Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi


Abstract
:1. Introduction
2. Results
2.1. Dexamethasone Suppresses Synergistic Induction of PDE4B Expression by Roflumilast and NTHi In Vitro and In Vivo
2.2. Dexamethasone Suppresses NTHi-Induced Inflammation In Vitro and Mouse Lung In Vivo
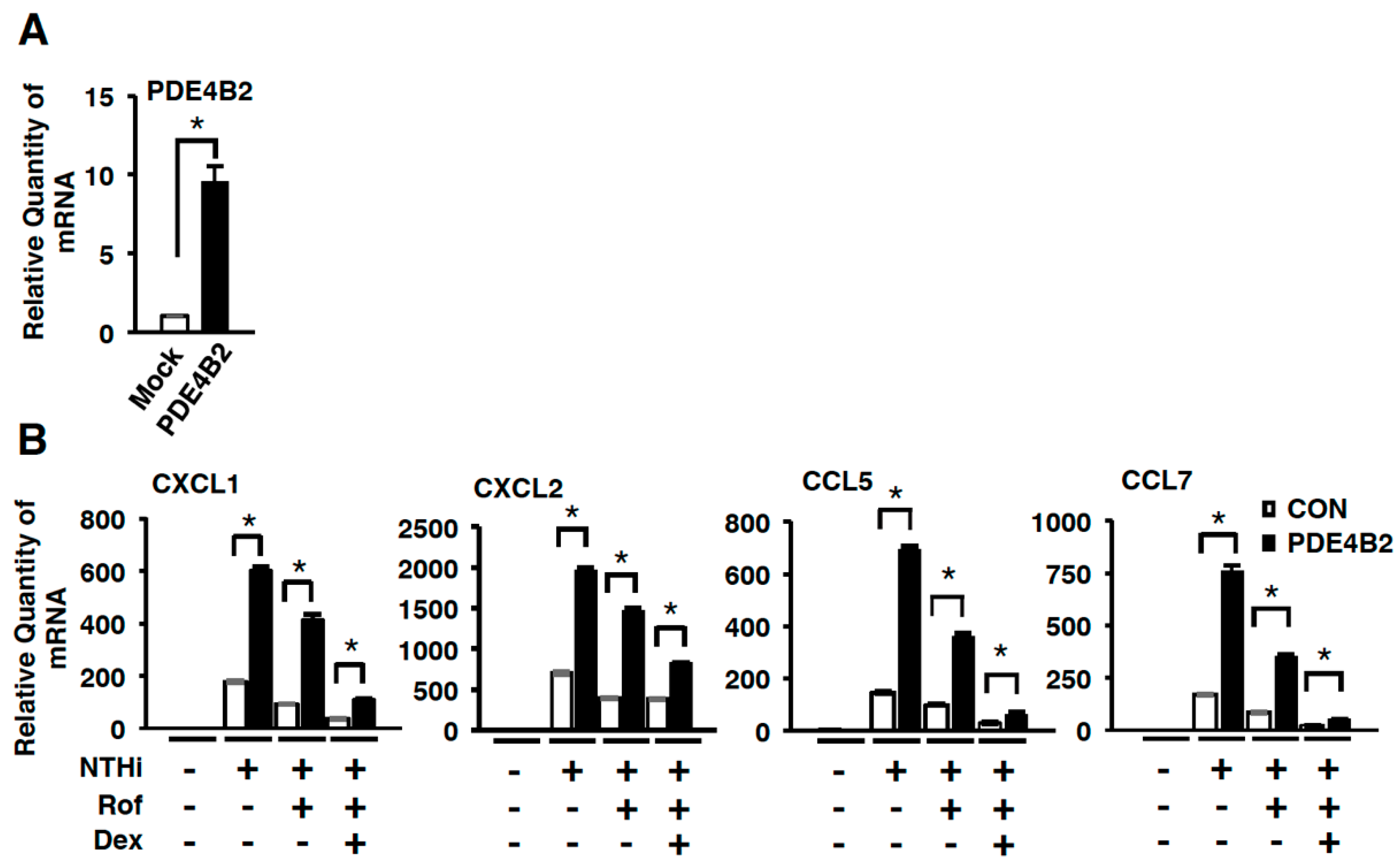
2.3. Dexamethasone Improves the Inhibitory Effect of Roflumilast on NTHi-Induced Inflammation by Suppressing Up-Regulated PDE4B
2.4. Dexamethasone Suppresses PDE4B Expression in a GR-Dependent Manner
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Bacterial Strains and Culture Condition
4.3. Cell Culture
4.4. Real-Time Quantitative and Semi-Quantitative RT-PCR Analyses
4.5. Plasmids and Transfections
4.6. Immunoblotting
4.7. Mice and Animal Experiments
4.8. Histology and Immunofluorescence Assay
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PDE4B | Phosphodiesterase 4B |
| NTHi | Nontypeable Haemophilus influenzae |
| GR | Glucocorticoid receptor |
| COPD | Chronic obstructive pulmonary disease |
| CF | Cystic fibrosis |
| GCs | Glucocorticoids |
| PMN | Polymorphonuclear neutrophils |
| BAL | Bronchoalveolar lavage |
References
- Ollivier, V.; Parry, G.C.; Cobb, R.R.; de Prost, D.; Mackman, N. Elevated cyclic AMP inhibits NF-kappaB-mediated transcription in human monocytic cells and endothelial cells. J. Biol. Chem. 1996, 271, 20828–20835. [Google Scholar] [CrossRef] [PubMed]
- Hertz, A.L.; Bender, A.T.; Smith, K.C.; Gilchrist, M.; Amieux, P.S.; Aderem, A.; Beavo, J.A. Elevated cyclic AMP and PDE4 inhibition induce chemokine expression in human monocyte-derived macrophages. Proc. Natl. Acad. Sci. USA 2009, 106, 21978–21983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-kappaB action. Cell. Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef] [PubMed]
- Oldenburger, A.; Roscioni, S.S.; Jansen, E.; Menzen, M.H.; Halayko, A.J.; Timens, W.; Meurs, H.; Maarsingh, H.; Schmidt, M. Anti-inflammatory role of the cAMP effectors Epac and PKA: Implications in chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e31574. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Annu. Rev. Biochem. 2007, 76, 481–511. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Mika, D.; Richter, W. Cyclic AMP compartments and signaling specificity: Role of cyclic nucleotide phosphodiesterases. J. Gen. Physiol. 2014, 143, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 58, 488–520. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.K.; Connett, J.; Bailey, W.C.; Casaburi, R.; Cooper, J.A., Jr.; Criner, G.J.; Curtis, J.L.; Dransfield, M.T.; Han, M.K.; Lazarus, S.C.; et al. Azithromycin for prevention of exacerbations of COPD. N. Engl. J. Med. 2011, 365, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.; Zlock, L.; Lao, A.; Mika, D.; Namkung, W.; Xie, M.; Scheitrum, C.; Gruenert, D.C.; Verkman, A.S.; Finkbeiner, W.E.; et al. Anchored PDE4 regulates chloride conductance in wild-type and DeltaF508-CFTR human airway epithelia. FASEB J. 2014, 28, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Lipworth, B.J. Phosphodiesterase-4 inhibitors for asthma and chronic obstructive pulmonary disease. Lancet 2005, 365, 167–175. [Google Scholar] [CrossRef]
- Milara, J.; Armengot, M.; Banuls, P.; Tenor, H.; Beume, R.; Artigues, E.; Cortijo, J. Roflumilast N-oxide, a PDE4 inhibitor, improves cilia motility and ciliated human bronchial epithelial cells compromised by cigarette smoke in vitro. Br. J. Pharmacol. 2012, 166, 2243–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milara, J.; Peiro, T.; Serrano, A.; Artigues, E.; Aparicio, J.; Tenor, H.; Sanz, C.; Cortijo, J. Simvastatin Increases the Ability of Roflumilast N-oxide to Inhibit Cigarette Smoke-Induced Epithelial to Mesenchymal Transition in Well-differentiated Human Bronchial Epithelial Cells in vitro. COPD 2015, 12, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Peiro, T.; Serrano, A.; Guijarro, R.; Zaragoza, C.; Tenor, H.; Cortijo, J. Roflumilast N-oxide inhibits bronchial epithelial to mesenchymal transition induced by cigarette smoke in smokers with COPD. Pulm. Pharmacol. Ther. 2014, 28, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Mehats, C.; Tanguy, G.; Dallot, E.; Robert, B.; Rebourcet, R.; Ferre, F.; Leroy, M.J. Selective up-regulation of phosphodiesterase-4 cyclic adenosine 3′,5′-monophosphate (cAMP)-specific phosphodiesterase variants by elevated cAMP content in human myometrial cells in culture. Endocrinology 1999, 140, 3228–3237. [Google Scholar] [CrossRef] [PubMed]
- Campos-Toimil, M.; Keravis, T.; Orallo, F.; Takeda, K.; Lugnier, C. Short-term or long-term treatments with a phosphodiesterase-4 (PDE4) inhibitor result in opposing agonist-induced Ca(2+) responses in endothelial cells. Br. J. Pharmacol. 2008, 154, 82–92. [Google Scholar] [CrossRef] [PubMed]
- D’Sa, C.; Tolbert, L.M.; Conti, M.; Duman, R.S. Regulation of cAMP-specific phosphodiesterases type 4B and 4D (PDE4) splice variants by cAMP signaling in primary cortical neurons. J. Neurochem. 2002, 81, 745–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallstrand, T.S.; Hackett, T.L.; Altemeier, W.A.; Matute-Bello, G.; Hansbro, P.M.; Knight, D.A. Airway epithelial regulation of pulmonary immune homeostasis and inflammation. Clin. Immunol. 2014, 151, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.M.; Golden, G.; Ikari, J.; Rennard, S.I. PDE4: A novel target in the treatment of chronic obstructive pulmonary disease. Clin. Pharmacol. Ther. 2012, 91, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Price, D.; Chisholm, A.; Ryan, D.; Crockett, A.; Jones, R. The use of roflumilast in COPD: A primary care perspective. Prim. Care Respir. J. 2010, 19, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Tashkin, D.P. Roflumilast: The new orally active, selective phophodiesterase-4 inhibitor, for the treatment of COPD. Expert Opin. Pharmacother. 2014, 15, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Grootendorst, D.C.; Gauw, S.A.; Verhoosel, R.M.; Sterk, P.J.; Hospers, J.J.; Bredenbroker, D.; Bethke, T.D.; Hiemstra, P.S.; Rabe, K.F. Reduction in sputum neutrophil and eosinophil numbers by the PDE4 inhibitor roflumilast in patients with COPD. Thorax 2007, 62, 1081–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannheimer, S.L.; Sorensen, E.A.; Haran, A.C.; Mansfield, C.N.; Wright, C.D.; Salmon, M. Additive anti-inflammatory effects of beta 2 adrenoceptor agonists or glucocorticosteroid with roflumilast in human peripheral blood mononuclear cells. Pulm. Pharmacol. Ther. 2012, 25, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Morell, A.; Ballester, B.; Sanz, C.; Freire, J.; Qian, X.; Alonso-Garcia, M.; Morcillo, E.; Cortijo, J. Roflumilast improves corticosteroid resistance COPD bronchial epithelial cells stimulated with toll like receptor 3 agonist. Respir. Res. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef] [PubMed]
- Rabe, K.F.; Bateman, E.D.; O’Donnell, D.; Witte, S.; Bredenbroker, D.; Bethke, T.D. Roflumilast--an oral anti-inflammatory treatment for chronic obstructive pulmonary disease: A randomised controlled trial. Lancet 2005, 366, 563–571. [Google Scholar] [CrossRef]
- Calverley, P.M.; Sanchez-Toril, F.; McIvor, A.; Teichmann, P.; Bredenbroeker, D.; Fabbri, L.M. Effect of 1-year treatment with roflumilast in severe chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2007, 176, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Vestbo, J.; Tan, L.; Atkinson, G.; Ward, J.; Uk, G.S.T. A controlled trial of 6-weeks’ treatment with a novel inhaled phosphodiesterase type-4 inhibitor in COPD. Eur. Respir. J. 2009, 33, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Susuki-Miyata, S.; Miyata, M.; Lee, B.C.; Xu, H.; Kai, H.; Yan, C.; Li, J.D. Cross-talk between PKA-Cbeta and p65 mediates synergistic induction of PDE4B by roflumilast and NTHi. Proc. Natl. Acad. Sci. USA 2015, 112, E1800–E1809. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Sargent, C.; Singh, D. Glucocorticoid sensitivity of lipopolysaccharide-stimulated chronic obstructive pulmonary disease alveolar macrophages. Clin. Exp. Immunol. 2009, 158, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.; Robinson, L.; Lea, S.; Banyard, A.; Blaikley, J.; Ray, D.; Bizzi, A.; Volpi, G.; Facchinetti, F.; Singh, D. Evaluation of glucocorticoid receptor function in COPD lung macrophages using beclomethasone-17-monopropionate. PLoS ONE 2013, 8, e64257. [Google Scholar] [CrossRef] [PubMed]
- Toward, T.J.; Broadley, K.J. Airway function, oedema, cell infiltration and nitric oxide generation in conscious ozone-exposed guinea-pigs: Effects of dexamethasone and rolipram. Br. J. Pharmacol. 2002, 136, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.L.; Milara, J.; Lluch, J.; De Diego, A.; Sanz, C.; Cortijo, J. Phosphodiesterase-4 inhibition improves corticosteroid insensitivity in pulmonary endothelial cells under oxidative stress. Allergy 2013, 68, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Luecke, H.F.; Yamamoto, K.R. The glucocorticoid receptor blocks P-TEFb recruitment by NFkappaB to effect promoter-specific transcriptional repression. Genes Dev. 2005, 19, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Busillo, J.M.; Cidlowski, J.A. The five Rs of glucocorticoid action during inflammation: Ready, reinforce, repress, resolve, and restore. Trends Endocrinol. Metab. 2013, 24, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Ahlstrom, M.; Pekkinen, M.; Huttunen, M.; Lamberg-Allardt, C. Dexamethasone down-regulates cAMP-phosphodiesterase in human osteosarcoma cells. Biochem. Pharmacol. 2005, 69, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; De Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell. Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Tillis, C.C.; Huang, H.W.; Bi, W.; Pan, S.; Bruce, S.R.; Alcorn, J.L. Glucocorticoid regulation of human pulmonary surfactant protein-B (SP-B) mRNA stability is independent of activated glucocorticoid receptor. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L940–L950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otulakowski, G.; Duan, W.; Sarangapani, A.; Gandhi, S.; O’Brodovich, H. Glucocorticoid-mediated repression of REDD1 mRNA expression in rat fetal distal lung epithelial cells. Pediatr. Res. 2009, 65, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, L.; Liu, B.; Blaxall, B.C.; Taubman, M.B. A novel role for the glucocorticoid receptor in the regulation of monocyte chemoattractant protein-1 mRNA stability. J. Biol. Chem. 2007, 282, 10146–10152. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Conti, M. Induction of the cyclic nucleotide phosphodiesterase PDE4B is essential for LPS-activated TNF-alpha responses. Proc. Natl. Acad. Sci. USA 2002, 99, 7628–7633. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Brooks, J.; Hagan, G.; Cahn, A.; O’Connor, B.J. Superiority of “triple” therapy with salmeterol/fluticasone propionate and tiotropium bromide versus individual components in moderate to severe COPD. Thorax 2008, 63, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Calverley, P.M.; Rabe, K.F.; Goehring, U.M.; Kristiansen, S.; Fabbri, L.M.; Martinez, F.J. M2-124 and M2-125 study groups. Roflumilast in symptomatic chronic obstructive pulmonary disease: Two randomised clinical trials. Lancet 2009, 374, 685–694. [Google Scholar] [CrossRef]
- Donnelly, L.E.; Barnes, P.J. Chemokine receptors as therapeutic targets in chronic obstructive pulmonary disease. Trends Pharmacol. Sci. 2006, 27, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.; Zhou, Y.; Iribarren, P.; Wang, J. Chemokines and chemokine receptors: Their manifold roles in homeostasis and disease. Cell. Mol. Immunol. 2004, 1, 95–104. [Google Scholar] [PubMed]
- Quint, J.K.; Wedzicha, J.A. The neutrophil in chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2007, 119, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Howarth, P.H. A comparison of the anti-inflammatory properties of intranasal corticosteroids and antihistamines in allergic rhinitis. Allergy 2000, 55 (Suppl. 62), 6–11. [Google Scholar] [CrossRef] [PubMed]
- Hidi, R.; Timmermans, S.; Liu, E.; Schudt, C.; Dent, G.; Holgate, S.T.; Djukanovic, R. Phosphodiesterase and cyclic adenosine monophosphate-dependent inhibition of T-lymphocyte chemotaxis. Eur. Respir. J. 2000, 15, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, S.; Plumb, J.; Kaur, M.; Ray, D.; Singh, D. Additive anti-inflammatory effects of corticosteroids and phosphodiesterase-4 inhibitors in COPD CD8 cells. Respir. Res. 2016, 17, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F.J.; Calverley, P.M.; Goehring, U.M.; Brose, M.; Fabbri, L.M.; Rabe, K.F. Effect of roflumilast on exacerbations in patients with severe chronic obstructive pulmonary disease uncontrolled by combination therapy (REACT): A multicentre randomised controlled trial. Lancet 2015, 385, 857–866. [Google Scholar] [CrossRef]
- Kanehiro, A.; Ikemura, T.; Makela, M.J.; Lahn, M.; Joetham, A.; Dakhama, A.; Gelfand, E.W. Inhibition of phosphodiesterase 4 attenuates airway hyperresponsiveness and airway inflammation in a model of secondary allergen challenge. Am. J. Respir. Crit. Care Med. 2001, 163, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.G.; Deng, Y.M.; Wu, X.; Tang, H.F.; Deng, J.F.; Chen, J.Q.; Yang, S.Y.; Xie, Q.M. Inhibition of phosphodiesterase activity, airway inflammation and hyperresponsiveness by PDE4 inhibitor and glucocorticoid in a murine model of allergic asthma. Life Sci. 2006, 79, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Guess, A.; Agrawal, S.; Wei, C.C.; Ransom, R.F.; Benndorf, R.; Smoyer, W.E. Dose- and time-dependent glucocorticoid receptor signaling in podocytes. Am. J. Physiol. Renal. Physiol. 2010, 299, F845–F853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almawi, W.Y.; Melemedjian, O.K. Negative regulation of nuclear factor-kappaB activation and function by glucocorticoids. J. Mol. Endocrinol. 2002, 28, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, J.J.; Land, S.C.; Tarnow-Mordi, W.O.; Zembala, M.; Kowalczyk, D.; Lauterbach, R. Immunopharmacological potential of selective phosphodiesterase inhibition. I. Differential regulation of lipopolysaccharide-mediated proinflammatory cytokine (interleukin-6 and tumor necrosis factor-alpha) biosynthesis in alveolar epithelial cells. J. Pharmacol. Exp. Ther. 2002, 300, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.A.; Taverna, J.; Chaves, J.; Makkinje, A.; Lerner, A. Phosphodiesterase 4 inhibitors augment levels of glucocorticoid receptor in B cell chronic lymphocytic leukemia but not in normal circulating hematopoietic cells. Clin. Cancer Res. 2007, 13, 4920–4927. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Dong, H.; Kim, E.J.; Weintraub, L.; Epstein, P.M.; Lerner, A. Type 4 cAMP phosphodiesterase (PDE4) inhibitors augment glucocorticoid-mediated apoptosis in B cell chronic lymphocytic leukemia (B-CLL) in the absence of exogenous adenylyl cyclase stimulation. Biochem. Pharmacol. 2005, 69, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Barenkamp, S.J.; Leininger, E. Cloning, expression, and DNA sequence analysis of genes encoding nontypeable Haemophilus influenzae high-molecular-weight surface-exposed proteins related to filamentous hemagglutinin of Bordetella pertussis. Infect. Immun. 1992, 60, 1302–1313. [Google Scholar] [PubMed]
- Lee, B.C.; Miyata, M.; Lim, J.H.; Li, J.D. Deubiquitinase CYLD acts as a negative regulator for bacterium NTHi-induced inflammation by suppressing K63-linked ubiquitination of MyD88. Proc. Natl. Acad. Sci. USA 2016, 113, E165–E171. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Lee, J.Y.; Susuki-Miyata, S.; Wang, W.Y.; Xu, H.; Kai, H.; Kobayashi, K.S.; Flavell, R.A.; Li, J.D. Glucocorticoids suppress inflammation via the upregulation of negative regulator IRAK-M. Nat. Commun. 2015, 6, 6062. [Google Scholar] [CrossRef] [PubMed]
- Horvat, J.C.; Beagley, K.W.; Wade, M.A.; Preston, J.A.; Hansbro, N.G.; Hickey, D.K.; Kaiko, G.E.; Gibson, P.G.; Foster, P.S.; Hansbro, P.M. Neonatal chlamydial infection induces mixed T-cell responses that drive allergic airway disease. Am. J. Respir. Crit. Care Med. 2007, 176, 556–564. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| human PDE4B | CTATACCGATCGCATTCAGGTC | CTGTCCATTGCCGATACAATT |
| human PDE4B2 | AGCGGTGGTAGCGGTGACTC | GCAGCGTGCAGGCTGTTGTG |
| human CXCL1 | TGCAGGGAATTCACCCCAAG | AGCTTTCCGCCCATTCTTGA |
| human CXCL2 | GTGTGAAGGTGAAGTCCCCC | AGCTTTCTGCCCATTCTTGA |
| human CCL5 | CTACACCAGTGGCAAGTGC | CTTTCGGGTGACAAAGACGAC |
| human CCL7 | GGCTTGCTCAGCCAGTTG | GGTGGTCCTTCTGTAGCTCTC |
| human Cyclophillin A | CGGGTCCTGGCATCTTGT | GCAGATGAAAAACTGGGAACCA |
| mouse pde4b2 | GTAGAGGCCAGTTCCCATCA | CCAACACCTAGTGCAGAGC |
| mouse cxcl1 | CATGGCTGGGATTCACCTCA | CCTCGCGACCATTCTTGAGT |
| mouse cxcl2 | TCAATGCCTGAAGACCCTG | GCAAACTTTTTGACCGCCCT |
| mouse ccl5 | CCTCACCATATGGCTCGGAC | ACGACTGCAAGATTGGAGCA |
| mouse ccl7 | CAATGCATCCACATGCTGCT | GCAGACTTCCATGCCCTTCT |
| mouse gapdh | ACCCAGAAGACTGTGGATGG | GGATGCAGGGATGATGTTCT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.-C.; Susuki-Miyata, S.; Yan, C.; Li, J.-D. Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. Int. J. Mol. Sci. 2018, 19, 3511. https://doi.org/10.3390/ijms19113511
Lee B-C, Susuki-Miyata S, Yan C, Li J-D. Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. International Journal of Molecular Sciences. 2018; 19(11):3511. https://doi.org/10.3390/ijms19113511
Chicago/Turabian StyleLee, Byung-Cheol, Seiko Susuki-Miyata, Chen Yan, and Jian-Dong Li. 2018. "Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi" International Journal of Molecular Sciences 19, no. 11: 3511. https://doi.org/10.3390/ijms19113511
APA StyleLee, B. -C., Susuki-Miyata, S., Yan, C., & Li, J. -D. (2018). Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. International Journal of Molecular Sciences, 19(11), 3511. https://doi.org/10.3390/ijms19113511