Dietary Phenethyl Isothiocyanate Protects Mice from Colitis Associated Colon Cancer

Abstract
:1. Introduction
2. Results
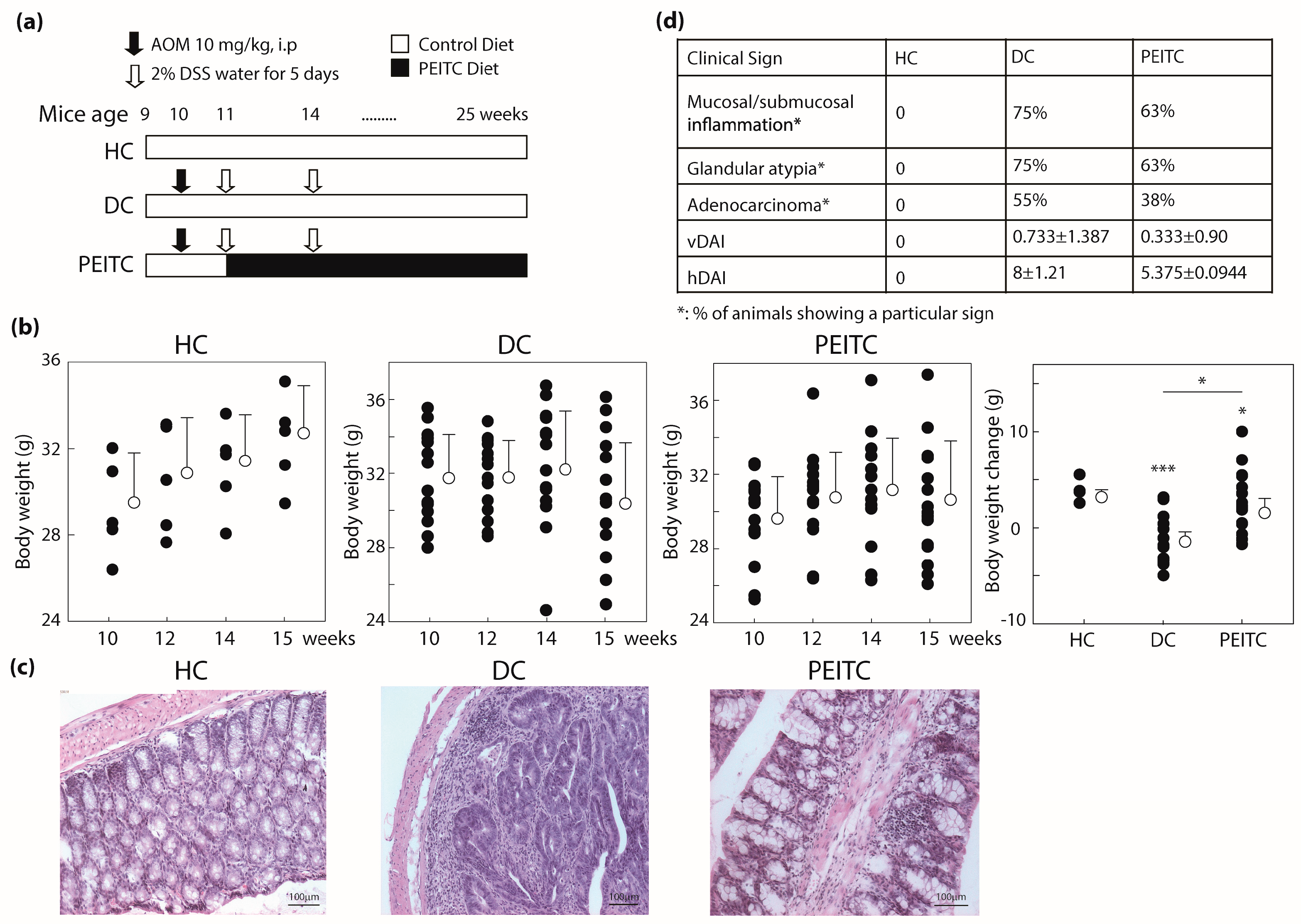
2.1. Phenethyl Isothiocyanate (PEITC) Treatment Improved Colon Cancer Associated Clinical Signs in Mice
2.2. Concentration Dependent Inhibition of Cell Survival by PEITC
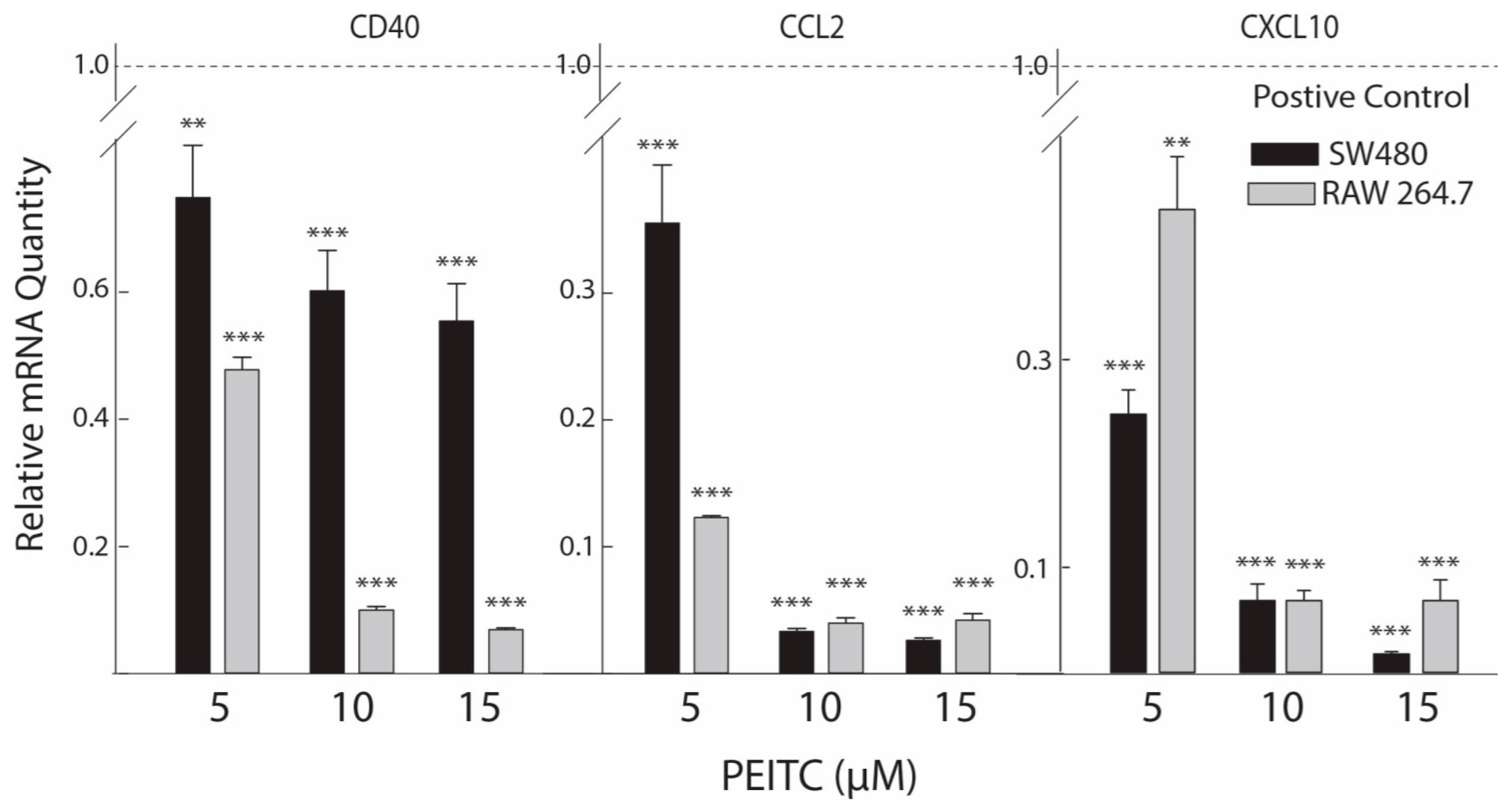
2.3. Reduced Expression of Inflammatory Mediators after PEITC Exposure
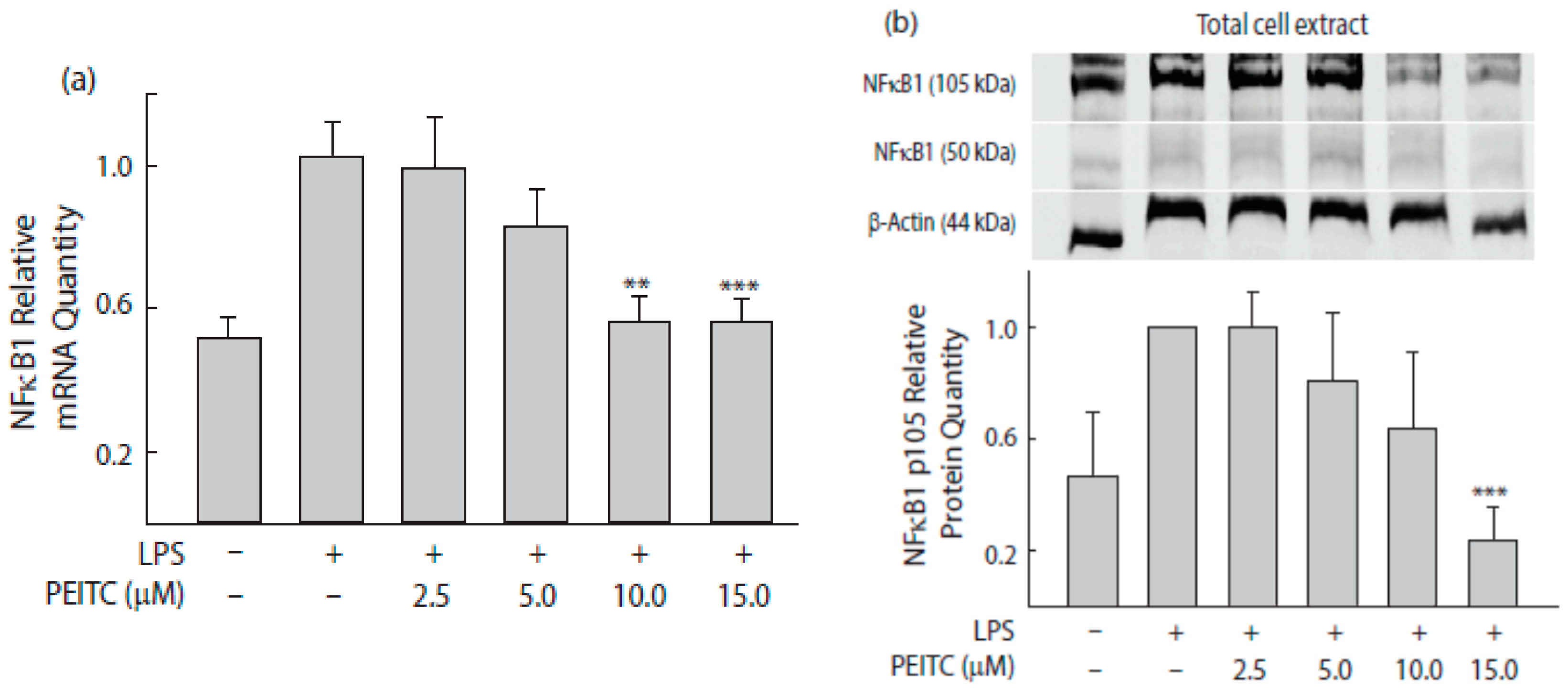
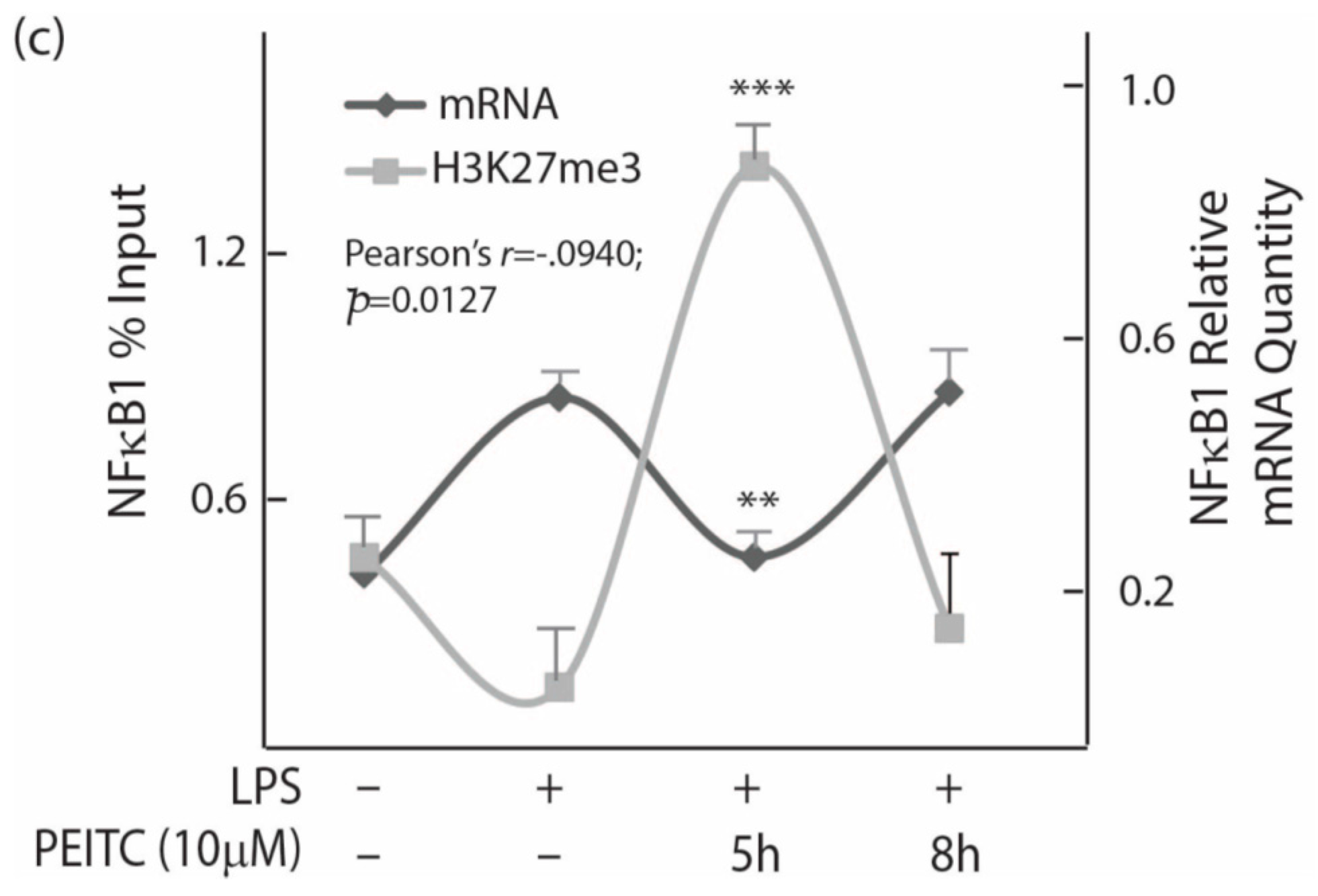
2.4. PEITC Exposure Attenuates NFκB1 Transcription Factor That Inversely Correlate with Tri-Methylation Levels of Lysine 27 on Histone 3 near Its Promoter
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. In Vivo Animal Studies
4.3. Cell Culture and PEITC Treatments
4.4. Total RNA Extraction, Purification, and cDNA Synthesis and Real-Time Quantitative PCR
4.5. Western Blot Analysis
4.6. ChIP Assay in Cell Culture
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AOM | Azoxymethane |
| CAC | Colitis Associated Cancer |
| CRC | Colorectal cancer |
| DSS | Dextran Sodium Sulfate |
| hDAI | histological Disease Activity Index |
| IBD | Inflammatory Bowel Disease |
| NFκB1 | Nuclear factor kappa B1 |
| PEITC | Phenethyl Isothiocyanate |
| UC | Ulcerative Colitis |
| vDAI | visible Disease Activity Index |
References
- Balch, C.; Ramapuram, J.B.; Tiwari, A.K. The Epigenomics of Embryonic Pathway Signaling in Colorectal Cancer. Front. Pharmacol. 2017, 8, 267. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Upadhyaya, B.; Liu, Y.; Knudsen, D.; Dey, M. Phenethyl isothiocyanate upregulates death receptors 4 and 5 and inhibits proliferation in human cancer stem-like cells. BMC Cancer 2014, 14, 591. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Impact and Application of Nutraceuticals on Inflammation-Induced Colorectal Cancer Development. In Molecular Oncology: Underlying Mechanisms and Translational Advancements; Farooqi, A.A., Ismail, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 273–291. [Google Scholar]
- Coope, A.; Pascoal, L.B.; da Silva, F.A.R.; Botezelli, J.D.; Ayrizono, M.L.S.; Milanski, M.; Camargo, M.G.; Planell, N.; Portovedo, M.; Dias, C.B.; et al. Transcriptional and Molecular Pathways Activated in Mesenteric Adipose Tissue and Intestinal Mucosa of Crohn’s Disease Patients. Int. J. Inflamm. 2017, 2017, 7646859. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chakravarty, S.; Dey, M. Phenethylisothiocyanate alters site- and promoter-specific histone tail modifications in cancer cells. PLoS ONE 2013, 8, e64535. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bhattacharya, R.K.; Roy, M. Targeting protein kinase C (PKC) and telomerase by phenethyl isothiocyanate (PEITC) sensitizes PC-3 cells towards chemotherapeutic drug-induced apoptosis. J. Environ. Pathol. Toxicol. Oncol. 2009, 28, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Wang, Z.Y. Breast cancer cell growth inhibition by phenethyl isothiocyanate is associated with down-regulation of oestrogen receptor-alpha36. J. Cell Mol. Med. 2010, 14, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Govind, S.; Sajankila, S.P.; Mi, L.; Roy, R.; Chung, F.L. Phenethyl isothiocyanate sensitizes human cervical cancer cells to apoptosis induced by cisplatin. Mol. Nutr. Food Res. 2011, 55, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Satyan, K.S.; Swamy, N.; Dizon, D.S.; Singh, R.; Granai, C.O.; Brard, L. Phenethyl isothiocyanate (PEITC) inhibits growth of ovarian cancer cells by inducing apoptosis: Role of caspase and MAPK activation. Gynecol. Oncol. 2006, 103, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, A.; Furukawa, F.; Lee, I.S.; Tanaka, T.; Hirose, M. Potent chemopreventive agents against pancreatic cancer. Curr. Cancer Drug Targets 2004, 4, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Wright, S.E.; Kim, S.H.; Srivastava, S.K. Phenethyl isothiocyanate: A comprehensive review of anti-cancer mechanisms. Biochim. Biophys. Acta 2014, 1846, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Chung, F.L.; Conaway, C.C.; Rao, C.V.; Reddy, B.S. Chemoprevention of colonic aberrant crypt foci in Fischer rats by sulforaphane and phenethyl isothiocyanate. Carcinogenesis 2000, 21, 2287–2291. [Google Scholar] [CrossRef] [PubMed]
- Plate, A.Y.; Gallaher, D.D. Effects of indole-3-carbinol and phenethyl isothiocyanate on colon carcinogenesis induced by azoxymethane in rats. Carcinogenesis 2006, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.L.; Khor, T.O.; Huang, M.T.; Kong, A.N. Differential in vivo mechanism of chemoprevention of tumor formation in azoxymethane/dextran sodium sulfate mice by PEITC and DBM. Carcinogenesis 2010, 31, 880–885. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, M.; Massi, E.; Poeta, M.L.; Carotti, S.; Morini, S.; Cecchetelli, L.; Signori, E.; Fazio, V.M. The AOM/DSS murine model for the study of colon carcinogenesis: From pathways to diagnosis and therapy studies. J. Carcinog. 2011, 10, 9. [Google Scholar] [PubMed]
- Dey, M.; Kuhn, P.; Ribnicky, D.; Premkumar, V.; Reuhl, K.; Raskin, I. Dietary phenethylisothiocyanate attenuates bowel inflammation in mice. BMC Chem. Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Thaker, A.I.; Shaker, A.; Rao, M.S.; Ciorba, M.A. Modeling colitis-associated cancer with azoxymethane (AOM) and dextran sulfate sodium (DSS). J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed]
- Bachmanov, A.A.; Reed, D.R.; Beauchamp, G.K.; Tordoff, M.G. Food intake, water intake, and drinking spout side preference of 28 mouse strains. Behav. Genet. 2002, 32, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Biomark. Prev. 2001, 10, 501–508. [Google Scholar]
- Waldner, M.J.; Neurath, M.F. Cytokines in colitis associated cancer: Potential drug targets? Inflamm. Allergy Drug Targets 2008, 7, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Hoeflinger, J.L.; Neme, B.P.; Jeffery, E.H.; Miller, M.J. Dietary Broccoli Alters Rat Cecal Microbiota to Improve Glucoraphanin Hydrolysis to Bioactive Isothiocyanates. Nutrients 2017, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Dey, M.; Ribnicky, D.; Kurmukov, A.G.; Raskin, I. In vitro and in vivo anti-inflammatory activity of a seed preparation containing phenethylisothiocyanate. J. Pharmacol. Exp. Ther. 2006, 317, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kong, W.; Jiang, J. Prevention and treatment of cancer targeting chronic inflammation: Research progress, potential agents, clinical studies and mechanisms. Sci. China Life Sci. 2017, 60, 601–616. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Floor, S.L.; Dumont, J.E.; Maenhaut, C.; Raspe, E. Hallmarks of cancer: Of all cancer cells, all the time? Trends Mol. Med. 2012, 18, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Upadhyaya, B.; Fardin-Kia, A.R.; Juenemann, R.M.; Dey, M. Dietary resistant starch type 4-derived butyrate attenuates nuclear factor-kappa-B1 through modulation of histone H3 trimethylation at lysine 27. Food Funct. 2016, 7, 3772–3781. [Google Scholar] [CrossRef] [PubMed]
- Rintisch, C.; Heinig, M.; Bauerfeind, A.; Schafer, S.; Mieth, C.; Patone, G.; Hummel, O.; Chen, W.; Cook, S.; Cuppen, E.; et al. Natural variation of histone modification and its impact on gene expression in the rat genome. Genome Res. 2014, 24, 942–953. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, B.; Larsen, T.; Barwari, S.; Louwagie, E.J.; Baack, M.L.; Dey, M. Prenatal Exposure to a Maternal High-Fat Diet Affects Histone Modification of Cardiometabolic Genes in Newborn Rats. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, G.; Wang, V.Y.; Huang, D.B.; Fusco, A. NF-kappaB regulation: Lessons from structures. Immunol. Rev. 2012, 246, 36–58. [Google Scholar] [CrossRef] [PubMed]
- Seufert, B.L.; Poole, E.M.; Whitton, J.; Xiao, L.; Makar, K.W.; Campbell, P.T.; Kulmacz, R.J.; Baron, J.A.; Newcomb, P.A.; Slattery, M.L.; et al. IkappaBKbeta and NFkappaB1, NSAID use and risk of colorectal cancer in the Colon Cancer Family Registry. Carcinogenesis 2013, 34, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.Y.; Zhu, Y.Q.; Wei, B.; Wang, H. Expression and functional research of TLR4 in human colon carcinoma. Am. J. Med. Sci. 2010, 339, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Asadzadeh-Aghdaee, H.; Shahrokh, S.; Norouzinia, M.; Hosseini, M.; Keramatinia, A.; Jamalan, M.; Naghibzadeh, B.; Sadeghi, A.; Jahani Sherafat, S.; Zali, M.R. Introduction of inflammatory bowel disease biomarkers panel using protein-protein interaction (PPI) network analysis. Gastroenterol. Hepatol. Bed Bench 2016, 9, S8–S13. [Google Scholar] [PubMed]
- Zhou, Y.; Zhou, S.X.; Gao, L.; Li, X.A. Regulation of CD40 signaling in colon cancer cells and its implications in clinical tissues. Cancer Immunol. Immunother. 2016, 65, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Chun, E.; Lavoie, S.; Michaud, M.; Gallini, C.A.; Kim, J.; Soucy, G.; Odze, R.; Glickman, J.N.; Garrett, W.S. CCL2 Promotes Colorectal Carcinogenesis by Enhancing Polymorphonuclear Myeloid-Derived Suppressor Cell Population and Function. Cell Rep. 2015, 12, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Wightman, S.C.; Uppal, A.; Pitroda, S.P.; Ganai, S.; Burnette, B.; Stack, M.; Oshima, G.; Khan, S.; Huang, X.; Posner, M.C.; et al. Oncogenic CXCL10 signalling drives metastasis development and poor clinical outcome. Br. J. Cancer 2015, 113, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Do, E.J.; Hwang, S.W.; Kim, S.Y.; Ryu, Y.M.; Cho, E.A.; Chung, E.J.; Park, S.; Lee, H.J.; Byeon, J.S.; Ye, B.D.; et al. Suppression of colitis-associated carcinogenesis through modulation of IL-6/STAT3 pathway by balsalazide and VSL#3. J. Gastroenterol. Hepatol. 2016, 31, 1453–1461. [Google Scholar] [PubMed]
- Hernandez, Y.; Sotolongo, J.; Breglio, K.; Conduah, D.; Chen, A.; Xu, R.; Hsu, D.; Ungaro, R.; Hayes, L.A.; Pastorini, C.; et al. The role of prostaglandin E2 (PGE 2) in toll-like receptor 4 (TLR4)-mediated colitis-associated neoplasia. BMC Gastroenterol. 2010, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PEITC (μM) | 6 h | 24 h | ||
|---|---|---|---|---|
| RAW264.7 | SW480 | RAW264.7 | SW480 | |
| 0 | 100 ± 0.066 | 100 ± 0.014 | 99.93 ± 0.067 | 100 ± 0.035 |
| 10 | 97.91 ± 0.072 | 101.14 ± 0.014 | 87.6 ± 1.25 ** | 78.26 ± 0.019 *** |
| 20 | 98.59 ± 0.074 | 94.24 ± 0.035 | 75.17 ± 1.95 *** | 78.04 ± 0.028 *** |
| 40 | 98.97 ± 0.12 | 95.48 ± 0.023 | 57.7 ± 0.55 *** | 64.44 ± 0.030 *** |
| Gene Name Abbreviation | Full Gene Name | Partial GO Term (Geneontology.Org) | % Suppression in RAW264.7 | % Suppression in SW480 |
|---|---|---|---|---|
| CCL2 | Chemokine (C–C motif) ligand 2 | Inflammatory response; Chemokine activity | 97.20 | 97.62 |
| CD40 | CD40 antigen | Signal transduction; Immune response; Apoptosis | 91.77 | 54.73 |
| CXCL10 | Chemokine (C–X–C motif) ligand 10 | Inflammatory response; Chemokine activity | 96.67 | 93.01 |
| NFκB1 | Nuclear factor of kappa light chain gene enhancer in B-cells 1, p105 | DNA binding; Regulation of transcription | 95.35 | 43.69 |
| NFκBiα | Nuclear factor of kappa light chain gene enhancer in B-cells inhibitor, alpha | Nucleus; Protein binding; Cytoplasm: Regulation of cell proliferation; Protein-nucleus import, translocation | 87.82 | 38.44 |
| REL | Reticuloendotheliosis oncogene | DNA binding; Regulation of transcription | 82.64 | 28.80 |
| RELβ | Avian reticuloendotheliosis viral (v-rel) oncogene related B | Transcription factor activity; Intracellular; T-helper 1 type immune response | 73.54 | 66.80 |
| RT-qPCR | ChIP-qPCR | |||
|---|---|---|---|---|
| RAW264.7 | SW480 | SW480 | ||
| CXCL10 | F: 5′-attctttaagggctggtctga-3′ | F: 5′-gaaagcagttagcaaggaaaggt-3′ | NA | |
| R: 5′-cacctccacatagcttacagt-3′ | R: 5′-gacatatactccatgtagggaagtga-3′ | |||
| CD40 | F:5′-acgagtcagactaatgtcatctgtg-3′ | F: 5′-ggtctcacctcgctatggtt-3′ | NA | |
| R:5′-ggtttcttgaccacctttttgat-3′ | R: 5′-cagtgggtggttctggatg-3′ | |||
| CCL2 | F: 5′-catccacgtgttggctca-3′ | F: 5′-agtctctgccgcccttct-3′ | NA | |
| R: 5′-gatcatcttgctggtgaatgagt-3′ | R: 5′-gtgactggggcattgattg-3′ | |||
| NFκB1 | F: 5′-gaggagaccggcaactca-3′ | F: 5′-accctgaccttgcctatttg-3′ | F:5′-ttggcaaaccccaaagag3′ | |
| R: 5′-gtccatctccttggtctgct-3′ | R: 5′-agctctttttcccgatctcc-3′ | R:5′-ggtttcccacgatcgattt-3′ | ||
| βActin/GAPDH | F: 5′-aaccgtgaaaagatgacccagat-3′ | F: 5′-agccacatcgctcagacac-3′ | NA | |
| R: 5′-cacagcctggatggctacgt-3′ | R: 5′-gcccaatacgaccaaatcc-3′ | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Dey, M. Dietary Phenethyl Isothiocyanate Protects Mice from Colitis Associated Colon Cancer. Int. J. Mol. Sci. 2017, 18, 1908. https://doi.org/10.3390/ijms18091908
Liu Y, Dey M. Dietary Phenethyl Isothiocyanate Protects Mice from Colitis Associated Colon Cancer. International Journal of Molecular Sciences. 2017; 18(9):1908. https://doi.org/10.3390/ijms18091908
Chicago/Turabian StyleLiu, Yi, and Moul Dey. 2017. "Dietary Phenethyl Isothiocyanate Protects Mice from Colitis Associated Colon Cancer" International Journal of Molecular Sciences 18, no. 9: 1908. https://doi.org/10.3390/ijms18091908
APA StyleLiu, Y., & Dey, M. (2017). Dietary Phenethyl Isothiocyanate Protects Mice from Colitis Associated Colon Cancer. International Journal of Molecular Sciences, 18(9), 1908. https://doi.org/10.3390/ijms18091908