Lactate Transporter Monocarboxylate Transporter 4 Induces Bone Pain in Head and Neck Squamous Cell Carcinoma

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MCT4 Expression in the Human HNSCC Samples
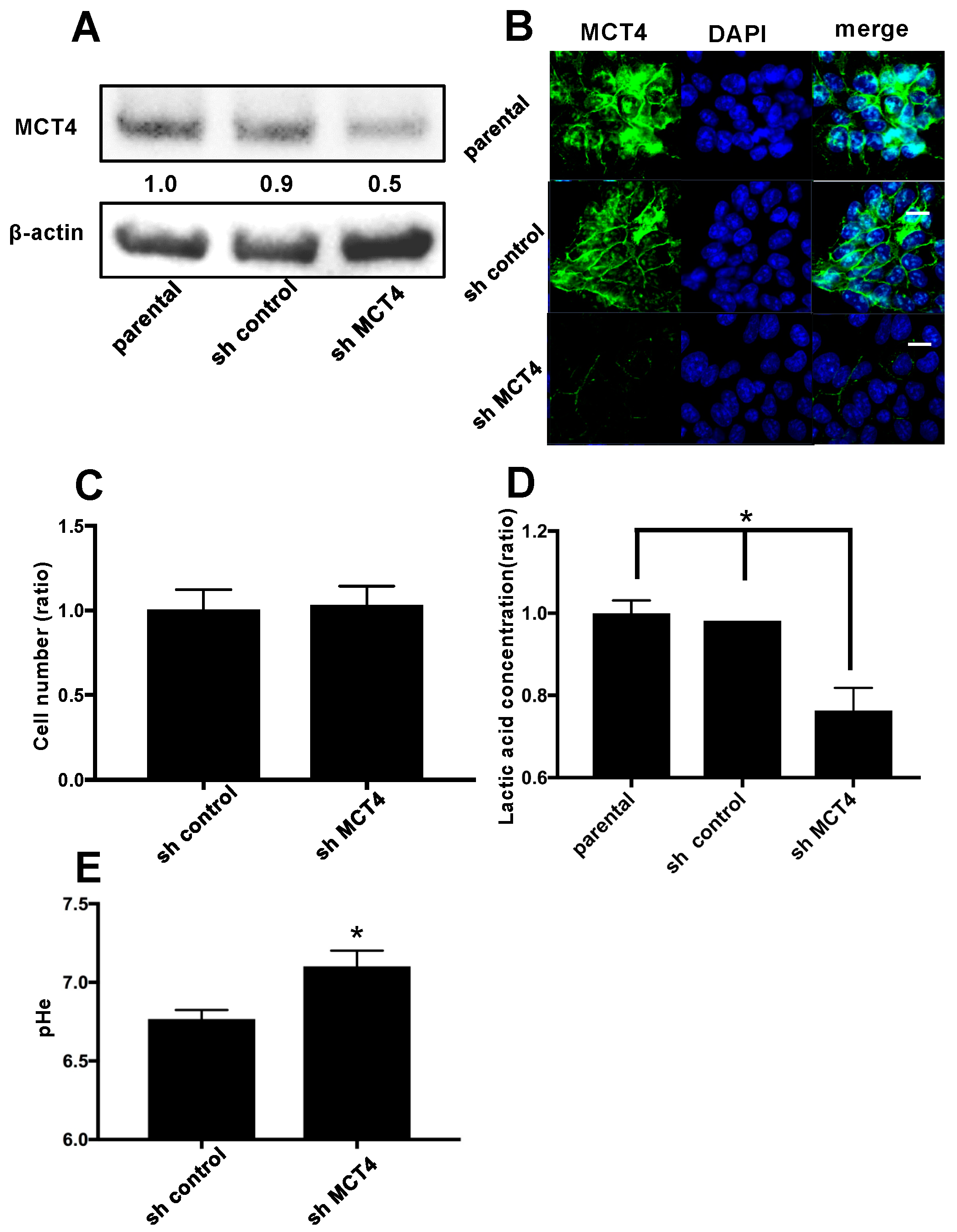
2.2. The Reduction of MCT4 Resulted in Decreased Lactic Acid Release
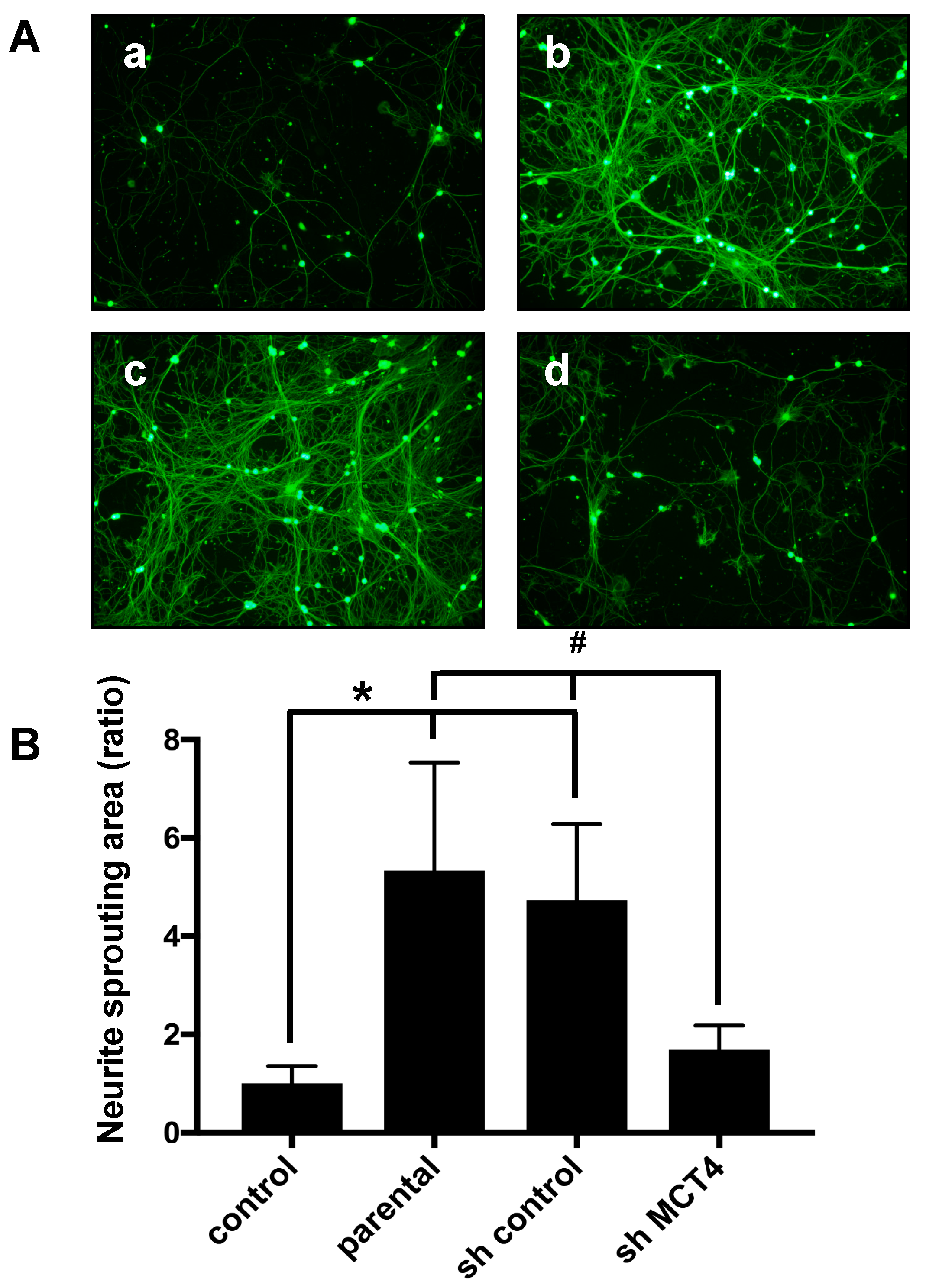
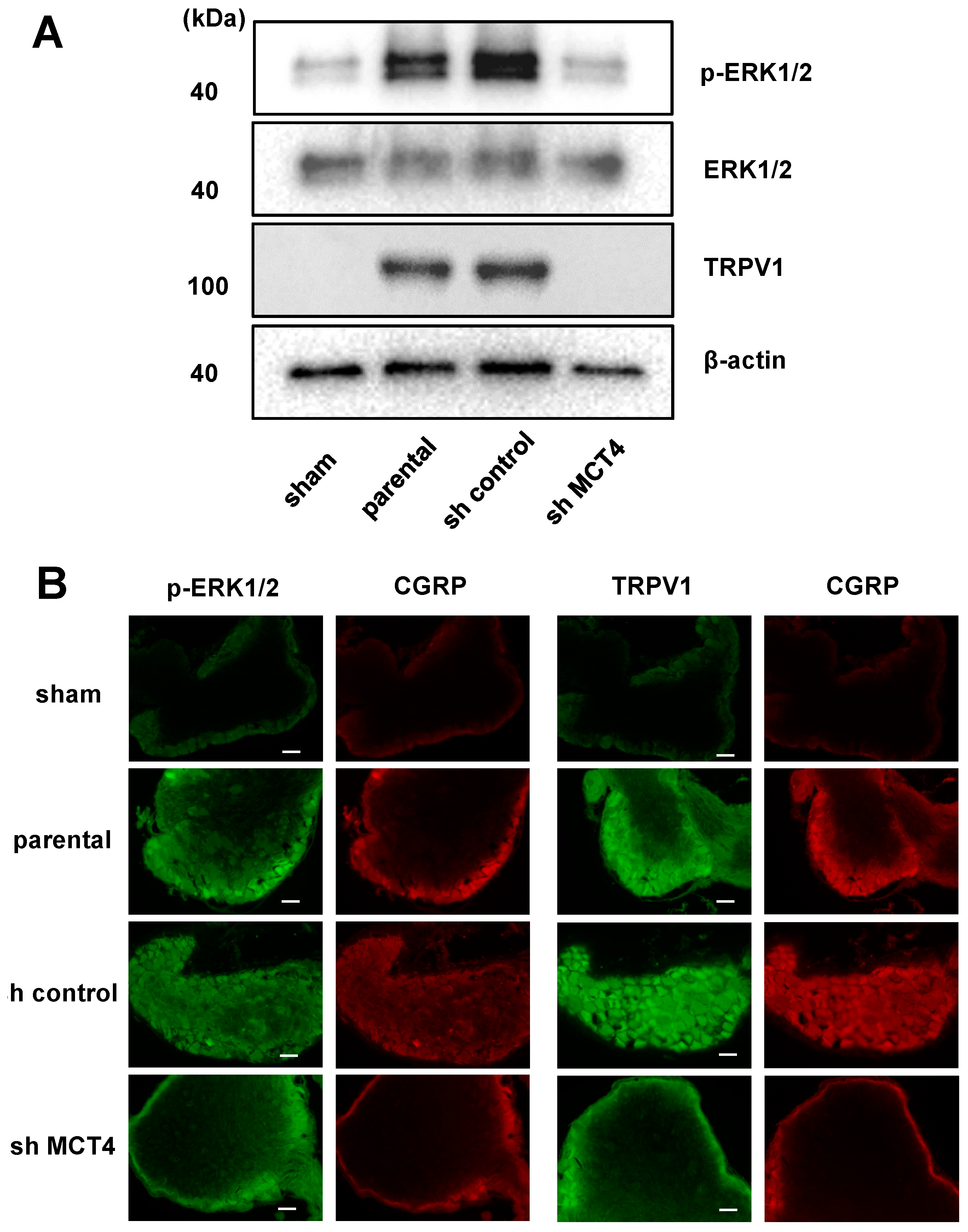
2.3. The Reduction of MCT4 in SAS Cells Decreased Sensory Nerve Sprouting
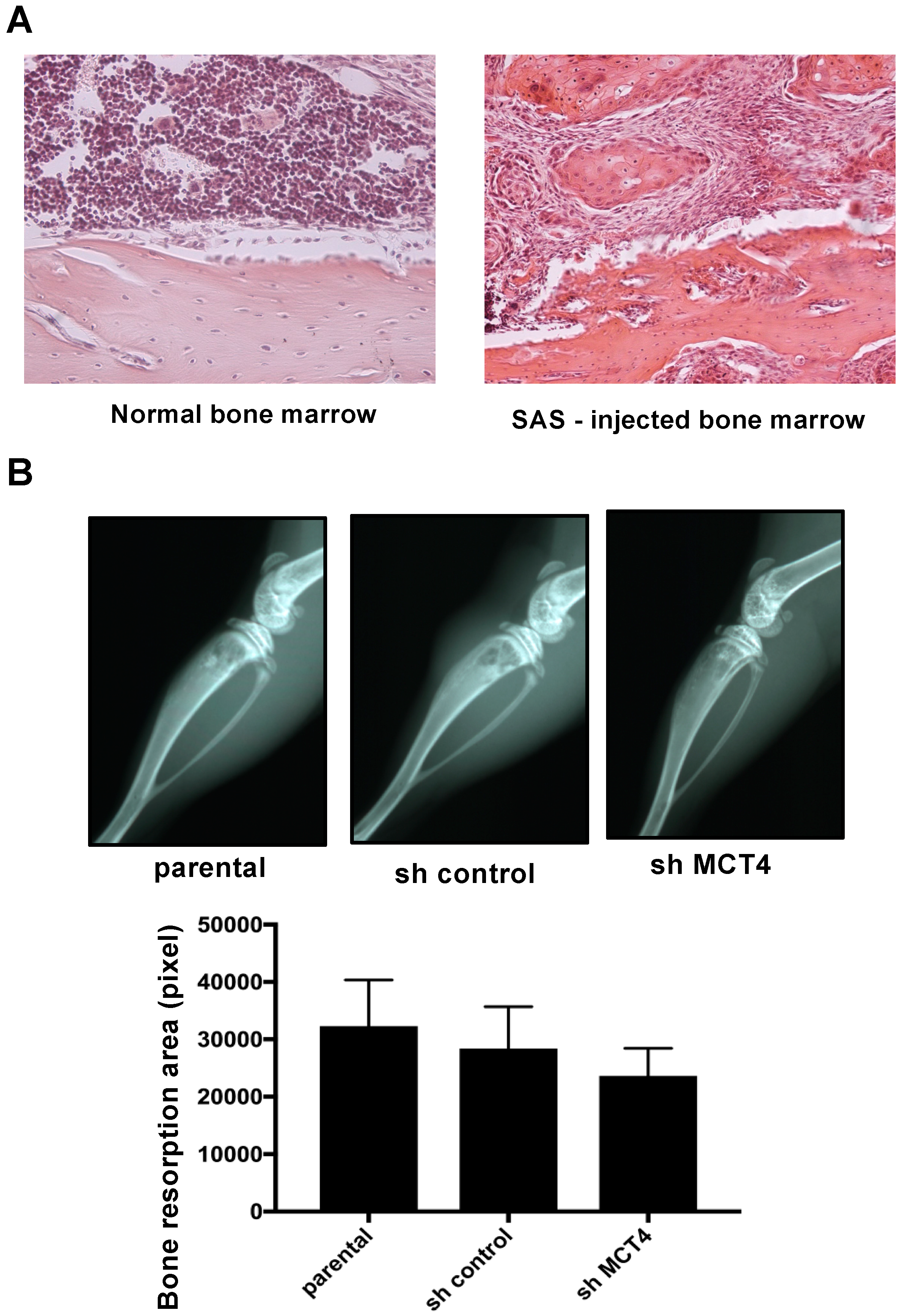
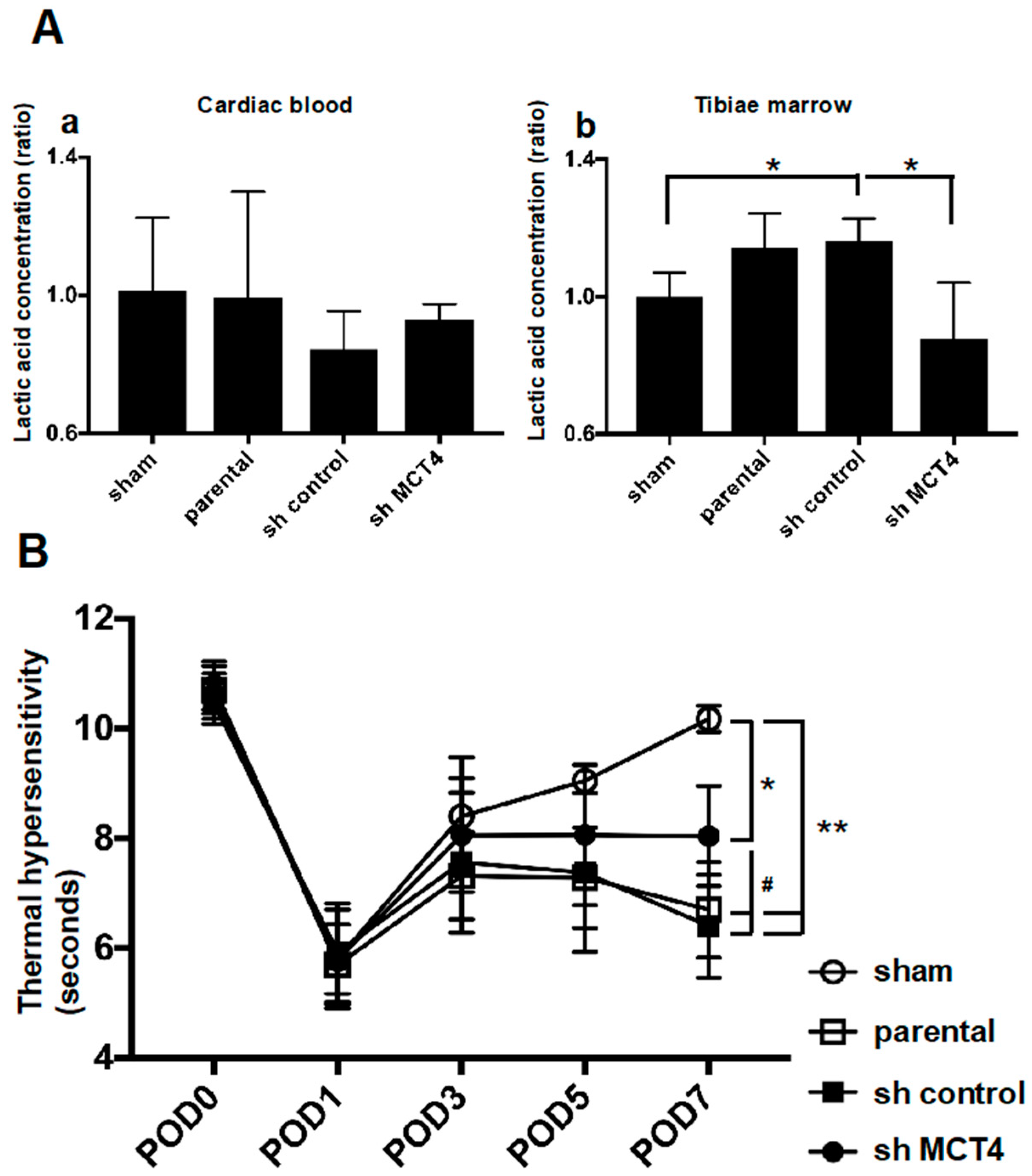
2.4. MCT4 Expression and HNSCC-BP Were Associated with SAS-Colonized bone
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Culture Conditions
4.3. Immunohistochemical Analysis
4.4. Analysis of MCT4 Expression in SAS Cells
4.5. Cell Proliferation Assay
4.6. Western Blot Analysis
4.7. DRG Fiber Sprouting Assay
4.8. Animal Experiments
4.9. In Vivo Analysis of HNSCC-BP
4.10. In Vivo Radiography and Measurement of Osteolytic Lesion Areas
4.11. Lactate Concentration Measurement
4.12. DRG Processing
4.13. Immunofluorescence Analysis
4.14. Statistical Analyses
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Brown, J.S.; Lowe, D.; Kalavrezos, N.; D’Souza, J.; Magennis, P.; Woolgar, J. Patterns of invasion and routes of tumor entry into the mandible by oral squamous cell carcinoma. Head Neck 2002, 24, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.F.; Rau, K.M.; Shao, Y.Y.; Yen, C.J.; Wu, M.F.; Chen, J.S.; Chang, C.S.; Yeh, S.P.; Chiou, T.J.; Hsieh, R.K.; et al. Patients with head and neck cancer may need more intensive pain management to maintain daily functioning: A multi-center study. Support Care Cancer 2018. [Google Scholar] [CrossRef] [PubMed]
- Schmitd, L.B.; Beesley, L.J.; Russo, N.; Bellile, E.L.; Inglehart, R.C.; Liu, M.; Romanowicz, G.; Wolf, G.T.; Taylor, J.M.G.; D’Silva, N.J. Redefining Perineural Invasion: Integration of Biology With Clinical Outcome. Neoplasia 2018, 20, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Guise, T.A. Parathyroid hormone-related protein and bone metastases. Cancer 1997, 80, 1572–1580. [Google Scholar] [CrossRef]
- Morisawa, A.; Okui, T.; Shimo, T.; Ibaragi, S.; Okusha, Y.; Ono, M.; Nguyen, T.T.H.; Hassan, N.M.M.; Sasaki, A. Ammonium tetrathiomolybdate enhances the antitumor effects of cetuximab via the suppression of osteoclastogenesis in head and neck squamous carcinoma. Int. J. Oncol. 2018, 52, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Wakisaka, S.; Hiraga, T.; Hata, K.; Nishimura, R.; Tominaga, M.; Yoneda, T. Decreased sensory nerve excitation and bone pain associated with mouse Lewis lung cancer in TRPV1-deficient mice. J. Bone Miner. Metab. 2018, 36, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Noel, J.; Gasull, X.; Delaunay, A.; Alloui, A.; Friend, V.; Eschalier, A.; Lazdunski, M.; Lingueglia, E. Acid-sensing ion channels in postoperative pain. J. Neurosci. 2011, 31, 6059–6066. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Daido, S.; Sun-Wada, G.H.; Wada, Y.; Futai, M.; Nakanishi-Matsuim, M. Diversity of proton pumps in osteoclasts: V-ATPase with a3 and d2 isoforms is a major form in osteoclasts. Biochim. Biophys. Acta 2014, 1837, 744–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertega-Gomes, N.; Vizcaino, J.R.; Attig, J.; Jurmeister, S.; Lopes, C.; Baltazar, F. A lactate shuttle system between tumour and stromal cells is associated with poor prognosis in prostate cancer. BMC Cancer 2014, 14, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samudio, I.; Fiegl, M.; Andreeff, M. Mitochondrial uncoupling and the Warburg effect: Molecular basis for the reprogramming of cancer cell metabolism. Cancer Res. 2009, 69, 2163–2166. [Google Scholar] [CrossRef] [PubMed]
- Elf, S.E.; Chen, J. Targeting glucose metabolism in patients with cancer. Cancer 2014, 120, 774–780. [Google Scholar]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Lane, S.; Korsak, A.; Paton, J.F.; Gourine, A.V.; Kasparov, S.; Teschemacher, A.G. Lactate-mediated glia-neuronal signalling in the mammalian brain. Nat. Commun. 2014, 5, 3284. [Google Scholar] [CrossRef] [PubMed]
- Hiasa, M.; Okui, T.; Allette, Y.M.; Ripsch, M.S.; Sun-Wada, G.H.; Wakabayashi, H.; Roodman, G.D.; White, F.A.; Yoneda, T. Bone Pain Induced by Multiple Myeloma Is Reduced by Targeting V-ATPase and ASIC3. Cancer Res. 2017, 77, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, D.; Christian, H.C. The SLC16 monocaboxylate transporter family. Xenobiotica 2008, 38, 1072–1106. [Google Scholar] [CrossRef] [PubMed]
- Perez-Escuredo, J.; Van Hee, V.F.; Sboarina, M.; Falces, J.; Payen, V.L.; Pellerin, L.; Sonveaux, P. Monocarboxylate transporters in the brain and in cancer. Biochim. Biophys. Acta 2016, 1863, 2481–2497. [Google Scholar] [CrossRef] [PubMed]
- Meredith, D.; Bell, P.; McClure, B.; Wilkins, R. Functional and molecular characterisation of lactic acid transport in bovine articular chondrocytes. Cell. Physiol. Biochem. 2002, 12, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Bonen, A. The expression of lactate transporters (MCT1 and MCT4) in heart and muscle. Eur. J. Appl. Physiol. 2011, 86, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, I.; Chang, W.J.; Ahn, S.M.; Lim, S.H.; Kim, H.S.; Yoo, K.H.; Jung, K.S.; Song, H.N.; Cho, J.H.; et al. MCT4 as a potential therapeutic target for metastatic gastric cancer with peritoneal carcinomatosis. Oncotarget 2016, 7, 43492–43503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Zhang, X.; Ding, X.; Li, H.; Geng, M.; Xie, Z.; Wu, H.; Huang, M. Lactate dehydrogenase B is associated with the response to neoadjuvant chemotherapy in oral squamous cell carcinoma. PLoS ONE 2015, 10, e0125976. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, S.J.; Simpson, I.A. Developmental switch in brain nutrient transporter expression in the rat. American journal of physiology. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1127–E1134. [Google Scholar] [CrossRef] [PubMed]
- Marchiq, I.; Pouyssegur, J. Hypoxia, cancer metabolism and the therapeutic benefit of targeting lactate/H(+) symporters. J. Mol. Med. 2016, 94, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Suzuki, T.; Hosono, K.; Hayashi, I.; Hashiba, S.; Onuma, Y.; Amano, H.; Kurihara, Y.; Kurihara, H.; Okamoto, H.; et al. Neuronal system-dependent facilitation of tumor angiogenesis and tumor growth by calcitonin gene-related peptide. Proc. Natl. Acad. Sci. USA 2008, 105, 13550–13555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneda, T.; Hata, K.; Nakanishi, M.; Nagae, M.; Nagayama, T.; Wakabayashi, H.; Nishisho, T.; Sakurai, T.; Hiraga, T. Involvement of acidic microenvironment in the pathophysiology of cancer-associated bone pain. Bone 2011, 48, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Ramer, L.M.; Borisoff, J.F.; Ramer, M.S. Rho-kinase inhibition enhances axonal plasticity and attenuates cold hyperalgesia after dorsal rhizotomy. J. Neurosci. 2004, 24, 10796–10805. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, K.; Okui, T.; Shimo, T.; Ibaragi, S.; Kawai, H.; Ryumon, S.; Kishimoto, K.; Okusha, Y.; Monsur Hassan, N.M.; Sasaki, A. Lactate Transporter Monocarboxylate Transporter 4 Induces Bone Pain in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 3317. https://doi.org/10.3390/ijms19113317
Hasegawa K, Okui T, Shimo T, Ibaragi S, Kawai H, Ryumon S, Kishimoto K, Okusha Y, Monsur Hassan NM, Sasaki A. Lactate Transporter Monocarboxylate Transporter 4 Induces Bone Pain in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2018; 19(11):3317. https://doi.org/10.3390/ijms19113317
Chicago/Turabian StyleHasegawa, Kazuaki, Tatsuo Okui, Tsuyoshi Shimo, Soichiro Ibaragi, Hotaka Kawai, Shoji Ryumon, Koji Kishimoto, Yuka Okusha, Nur Mohammad Monsur Hassan, and Akira Sasaki. 2018. "Lactate Transporter Monocarboxylate Transporter 4 Induces Bone Pain in Head and Neck Squamous Cell Carcinoma" International Journal of Molecular Sciences 19, no. 11: 3317. https://doi.org/10.3390/ijms19113317
APA StyleHasegawa, K., Okui, T., Shimo, T., Ibaragi, S., Kawai, H., Ryumon, S., Kishimoto, K., Okusha, Y., Monsur Hassan, N. M., & Sasaki, A. (2018). Lactate Transporter Monocarboxylate Transporter 4 Induces Bone Pain in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences, 19(11), 3317. https://doi.org/10.3390/ijms19113317