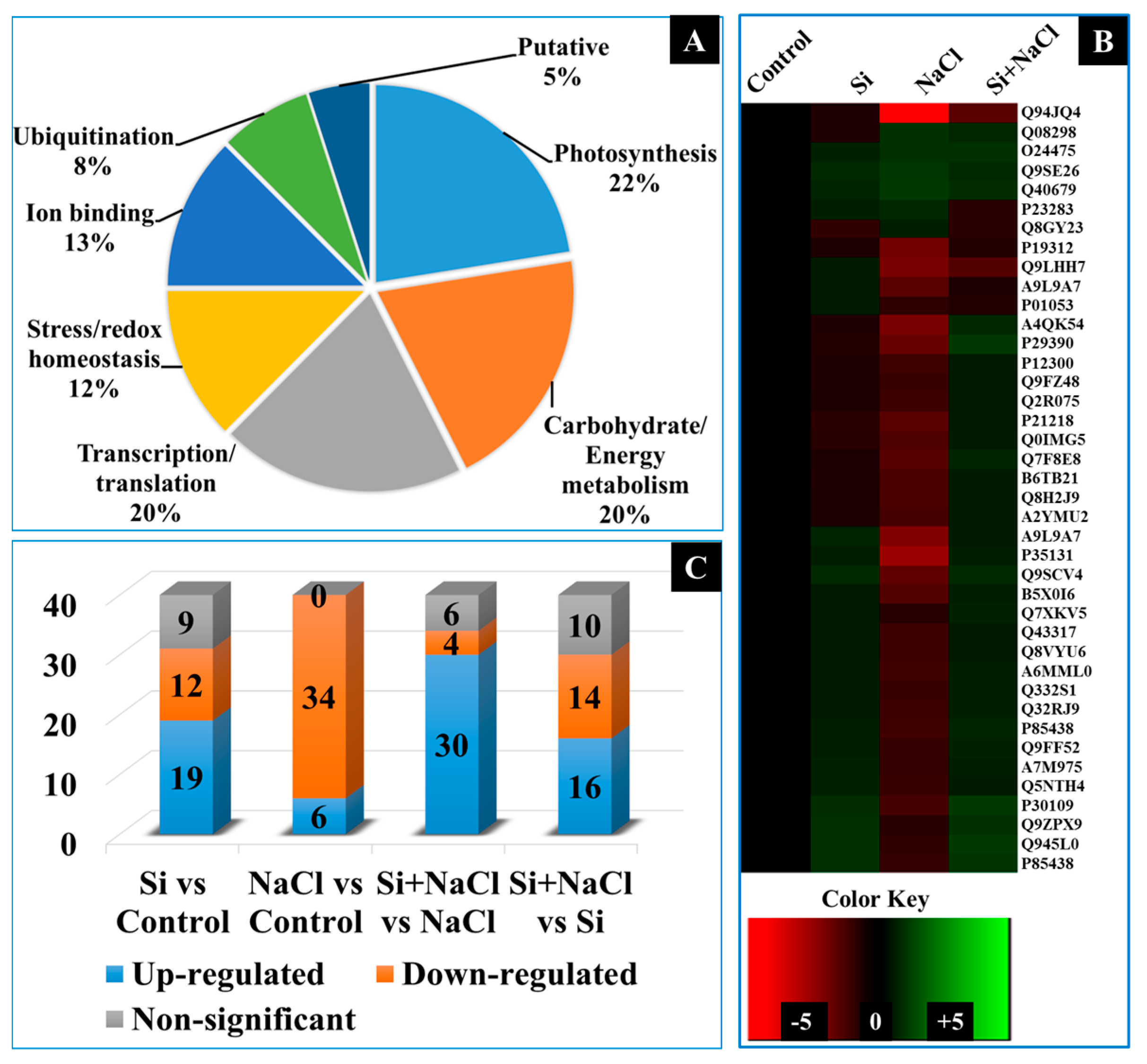
2.4. Dynamic Changes in the Expression of Proteins
2.4.1. Photosynthesis-Related Proteins
2.4.2. Proteins Related to Energy Metabolism
2.4.3. Proteins Related to Transcription and Translation
2.4.4. Proteins Involved in Redox Homeostasis
2.4.5. Proteins Involved in Ion Binding
2.4.6. Proteins Involved in Ubiquitination
3. Discussion
4. Experimental Section
4.1. Plant Materials, Treatments, and Growth Conditions
4.2. Microscopy Observation and Pigment Analysis
4.3. Reactive Oxygen Species and Lipid Peroxidation
4.4. Analysis of Antioxidant Enzymes
4.4.1. Activity and Native-PAGE Analysis of Antioxidant Enzymes
4.4.2. Immunoblotting
4.5. Analysis of Silicon, Sodium, and Potassium Uptake
4.6. Proteomics Analysis
4.6.1. Protein Isolation
4.6.2. Isoelectric Focusing (IEF)
4.6.3. Two-Dimensional Gel Electrophoresis and Silver Staining
4.6.4. Protein Identification and Database Search
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ rations. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Bojorquez-Quintal, E.; Velarde-Buendia, A.; Ku-Gonzalez, A.; Carillo-Pech, M.; Ortega-Camacho, D.; Echevarria-Machado, I.; Pottosin, I.; Martinez-Estevez, M. Mechanisms of salt tolerance in habanero pepper plants (Capsicum chinense Jacq.): Proline accumulation, ions dynamics and sodium root-shoot partition and compartmentation. Front. Plant Sci. 2014, 5, 1–14. [Google Scholar]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef]
- Davies, K.J. An overview of oxidative stress. IUBMB Life 2000, 50, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.D. Dissection of oxidative stress tolerance using transgenic plants. Plant Physiol. 1995, 107, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, R.I.; Perdomo, P. Reassessing the salinity tolerance of greenhouse roses under soilless production conditions. HortScience 2003, 38, 533–536. [Google Scholar]
- Wahome, P.K.; Jesch, H.H.; Grittner, I. Mechanisms of salt stress tolerance in two rose rootstocks: Rosa chinensis ‘Major’and R. rubiginosa. Sci. Hort. 2001, 87, 207–216. [Google Scholar] [CrossRef]
- Bernstein, L. Salinity and roses. Am. Rose Ann. 1964, 49, 120–124. [Google Scholar]
- Fernández Falcón, M.I.; Alvarez, C.E.A.; Garcia, V.; Baez, J. The effect of chloride and bicarbonate levels in irrigation water on nutrient content, production, and quality of cut roses ‘Mercedes’. Sci. Hort. 1986, 29, 373–385. [Google Scholar] [CrossRef]
- Niu, G.; Rodriguez, D.S.; Aguiniga, L. Effect of saline water irrigation on growth and physiological responses of three rose rootstocks. HortScience 2008, 43, 1479–1484. [Google Scholar] [PubMed]
- Cabrera, R.I.; Alma, R.S.; Sloan, J.J. Greenhouse rose yield and ion accumulation responses to salt stress as modulated by rootstock selection. HortScience 2009, 44, 2000–2008. [Google Scholar]
- Hwang, S.J.; Jeong, B.R.; Park, H.M. Effects of Potassium Silicate on the Growth of Miniature Rose ‘Pinocchio’ Grown on Rockwool and its Cut Flower Quality. J. Jpn. Soc. Hort. Sci. 2005, 74, 242–247. [Google Scholar] [CrossRef]
- Gillman, J.H.; Zlesak, D.C.; Smith, J.A. Applications of potassium silicate decrease black spot infection in Rosa hybrida ‘Meipelta’ (Fuchsia MeidilandTM). HortScience 2003, 38, 1144–1147. [Google Scholar]
- Larsen, A.K. Less mildew in pot roses with silicon. FlowerTech 2008, 11, 18–19. [Google Scholar]
- Epstein, E. Silicon. Annu. Rev. Plant Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhu, X.; Chen, K.; Wang, S.; Zhang, C. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2005, 169, 309–314. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum ‘Bugwang’. BioMed Res. Internat. 2016, 3076357. [Google Scholar] [CrossRef]
- Soundararajan, P.; Sivanesan, I.; Jana, S.; Jeong, B.R. Influence of silicon supplementation on the growth and tolerance to high temperature in Salvia splendens. Hortic. Environ. Biotechnol. 2014, 55, 271–279. [Google Scholar] [CrossRef]
- Fauteux, F.; Chain, F.; Belzile, F.; Menzies, J.G.; Bélanger, R.R. The protective role of silicon in the Arabidopsis-powdery mildew pathosystem. Proc. Natl. Acad. Sci. USA 2006, 103, 17554–17559. [Google Scholar] [CrossRef] [PubMed]
- Keeping, M.G.; Kvedaras, O.L. Silicon as a plant defence against insect herbivory: Response to Massey, Ennos and Hartley. J. Animal Ecol. 2008, 77, 631–633. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Agarie, S.; Agata, W.; Kubota, H.; Kaufmann, P.B. Physiological role of silicon in photosynthesis and dry matter production in rice plants. Jpn. J. Crop Sci. 1992, 61, 200–206. [Google Scholar] [CrossRef]
- Yeo, A.R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N.; Flowers, T.J. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpirational bypass flow. Plant Cell Environ. 1999, 22, 559–565. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Ahmad, R.; Zaheer, S.H.; Ismail, S. Role of silicon in salt tolerance of wheat (Triticum aestivum L.). Plant Sci. 1992, 85, 43–50. [Google Scholar] [CrossRef]
- Savvas, D.; Gizas, G.; Karras, G.; Lydakis-Simantiris, N.; Salahas, G.; Papadimitriou, M.; Tsouka, N. Interactions between silicon and NaCl-salinity in a soilless culture of roses in greenhouse. Eur. J. Hortic. Sci. 2007, 72, 73–79. [Google Scholar]
- Reezi, S.; Babalar, M.; Kalantari, S. Silicon alleviates salt stress, decreases malondialdehyde content and affects petal color of salt stressed cut rose (Rosa x hybrida L.) ‘Hot Lady’. Af. J. Biotechnol. 2009, 8, 1502–1508. [Google Scholar]
- Jorrin-Novo, J.V.; Pascual, J.; Sánchez-Lucas, R.; Romero-Rodríguez, M.C.; Rodríguez-Ortega, M.J.; Lenz, C.; Valledor, L. Fourteen years of plant proteomics reflected in Proteomics: Moving from model species and 2DE-based approaches to orphan species and gel-free platforms. Proteomics 2015, 15, 1089–1112. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Jeong, B.R. Proteomic analysis of salt-stress responsive proteins in roots of tomato (Lycopersicon esculentum L.) plants towards silicon efficiency. Plant Growth Regul. 2015, 77, 133–146. [Google Scholar] [CrossRef]
- MASCOT Software. Available online: http://www.matrixscience.com/ (accessed on 15 May 2015).
- Nwugo, C.C.; Huerta, A.J. The effect of silicon on the leaf proteome of rice (Oryza sativa L.) Plants under Cadmium-Stress. J. Proteome Res. 2011, 10, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.M.P.; Roberts, T.H.; Moller, I.M. Evidence for the presence of two rotenone-insensitive NAD(P)H dehydrogenases on the inner surface of the inner membrane of potato tuber mitochondria. Biochim. Biophys. Acta 1996, 1276, 133–139. [Google Scholar] [CrossRef]
- Lux, A.; Luxová, M.; Abe, J.; Tanimoto, E.; Hattori, T.; Inanaga, S. The dynamics of silicon deposition in the sorghum root endodermis. New Phytol. 2003, 158, 437–441. [Google Scholar] [CrossRef]
- Sivanesan, I.; Son, M.S.; Soundararajan, P.; Jeong, B.R. Effect of silicon on growth and temperature stress tolerance of Nephrolepis exaltata ‘Corditas’. Korean J. Hortic. Sci. 2014, 32, 142–148. [Google Scholar] [CrossRef]
- Pospisilova, J.; Solarova, J.; Catsky, J. Photosynthetic responses to stresses during in vitro cultivation. Photosynthetica 1996, 26, 3–18. [Google Scholar]
- Melo, A.M.; Bandeiras, T.M.; Teixeira, M. New insights into type II NAD(P)H: Quinone oxidoreductases. Microbiol. Mol. Biol. Rev. 2004, 68, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, N.; Ceccarelli, E.A. Open questions in ferredoxin-NADP+ reductase catalytic mechanism. Eur. J. Biochem. 2003, 270, 1900–1915. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, M.A.; Ceccarelli, A.; Catalano-Dupuy, D.L. The Plant–Type Ferredoxin-NADP+ Reductases. In Advances in Photosynthesis-Fundamental Aspects; Najafpour, M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 539–562. [Google Scholar]
- Spreitzer, R.J. Role of the small subunit in ribulose-1, 5-bisphosphate carboxylase/oxygenase. Arch. Biochem. Biophys. 2003, 414, 141–149. [Google Scholar] [CrossRef]
- Krech, K.; Ruf, S.; Masduki, F.F.; Thiele, W.; Bednarczyk, D.; Albus, C.A.; Bock, R. The plastid genome-encoded Ycf4 protein functions as a nonessential assembly factor for photosystem I in higher plants. Plant Physiol. 2012, 159, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Kluge, M.; Ting, I.P. Crassulacean Acid Metabolism: Analysis of an Ecological Adaptation; Springer Science & Business Media: New York, NY, USA, 1978. [Google Scholar]
- Sasaki, Y.; Nagano, Y. Plant acetyl-CoA carboxylase: Structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Botechnol. Biochem. 2004, 68, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, A.G.; Geisler, D.A.; Møller, I.M. The multiplicity of dehydrogenases in the electron transport chain of plant mitochondria. Mitochondrion 2008, 8, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.C. Chromatin modifiers that control plant development. Curr. Opin. Plant Biol. 2006, 9, 21–27. [Google Scholar] [CrossRef]
- Sobhanian, H.; Razavizadeh, R.; Nanjo, Y.; Ehsanpour, A.A.; Jazil, F.R.; Motamed, N.; Komatsu, S. Proteome analysis of soybean leaves, hypocotyls and roots under salt stress. Proteome Sci. 2010, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Derry, M.C.; Yanagiya, A.; Martineau, Y.; Sonenberg, N. Regulation of poly(A)-binding protein through PABP-interacting proteins. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Niehaus, T.D.; Nguyen, T.N.; Gidda, S.K.; ElBadawi-Sidhu, M.; Lambrecht, J.A.; McCarty, D.R.; Hanson, A.D. Arabidopsis and maize RidA proteins preempt reactive enamine/imine damage to branched-chain amino acid biosynthesis in plastids. Plant Cell 2014, 26, 3010–3022. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, M.; Hell, R. Functional analysis of the cysteine synthase protein complex from plants: Structural, biochemical and regulatory properties. J. Plant Physiol. 2006, 163, 273–286. [Google Scholar] [CrossRef]
- Kasai, K.; Kanno, T.; Akita, M.; Ikejiri-Kanno, Y.; Wakasa, K.; Tozawa, Y. Identification of three shikimate kinase genes in rice: Characterization of their differential expression during panicle development and of the enzymatic activities of the encoded proteins. Planta 2005, 222, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Janosi, L.; Hara, H.; Zhang, S.; Kaji, A. Ribosome recycling by ribosome recycling factor (RRF)—An important but overlooked step of protein biosynthesis. Adv. Biophys. 1996, 32, 121–201. [Google Scholar] [CrossRef]
- Kraft, E.; Stone, S.L.; Ma, L.; Su, N.; Gao, Y.; Lau, O.S.; Callis, J. Genome analysis and functional characterization of the E2 and RING-type E3 ligase ubiquitination enzymes of Arabidopsis. Plant Physiol. 2005, 139, 1597–1611. [Google Scholar] [CrossRef] [PubMed]
- Marrs, K.A. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef] [PubMed]
- Hanano, A.; Burcklen, M.; Flenet, M. Plant seed peroxygenase is an original heme-oxygenase with an EF-hand calcium binding motif. J. Biol. Chem. 2006, 281, 33140–33151. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, P.; Dommes, J.; Hoffmann, L.; Hausman, J.F.; Renaut, J. Quantitative changes in protein expression of cadmium exposed poplar plants. Proteomics 2008, 8, 2514–2530. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Boscaro, F.; Chatzi, A.; Mikolajczyk, M.; Tokatlidis, K.; Winkelmann, J. Anamorsin is a [2Fe-2S] cluster-containing substrate of the Mia40-dependent mitochondrial protein trapping machinery. Chem. Biol. 2011, 18, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xu, Y.; Li, J.; Yang, L.; Liu, J.Y. Molecular analyses of the metallothionein gene family in rice (Oryza sativa L.). BMB Rep. 2006, 39, 595–606. [Google Scholar] [CrossRef]
- Lobreaux, S.; Massenet, O.; Briat, J.F. Iron induces ferritin synthesis in maize plantlets. Plant Mol. Biol. 1992, 19, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Jambunathan, N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants. Plant Stress Toler. 2010, 639, 291–297. [Google Scholar]
- Tian, J.; Jian, F.G.; Wu, Z. The apoplastic oxidative burst as a key factor of hyperhydricity in garlic plantlet in vitro. Plant Cell Tiss Org. 2015, 120, 517–584. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Elliott, C.L.; Snyder, G.H. Autoclave-induced digestion for the colorimetric determination of silicon in rice straw. J. Agric. Food Chem. 1999, 39, 1118–1119. [Google Scholar] [CrossRef]
- Westermeier, R. Preparation of plant samples for 2-D electrophoresis. Proteomics 2006, 2, 56–60. [Google Scholar] [CrossRef] [PubMed]
- AgBase. Available online: http://www.agbase.msstate.edu/ (accessed on 15 May 2015).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. a | Accession Number b | Nominal Mass (Mr) c | Theo./Exp. pI d | Protein Identification | Species | Sc (%) e | Score f |
|---|---|---|---|---|---|---|---|
| Photosynthesis | |||||||
| 7 | Q332S1 | 45577 | 5.43/5.47 | NAD(P)H-quinone oxidoreductase subunit H | Lactuca sativa | 84 | 40 |
| 24 | A6MML0 | 18820 | 6.19/5.05 | NAD(P)H-quinone oxidoreductase subunit J | Dioscorea elephantipes | 39 | 47 |
| 12 | Q7F8E8 | 40407 | 8.72/4.45 | Ferredoxin-NADP reductase | Oryza sativa Japonica | 18 | 20 |
| 10 | Q945L0 | 9476 | 5.33/5.10 | Cytochrome c oxidase subunit 6b-2 | Arabidopsis thaliana | 60 | 46 |
| 21 | A9L9A7 | 9.59/5.40 | 9.59/5.20 | Photosystem I assembly protein Ycf4 | Lemna minor | 36 | 40 |
| 28 | A9L9A7 | 21440 | 9.59/6.15 | Photosystem I assembly protein Ycf4 | Lemna minor | 36 | 40 |
| 23 | P19312 | 20087 | 7.60/6.50 | Ribulose bisphosphate carboxylase small chain SSU5B | Lemna gibba | 28 | 34 |
| 26 | P21218 | 43587 | 9.23/4.50 | Protochlorophyllide reductase B | Arabidopsis thaliana | 25 | 30 |
| 30 | A7M975 | 21497 | 9.59/4.10 | Photosystem I assembly protein Ycf4 | Cuscuta reflexa | 28 | 36 |
| Energy metabolism | |||||||
| 9 | Q7XKV5 | 60290 | 7.21/5.90 | β-glucosidase 11 | Oryza sativa Japonica | 19 | 35 |
| 15 | Q9SCV4 | 94293 | 8.09/5.10 | β -galactosidase 8 | Arabidopsis thaliana | 9 | 41 |
| 3 | Q9SE26 | 64295 | 7.28/4.00 | Isocitrate lyase | Dendrobium crumenatum | 20 | 40 |
| 25 | P12300 | 56038 | 6.61/6.70 | Glucose-1-phosphate adenylyltransferase large subunit | Triticum aestivum | 16 | 32 |
| 27 | Q8H2J9 | 47104 | 9.76/6.80 | Glycerol-3-phosphate dehydrogenase (NAD+) | Oryza sativa Japonica | 22 | 35 |
| 11 | P85438 | 3396 | 9.99/4.10 | Acetyl-CoA carboxylase | Catharanthus roseus | 100 | 36 |
| 16 | P85438 | 3398 | 9.99/5.10 | Acetyl-CoA carboxylase | Catharanthus roseus | 96 | 34 |
| 22 | Q9LHH7 | 31589 | 5.77/6.55 | Bifunctional protein FolD 2 | Arabidopsis thaliana | 38 | 51 |
| Transcription/translation | |||||||
| 20 | A4QK54 | 13682 | 9.39/4.55 | 50S ribosomal protein L14 | Arabis hirsuta | 54 | 38 |
| 32 | Q9FF52 | 17946 | 9.02/5.15 | 60S ribosomal protein L12-3 | Arabidopsis thaliana | 51 | 34 |
| 40 | B5X0I6 | 124486 | 5.20/4.45 | Protein CTR9 homolog | Arabidopsis thaliana | 15 | 36 |
| 5 | Q94JQ4 | 19930 | 8.53/5.25 | Reactive Intermediate Deaminase A | Arabidopsis thaliana | 66 | 38 |
| 39 | Q43317 | 34492 | 6.25/5.20 | Cysteine synthase | Citrullus lanatus | 40 | 40 |
| 36 | Q5NTH4 | 33310 | 9.30/4.30 | Shikimate kinase 1 | Oryza sativa Japonica | 34 | 45 |
| 14 | A2YMU2 | 29709 | 9.35/5.10 | Ribosome-recycling factor, | Oryza sativa Indica Group | 23 | 33 |
| 17 | Q32RJ9 | 49484 | 9.55/5.87 | tRNA(Ile)-lysidine synthase | Zygnema circumcarinatum | 13 | 32 |
| Stress/redox homeostasis | |||||||
| 8 | P23283 | 21940 | 8.91/4.00 | Desiccation-related protein PCC3-06 | Craterostigma plantagineum | 38 | 33 |
| 18 | Q08298 | 42658 | 9.42/4.60 | Dehydration-responsive protein RD22 | Elaeis guineensis var. tenera | 20 | 28 |
| 37 | P01053 | 9381 | 6.58/6.15 | Subtilisin-chymotrypsin inhibitor-2A | Hordeum vulgare | 38 | 27 |
| 19 | Q40679 | 27454 | 6.00/4.00 | Peroxygenase | Oryza sativa Indica | 25 | 29 |
| 13 | P30109 | 24081 | 5.54/6.80 | Glutathione S-transferase PARB | Nicotiana tabacum | 41 | 36 |
| Ion binding | |||||||
| 2 | Q9ZPX9 | 15339 | 4.34/6.80 | Calcium-binding protein KIC | Arabidopsis thaliana | 28 | 19 |
| 6 | B6TB21 | 28001 | 5.20/4.45 | Anamorsin homolog | Zea mays | 48 | 30 |
| 31 | O24475 | 72088 | 5.48/4.00 | Pinene synthase | Abies grandis | 14 | 28 |
| 33 | Q0IMG5 | 8379 | 5.62/5.60 | Metallothionein-like protein 4A | Oryza sativa Japonica | 69 | 29 |
| 34 | P29390 | 27863 | 5.75/4.31 | Ferritin-2 | Zea mays | 23 | 36 |
| Ubiquitination | |||||||
| 4 | Q9FZ48 | 17277 | 6.74/4.47 | Ubiquitin-conjugating enzyme E2 8 | Arabidopsis thaliana | 59 | 32 |
| 38 | P35131 | 17277 | 6.74/4.60 | Ubiquitin-conjugating enzyme E2 36 | Arabidopsis thaliana | 59 | 32 |
| 35 | Q8GY23 | 404995 | 4.96/4.45 | E3 ubiquitin-protein ligase | Arabidopsis thaliana | 17 | 34 |
| Putative | |||||||
| 1 | Q9FKQ2 | 13622 | 8.53/5.38 | Putative clathrin assembly protein | Pinus koraiensis | 57 | 35 |
| 29 | Q2R075 | 43258 | 6.62/5.55 | Putative glutaredoxin-C11 | Oryza sativa Indica | 24 | 34 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soundararajan, P.; Manivannan, A.; Ko, C.H.; Muneer, S.; Jeong, B.R. Leaf Physiological and Proteomic Analysis to Elucidate Silicon Induced Adaptive Response under Salt Stress in Rosa hybrida ‘Rock Fire’. Int. J. Mol. Sci. 2017, 18, 1768. https://doi.org/10.3390/ijms18081768
Soundararajan P, Manivannan A, Ko CH, Muneer S, Jeong BR. Leaf Physiological and Proteomic Analysis to Elucidate Silicon Induced Adaptive Response under Salt Stress in Rosa hybrida ‘Rock Fire’. International Journal of Molecular Sciences. 2017; 18(8):1768. https://doi.org/10.3390/ijms18081768
Chicago/Turabian StyleSoundararajan, Prabhakaran, Abinaya Manivannan, Chung Ho Ko, Sowbiya Muneer, and Byoung Ryong Jeong. 2017. "Leaf Physiological and Proteomic Analysis to Elucidate Silicon Induced Adaptive Response under Salt Stress in Rosa hybrida ‘Rock Fire’" International Journal of Molecular Sciences 18, no. 8: 1768. https://doi.org/10.3390/ijms18081768
APA StyleSoundararajan, P., Manivannan, A., Ko, C. H., Muneer, S., & Jeong, B. R. (2017). Leaf Physiological and Proteomic Analysis to Elucidate Silicon Induced Adaptive Response under Salt Stress in Rosa hybrida ‘Rock Fire’. International Journal of Molecular Sciences, 18(8), 1768. https://doi.org/10.3390/ijms18081768