Burn Eschar Stimulates Fibroblast and Adipose Mesenchymal Stromal Cell Proliferation and Migration but Inhibits Endothelial Cell Sprouting


Abstract
:1. Introduction
2. Results
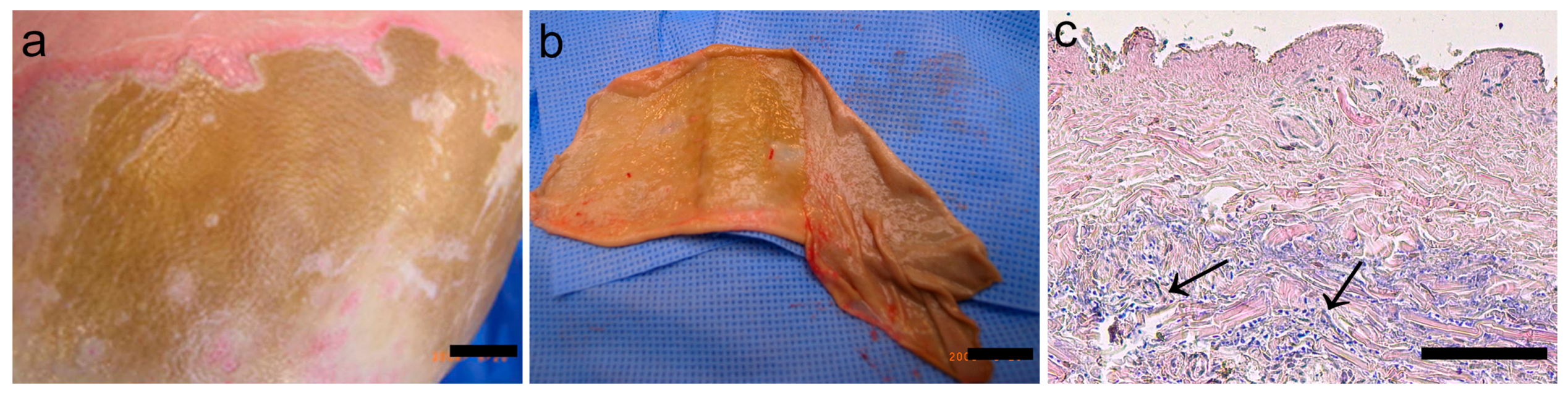
2.1. Eschar
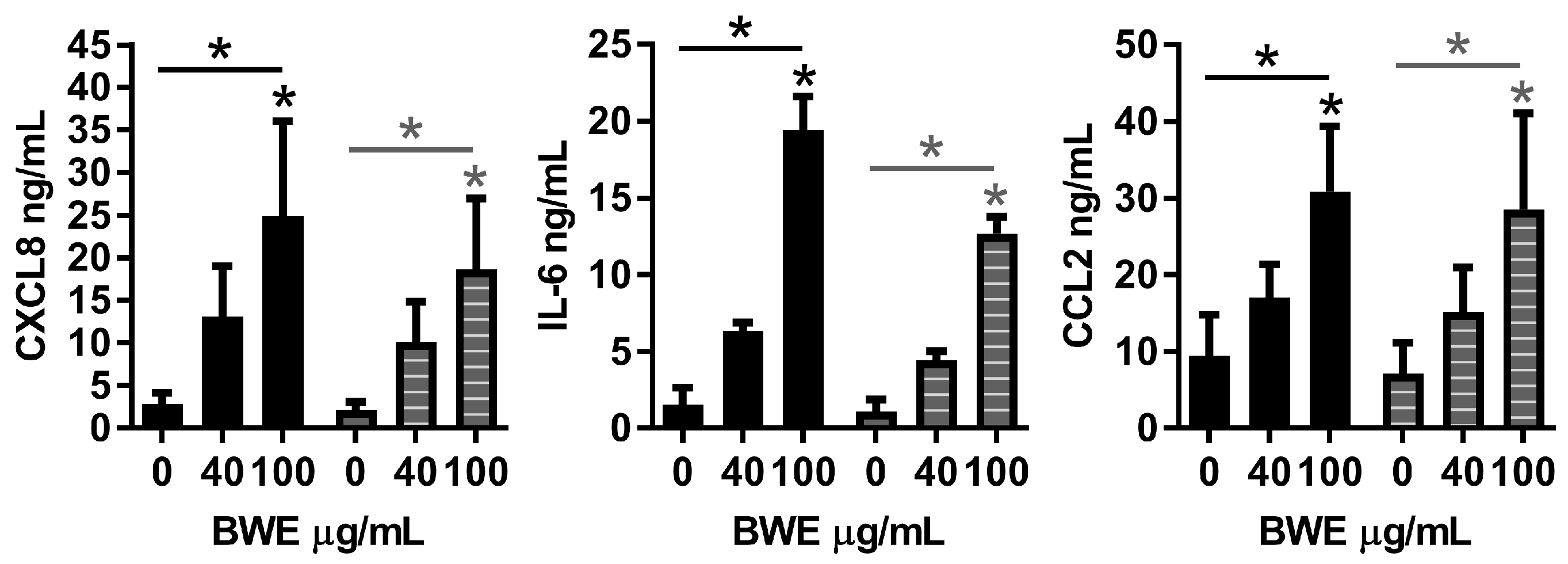
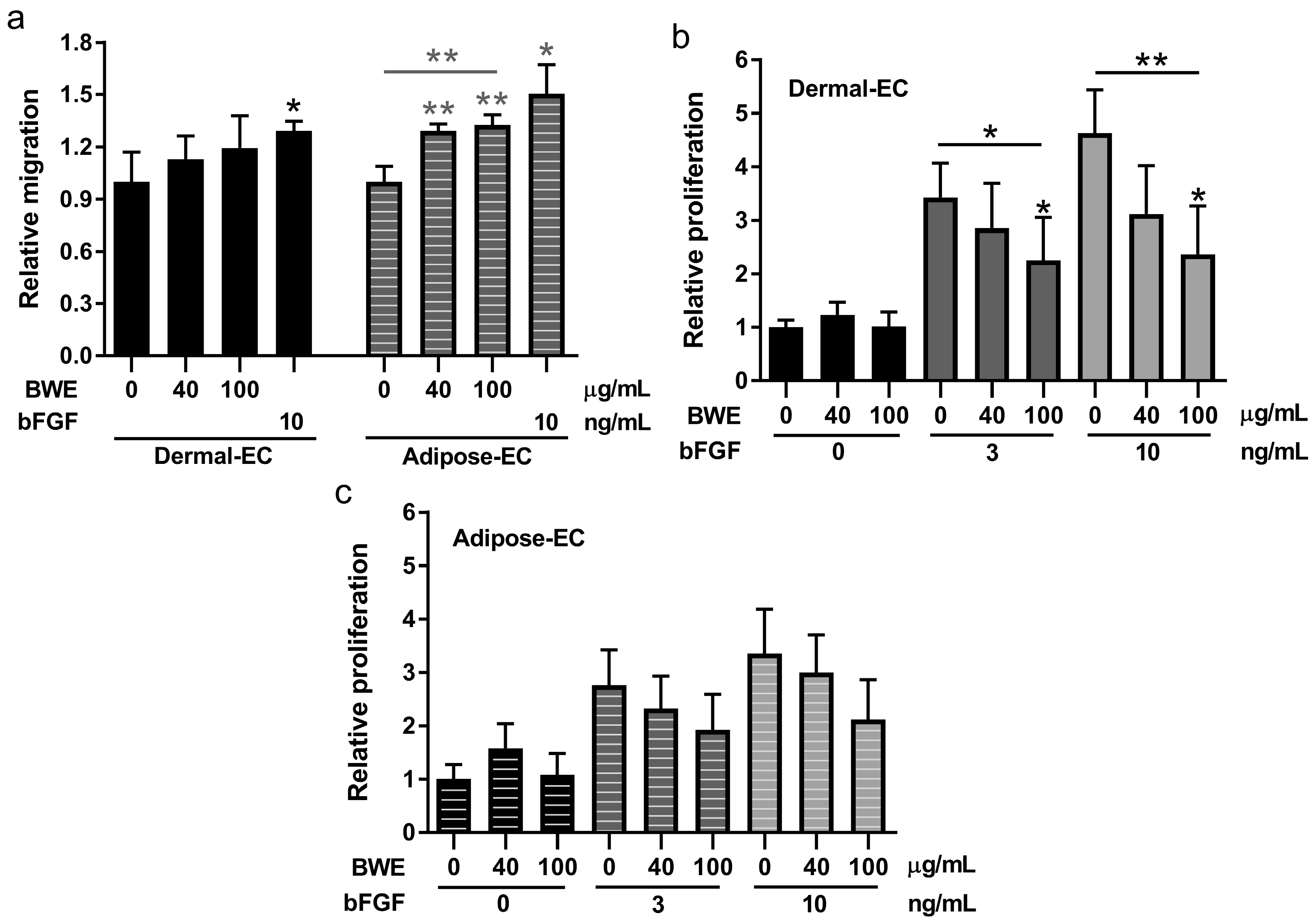
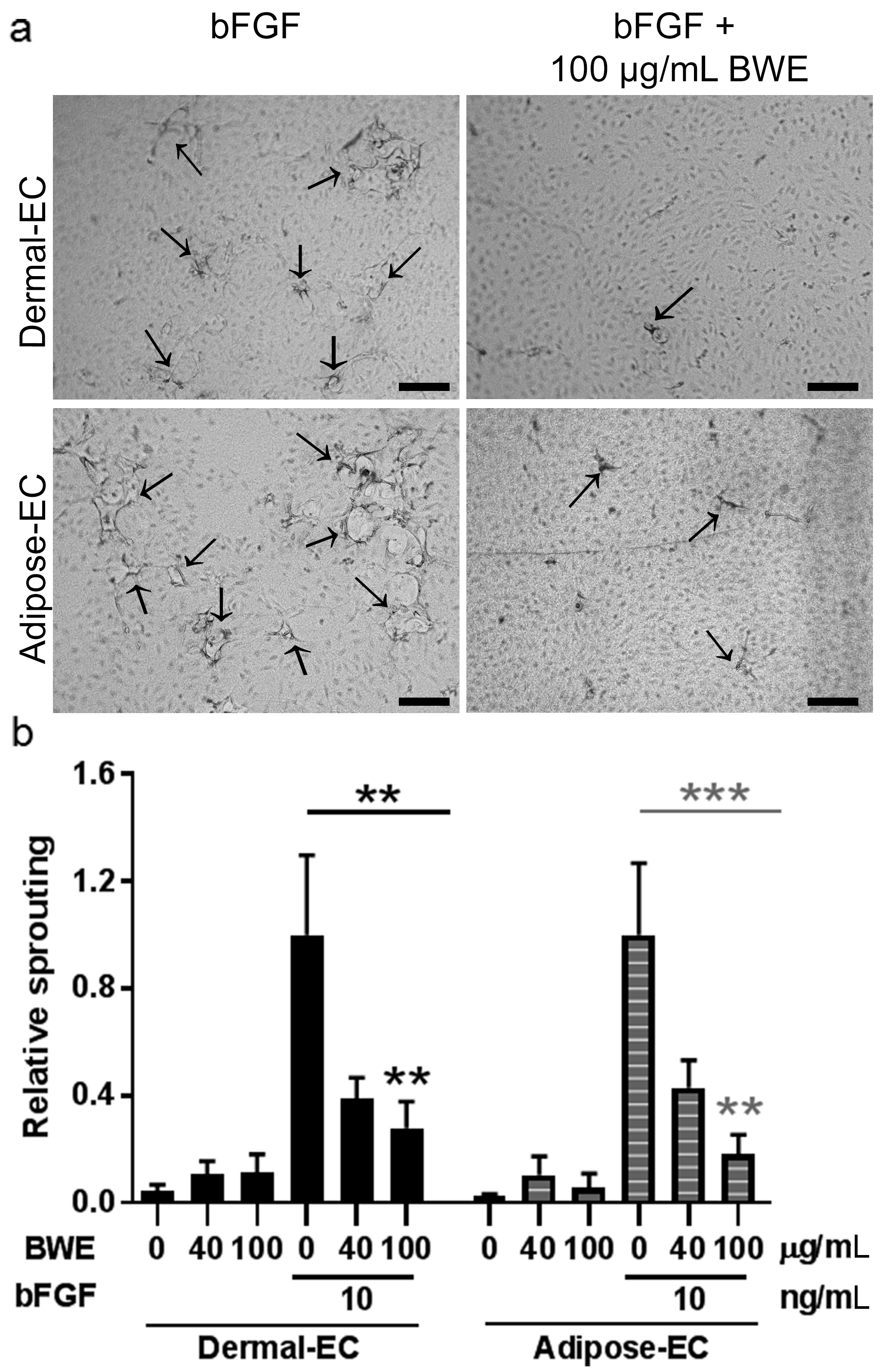
2.2. Burn Wound Extract Inhibits Endothelial Cell Proliferation and Sprouting
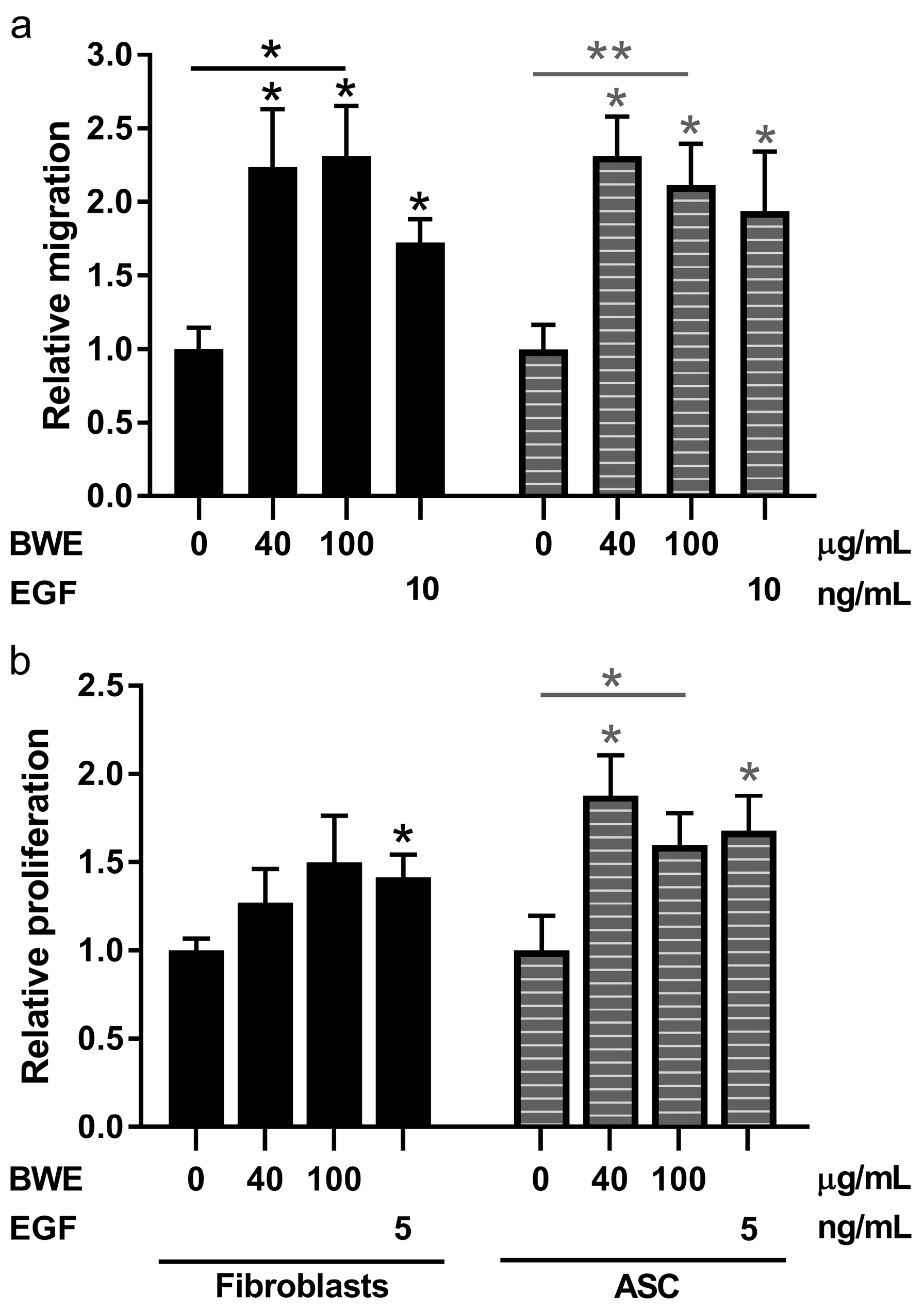
2.3. Burn Wound Extract Stimulates Both Migration and Proliferation of Fibroblasts and ASC
3. Discussion
4. Materials and Methods
4.1. Human Tissue
4.2. Burn Wound Extract
4.3. Cell Culture
4.4. Exposure of Endothelial Cells to BWE
4.5. Cell Migration Assay
4.6. Proliferation Assay
4.7. In Vitro Sprouting Assay
4.8. Histological Staining
4.9. Secretion of Cytokines and Chemokines
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ASC | Adipose tissue-derived mesenchymal stromal cell |
| bFGF | Basic fibroblast growth factor |
| BWE | Burn wound extract |
| EC | Endothelial cell |
| EGF | Epidermal growth factor |
| HE | Hematoxylin and eosin |
| PIC | Protease inhibitor cocktail |
| TBSA | Total body surface area |
References
- Van der Wal, M.B.; Vloemans, J.F.; Tuinebreijer, W.E.; van de Ven, P.; van Unen, E.; van Zuijlen, P.P.; Middelkoop, E. Outcome after burns: An observational study on burn scar maturation and predictors for severe scarring. Wound Repair Regen. 2012, 20, 676–687. [Google Scholar] [CrossRef] [PubMed]
- DeSanti, L. Pathophysiology and current management of burn injury. Adv. Skin Wound Care 2005, 18, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, M.C.; van der Veer, W.M.; Ulrich, M.M.; van Zuijlen, P.P.; Niessen, F.B.; Middelkoop, E. Prevention and curative management of hypertrophic scar formation. Burns 2009, 35, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Van der Veer, W.M.; Bloemen, M.C.; Ulrich, M.M.; Molema, G.; van Zuijlen, P.P.; Middelkoop, E.; Niessen, F.B. Potential cellular and molecular causes of hypertrophic scar formation. Burns 2009, 35, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Van den Bogaerdt, A.J.; van der Veen, V.C.; van Zuijlen, P.P.; Reijnen, L.; Verkerk, M.; Bank, R.A.; Middelkoop, E.; Ulrich, M.M. Collagen cross-linking by adipose-derived mesenchymal stromal cells and scar-derived mesenchymal cells: Are mesenchymal stromal cells involved in scar formation? Wound Repair Regen. 2009, 17, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Van den Broek, L.J.; Niessen, F.B.; Scheper, R.J.; Gibbs, S. Development, validation and testing of a human tissue engineered hypertrophic scar model. ALTEX 2012, 29, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Engrav, L.H.; Gibran, N.S.; Yang, T.M.; Grant, J.H.; Yunusov, M.Y.; Fang, P.; Reichenbach, D.D.; Heimbach, D.M.; Isik, F.F. Cones of skin occur where hypertrophic scar occurs. Wound Repair Regen. 2001, 9, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Van der Veer, W.M.; Niessen, F.B.; Ferreira, J.A.; Zwiers, P.J.; de Jong, E.H.; Middelkoop, E.; Molema, G. Time course of the angiogenic response during normotrophic and hypertrophic scar formation in humans. Wound Repair Regen. 2011, 19, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Amadeu, T.; Braune, A.; Mandarim-de-Lacerda, C.; Porto, L.C.; Desmouliere, A.; Costa, A. Vascularization pattern in hypertrophic scars and keloids: A stereological analysis. Pathol. Res. Pract. 2003, 199, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Widgerow, A.D.; King, K.; Tocco-Tussardi, I.; Banyard, D.A.; Chiang, R.; Awad, A.; Afzel, H.; Bhatnager, S.; Melkumyan, S.; Wirth, G.; et al. The burn wound exudate-an under-utilized resource. Burns 2015, 41, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Van den Broek, L.J.; Kroeze, K.L.; Waaijman, T.; Breetveld, M.; Sampat-Sardjoepersad, S.C.; Niessen, F.B.; Middelkoop, E.; Scheper, R.J.; Gibbs, S. Differential response of human adipose tissue-derived mesenchymal stem cells, dermal fibroblasts, and keratinocytes to burn wound exudates: Potential role of skin-specific chemokine ccl27. Tissue Eng. Part A 2014, 20, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Van den Bogaerdt, A.J.; van Zuijlen, P.P.; van Galen, M.; Lamme, E.N.; Middelkoop, E. The suitability of cells from different tissues for use in tissue-engineered skin substitutes. Arch. Dermatol. Res. 2002, 294, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Monsuur, H.N.; Weijers, E.M.; Niessen, F.B.; Gefen, A.; Koolwijk, P.; Gibbs, S.; van den Broek, L.J. Extensive characterization and comparison of endothelial cells derived from dermis and adipose tissue: Potential use in tissue engineering. PLoS ONE 2016, 11, e0167056. [Google Scholar] [CrossRef] [PubMed]
- Kroeze, K.L.; Jurgens, W.J.; Doulabi, B.Z.; van Milligen, F.J.; Scheper, R.J.; Gibbs, S. Chemokine-mediated migration of skin-derived stem cells: Predominant role for ccl5/rantes. J. Investig. Dermatol. 2009, 129, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- Suffee, N.; Hlawaty, H.; Meddahi-Pelle, A.; Maillard, L.; Louedec, L.; Haddad, O.; Martin, L.; Laguillier, C.; Richard, B.; Oudar, O.; et al. Rantes/ccl5-induced pro-angiogenic effects depend on ccr1, ccr5 and glycosaminoglycans. Angiogenesis 2012, 15, 727–744. [Google Scholar] [CrossRef] [PubMed]
- De Smet, F.; Segura, I.; de Bock, K.; Hohensinner, P.J.; Carmeliet, P. Mechanisms of vessel branching: Filopodia on endothelial tip cells lead the way. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.C.; Wu, L.W.; Chen, C.L.; Shieh, S.J.; Chiu, H.Y. Deep partial thickness burn blister fluid promotes neovascularization in the early stage of burn wound healing. Wound Repair Regen. 2010, 18, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.C.; Wu, L.W.; Chen, C.L.; Shieh, S.J.; Chiu, H.Y. Angiogenin expression in burn blister fluid: Implications for its role in burn wound neovascularization. Wound Repair Regen. 2012, 20, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Nissen, N.N.; Gamelli, R.L.; Polverini, P.J.; DiPietro, L.A. Differential angiogenic and proliferative activity of surgical and burn wound fluids. J. Trauma Acute Care Surg. 2003, 54, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.M.; McGrouther, D.A.; Eastwood, M.; Brown, R.A. The effect of burn blister fluid on fibroblast contraction. Burns 1997, 23, 306–312. [Google Scholar] [CrossRef]
- Inoue, M.; Zhou, L.J.; Gunji, H.; Ono, I.; Kaneko, F. Effects of cytokines in burn blister fluids on fibroblast proliferation and their inhibition with the use of neutralizing antibodies. Wound Repair Regen. 1996, 4, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Topman, G.; Sharabani-Yosef, O.; Gefen, A. A standardized objective method for continuously measuring the kinematics of cultures covering a mechanically damaged site. Med. Eng. Phys. 2012, 34, 225–232. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Gender | Age | Cause of Burn | TBSA | Time after Injury (Days) | Protein Concentration BWE (μg/mL) | BWE Used in Figure |
|---|---|---|---|---|---|---|---|
| 1 | female | 49 | hot water | 9% | 21 | 2850 | F2 |
| 2 | female | 62 | hot object | 2% | 13 | 2420 | F2 |
| 3 | female | 30 | flame | 60% | 6 | 1120 | F2 |
| 4 | female | 73 | flame | 2% | 17 | 3260 | F4 |
| 5 | male | 49 | chemicals | 48% | 6 | 720 | F3,4,5 |
| 6 | male | 64 | hot object | 7% | 12 | 1780 | F3,4,5 |
| 7 | male | 45 | chemicals | 0.5% | 14 | 830 | F3,4,5 |
| 8 | female | 46 | hot object | 0.5% | 10 | 1620 | F3,4,5 |
| 9 | male | 52 | hot water | 2.5% | 10 | 2970 | F3,4,5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monsuur, H.N.; Van den Broek, L.J.; Jhingoerie, R.L.; Vloemans, A.F.P.M.; Gibbs, S. Burn Eschar Stimulates Fibroblast and Adipose Mesenchymal Stromal Cell Proliferation and Migration but Inhibits Endothelial Cell Sprouting. Int. J. Mol. Sci. 2017, 18, 1790. https://doi.org/10.3390/ijms18081790
Monsuur HN, Van den Broek LJ, Jhingoerie RL, Vloemans AFPM, Gibbs S. Burn Eschar Stimulates Fibroblast and Adipose Mesenchymal Stromal Cell Proliferation and Migration but Inhibits Endothelial Cell Sprouting. International Journal of Molecular Sciences. 2017; 18(8):1790. https://doi.org/10.3390/ijms18081790
Chicago/Turabian StyleMonsuur, Hanneke N., Lenie J. Van den Broek, Renushka L. Jhingoerie, Adrianus F. P. M. Vloemans, and Susan Gibbs. 2017. "Burn Eschar Stimulates Fibroblast and Adipose Mesenchymal Stromal Cell Proliferation and Migration but Inhibits Endothelial Cell Sprouting" International Journal of Molecular Sciences 18, no. 8: 1790. https://doi.org/10.3390/ijms18081790
APA StyleMonsuur, H. N., Van den Broek, L. J., Jhingoerie, R. L., Vloemans, A. F. P. M., & Gibbs, S. (2017). Burn Eschar Stimulates Fibroblast and Adipose Mesenchymal Stromal Cell Proliferation and Migration but Inhibits Endothelial Cell Sprouting. International Journal of Molecular Sciences, 18(8), 1790. https://doi.org/10.3390/ijms18081790