Regenerative Models for the Integration and Regeneration of Head Skeletal Tissues


Abstract
:1. Introduction
2. Current Regenerative Therapies for the Jaw
3. The Patterning and Development of the Cranial-Facial Skeleton
4. Differentiation of the Head Skeleton
5. Vertebrate Model Systems to Study for Jaw Regeneration
6. The Anatomy of the Lower Jaw in Regenerating Species
7. Endogenous Regeneration of the Jaw
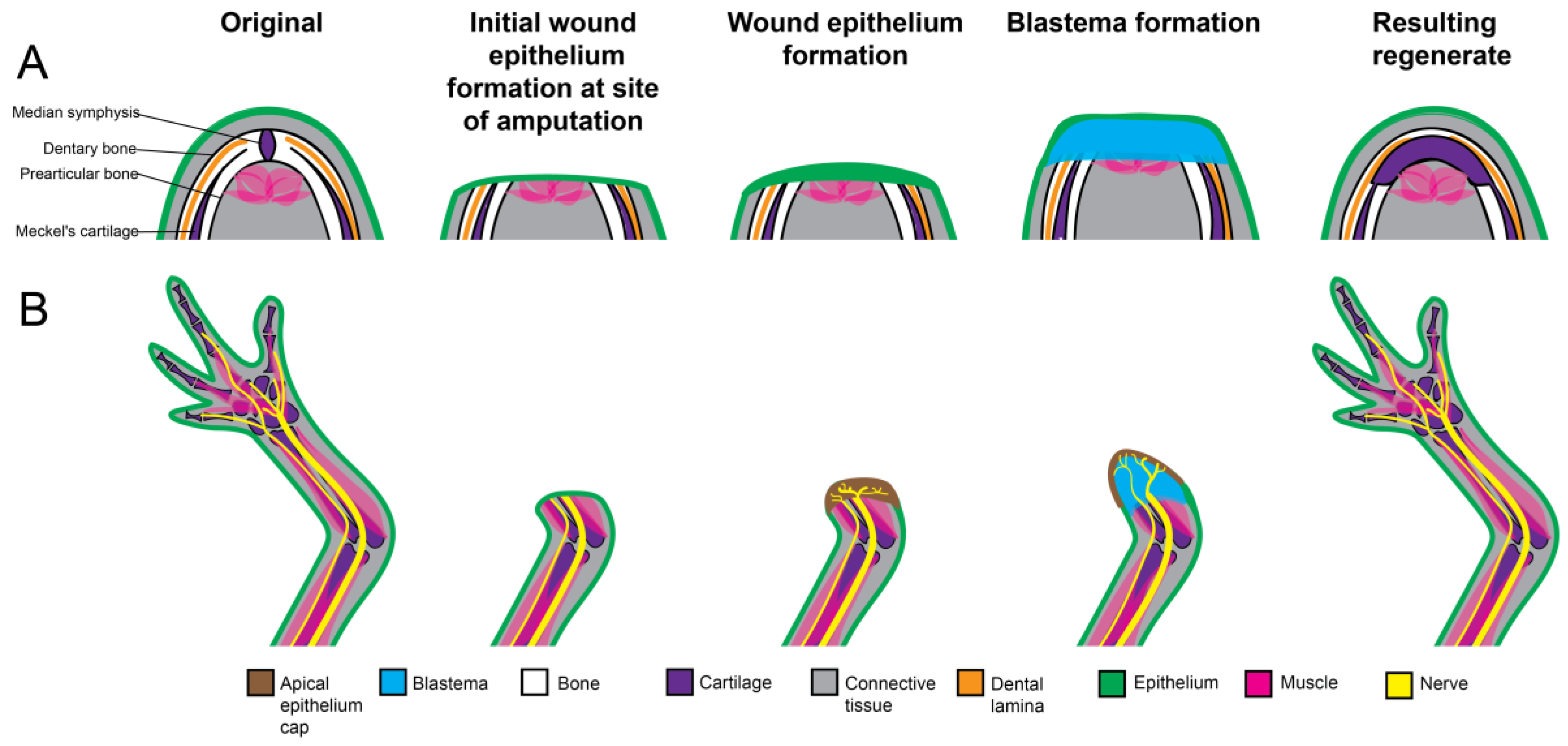
8. Jaw Blastema Formation
9. Jaw Blastema Cell Differentiation
9.1. Skeleton Formation
9.2. Teeth Restoration
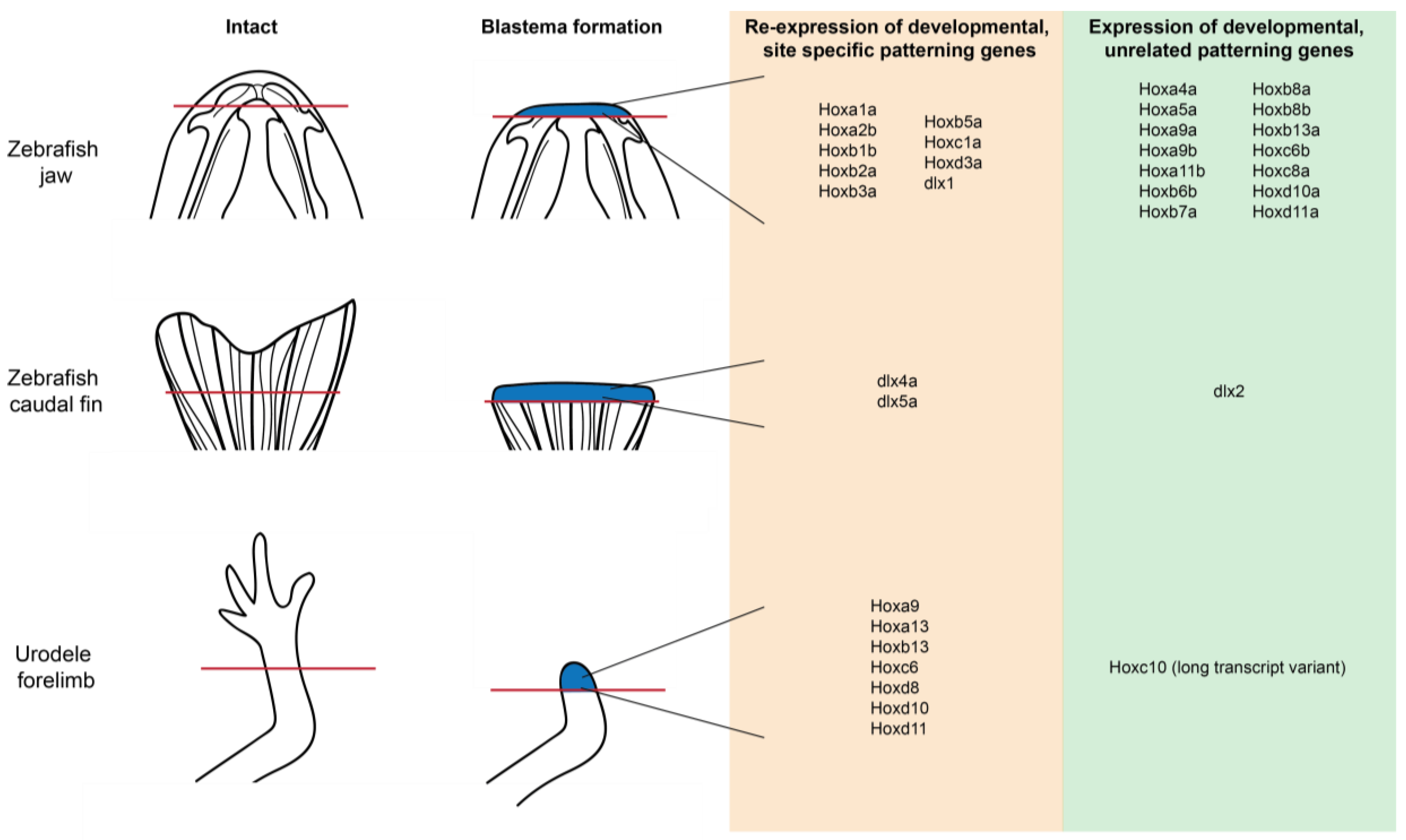
10. Pattern Formation and Positional Identity
Hox Gene Re-Expression and Positional Plasticity
11. Mechanisms of Jaw Fracture Healing
12. Tissue Integration
13. On the Development of New Regenerative Therapies: Conserved Mechanisms of Endogenous Regeneration
13.1. Positional Plasticity
13.2. Imperfect Fracture Repair
13.3. Age-Related Changes in Regenerative Capacity
13.4. Transplantations
14. Conclusions
Funding
Conflicts of Interest
References
- Cawood, J.I.; Howell, R.A. A classification of the edentulous jaws. Int. J. Oral Maxillofac. Surg. 1988, 17, 232–236. [Google Scholar] [CrossRef]
- McCarthy, J.G. The role of distraction osteogenesis in the reconstruction of the mandible in unilateral craniofacial microsomia. Clin. Plast. Surg. 1994, 21, 625–631. [Google Scholar] [PubMed]
- Shwyrkow, M.B.; Shamsudinov, A.K. Methods of simultaneous treatment of the mandible defects and the adjacent soft tissues. Acta Chir. Plast. 1989, 31, 226–235. [Google Scholar] [PubMed]
- Komisar, A. Mandibular Reconstruction; Thieme: Stuttgart, Germany, 1997; ISBN 313103811X. [Google Scholar]
- Glenske, K.; Donkiewicz, P.; Kowitsch, A.; Milosevic-Oljaca, N.; Rider, P.; Rofall, S.; Franke, J.; Jung, O.; Smeets, R.; Schnettler, R.; Wenisch S, S.; Barbeck, M. Applications of Metals for Bone Regeneration. Int. J. Mol. Sci. 2018, 19, 826. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Xie, J.; Liao, J.; Lin, S.; Lin, Y. Nanomaterials and bone regeneration. Bone Res. 2015, 3, 15029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.E.; Choi, J.; Joung, Y.K.; Park, K.; Han, D.K. Controlled release of bone morphogenetic protein (BMP)-2 from nanocomplex incorporated on hydroxyapatite-formed titanium surface. J. Control. Release 2012, 160, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, J.; Liu, J.; Wei, L.; Wu, G. BMP-functionalised coatings to promote osteogenesis for orthopaedic implants. Int. J. Mol. Sci. 2014, 15, 10150–10168. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Yague, M.A.; Abbah, S.A.; McNamara, L.; Zeugolis, D.I.; Pandit, A.; Biggs, M.J. Biomimetic approaches in bone tissue engineering: Integrating biological and physicomechanical strategies. Adv. Drug Deliv. Rev. 2015, 84, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Faruq, O.; Kim, B.; Padalhin, A.R.; Lee, G.H.; Lee, B.-T. A hybrid composite system of biphasic calcium phosphate granules loaded with hyaluronic acid-gelatin hydrogel for bone regeneration. J. Biomater. Appl. 2017, 32, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Florencio-Silva, R.; Sasso, G.R.D.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci. 2018, 19, 329. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.; Unger, R.; Sousa, R.; Reis, R. Crosstalk between osteoblasts and endothelial cells co-cultured on a polycaprolactone–starch scaffold and the in vitro development of vascularization. Biomaterials 2009, 30, 4407–4415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, D.A. Fibula free flap mandible reconstruction. Microsurgery 1994, 15, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Sanger, J.R.; Matloub, H.S.; Yousif, N.J. Sequential connection of flaps: A logical approach to customized mandibular reconstruction. Am. J. Surg. 1990, 160, 402–404. [Google Scholar] [CrossRef]
- Tang, Y.B.; Hahn, L.J. Major mandibular reconstruction with vascularized bone graft. J. Formos. Med. Assoc. 1990, 89, 34–40. [Google Scholar] [PubMed]
- Nkenke, E.; Neukam, F.W. Autogenous bone harvesting and grafting in advanced jaw resorption: Morbidity, resorption and implant survival. Eur. J. Oral Implantol. 2014, 7, S203–S217. [Google Scholar] [PubMed]
- Sakkas, A.; Wilde, F.; Heufelder, M.; Winter, K.; Schramm, A. Autogenous bone grafts in oral implantology—Is it still a “gold standard?” A consecutive review of 279 patients with 456 clinical procedures. Int. J. Implant Dent. 2017, 2, 23. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Kim, H.M.; Byun, J.H.; Kim, U.K.; Sung, I.Y.; Cho, Y.C.; Park, B.W. Stability of simultaneously placed dental implants with autologous bone grafts harvested from the iliac crest or intraoral jaw bone. BMC Oral Health 2015, 15, 172. [Google Scholar] [CrossRef] [PubMed]
- Mertens, C.; Decker, C.; Seeberger, R.; Hoffman, J.; Sander, A.; Freier, K. Early bone resorption after vertical bone augmentation—A comparison of calvarial and iliac grafts. Clin. Oral Implants Res. 2012, 24, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, K.A.; Schultze-Mosgau, S.; Wiltfang, J.; Neikam, F.W.; Rupprecht, S.; Thorwarth, M. Changes of mineralization of free autogenous bone grafts used for sinus floor elevation. Clin. Oral Implants Res. 2006, 17, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Akintoye, S.O.; Lam, T.; Shi, S.; Brahim, J.; Collins, M.T.; Robey, P.G. Skeletal site-specific characterization of orofacial and iliac crest human bone marrow stromal cells in same individuals. Bone 2006, 38, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Ge, J.; Zhang, P.; Fu, Y.; Zhang, Z.; Cheng, J.; Jiang, H. Phenotypic characterization of craniofacial bone marrow stromal cells: Unique properties of enhanced osteogenesis, cell recruitment, autophagy, and apoptosis resistance. Cell Tissue Res. 2014, 358, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, P.; Duvina, M.; Barbato, L.; Biondi, E.; Nuti, N.; Brancato, L.; Delle Rose, G. Bone regeneration in dentistry. Clin. Cases Miner. Bone Metab. 2011, 8, 24–28. [Google Scholar] [PubMed]
- Khojasteh, A.; Kheiri, L.; Motamedian, S.; Khoshkam, V. Guided bone regeneration for the reconstruction of alveolar bone defects. Ann. Maxillofac. Surg. 2017, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Castellon, L.; Jerez, D.; Mayorga, J.; Fuenzalida, C. Remodeling of Costochondral Graft After Mandibular Reconstruction. J. Oral Maxillofac. Surg. 2017, 75, 226.e1–226.e7. [Google Scholar] [CrossRef] [PubMed]
- Almaiman, M.; Al-Bargi, H.; Manson, P. Complication of Anterior Iliac Bone Graft Harvesting in 372 Adult Patients from May 2006 to May 2011 and a Literature Review. Cranial Maxillofac. Trauma Reconstruct. 2013, 6, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosoglu, M. Enterocutaneous Fistula: A Complication of Posterior Iliac Bone Graft Harvesting not Previously Described. Acta Neurochir. 1998, 140, 1089–1092. [Google Scholar] [CrossRef] [PubMed]
- Younger, E.; Chapman, M. Morbidity at bone graft donor sites. J. Orthop. Trauma 1989, 3, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, Y. Regenerative Medicine for Jawbone. JMAJ 2004, 47, 294–297. [Google Scholar]
- Manimaran, K.; Sharma, R.; Sankaranarayanan, S.; Perumal, S.M. Regeneration of mandibular ameloblastema defect with the help of autologous dental pulp stem cells and buccal pad of fat stromal vascular fraction. Ann. Maxillofac. Surg. 2016, 6, 97–100. [Google Scholar] [PubMed]
- Knight, R.D.; Schilling, T.F. Cranial neural crest and development of the head skeleton. Adv. Exp. Med. Biol. 2006, 589, 120–133. [Google Scholar] [PubMed]
- Depew, M.J.; Lufkin, T.; Rubenstein, J.L.R. Specification of jaw subdivisions by Dlx genes. Science 2002, 298, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Li, X.; McEvilly, R.J.; Rosenfeld, M.G.; Lufkin, T.; Rubenstein, J.L. Dlx genes pattern mammalian jaw primordium by regulating both lower jaw-specific and upper jaw-specific genetic programs. Development 2008, 135, 2905–2916. [Google Scholar] [CrossRef] [PubMed]
- Minoux, M.; Rijli, F.M. Molecular mechanisms of cranial neural crest cell migration and patterning in craniofacial development. Development 2010, 137, 2605–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijli, F.M.; Mark, M.; Lakkaraju, S.; Dierich, A.; Dolle, P.; Chambron, P. A Homeotic Transformation Is Generated in the Rostra1 Branchial Region of the Head by Disruption of Hoxa-2, Which Acts as a Selector Gene. Cell 1993, 75, 1333–1349. [Google Scholar] [CrossRef]
- Gendron-Maguire, M.; Mallo, M.; Zhang, M.; Gridley, T. Hoxa-2 mutant mice exhibit homeotic transformation of skeletal elements derived from cranial neural crest. Cell 1993, 75, 1317–1331. [Google Scholar] [CrossRef]
- Pasqualetti, M.; Ori, M.; Nardi, I.; Rijli, F.M. Ectopic Hoxa2 induction after neural crest migration results in homeosis of jaw elements in Xenopus. Development 2000, 127, 5367–5378. [Google Scholar] [PubMed]
- Hunter, M.P.; Prince, V.E. Zebrafish Hox Paralogue Group 2 Genes Function Redundantly as Selector Genes to Pattern the Second Pharyngeal Arch. Dev. Biol. 2002, 247, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Noden, D.M. The role of the neural crest in patterning of avian cranial skeletal, connective, and muscle tissues. Dev. Biol. 1983, 96, 144–165. [Google Scholar] [CrossRef]
- Schilling, T.F.; Prince, V.; Ingham, P.W. Plasticity in Zebrafish hox Expression in the Hindbrain and Cranial Neural Crest. Dev. Biol. 2001, 231, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Trainor, P.A.; Krumlauf, R. Patterning the cranial neural crest: Hindbrain segmentation and Hox gene plasticity. Nat. Rev. Neurosci. 2000, 1, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.A.; Helms, J.A. The cellular and molecular origins of beak morphology. Science 2003, 299, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Couly, G.; Creuzet, S.; Bennaceur, S.; Vincent, C.; le Douarin, N.M. Interactions between Hox-negative cephalic neural crest cells and the foregut endoderm in patterning the facial skeleton in the vertebrate head. Development 2002, 129, 1061–1073. [Google Scholar] [PubMed]
- David, N.B.; Saint-Etienne, L.; Tsang, M.; Schilling, T.F.; Rosa, F.M. Requirement for endoderm and FGF3 in ventral head skeleton formation. Development 2002, 129, 4457–4468. [Google Scholar] [PubMed]
- Creuzet, S.; Couly, G.; Vincent, C.; le Douarin, N.M. Negative effect of Hox gene expression on the development of the neural crest-derived facial skeleton. Development 2002, 129, 4301–4313. [Google Scholar] [PubMed]
- Abzhanov, A.; Tzahor, E.; Lassar, A.B.; Tabin, C.J. Dissimilar regulation of cell differentiation in mesencephalic (cranial) and sacral (trunk) neural crest cells in vitro. Development 2003, 130, 4567–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGonnell, I.M.; Graham, A. Trunk Neural Crest Has Skeletogenic Potential. Curr. Biol. 2002, 12, 767–771. [Google Scholar] [CrossRef]
- Opperman, L.A. Cranial sutures as intramembranous bone growth sites. Dev. Dyn. 2000, 219, 472–485. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.T.; Schilling, T.F.; Lee, K.; Parker, J.; Kimmel, C.B. Sucker encodes a zebrafish Endothelin-1 required for ventral pharyngeal arch development. Development 2000, 127, 3815–3828. [Google Scholar] [PubMed]
- Kimmel, C.B. Endothelin 1-mediated regulation of pharyngeal bone development in zebrafish. Development 2003, 130, 1339–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.T. Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint. Development 2003, 130, 1353–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouthier, D.E.; Hosoda, K.; Richardson, J.A.; Williams, S.C.; Yanagisawa, H.; Kuwaki, T.; Kumada, M.; Hammer, R.E.; Yanagisawa, M. Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development 1998, 125, 813–824. [Google Scholar] [PubMed]
- Kurihara, Y.; Kurihara, H.; Suzuki, H.; Kodama, T.; Maemura, K.; Nagai, R.; Oda, H.; Kuwaki, T.; Cao, W.H.; Kamada, N.; et al. Elevated blood pressure and craniofaclal abnormalities in mice deficient in endothelin-1. Nature 1994, 368, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.A.; Tucker, A.S.; Sharpe, P.T. Temporospatial cell interactions regulating mandibular and maxillary arch patterning. Development 2000, 127, 403–412. [Google Scholar] [PubMed]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The zebrafish as a model for complex tissue regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, F.; Li, B.; Stocum, D.L. Amphibians as research models for regenerative medicine. Organogenesis 2010, 6, 1–10. [Google Scholar] [CrossRef]
- Tazaki, A.; Tanaka, E.M.; Fei, J.-F. Developmental Biology. Dev. Biol. 2017, 432, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kurosaka, H.; Takano-Yamamoto, T.; Yamashiro, T.; Agata, K. Comparison of molecular and cellular events during lower jaw regeneration of newt (Cynops pyrrhogaster) and West African clawed frog (Xenopus tropicalis). Dev. Dyn. 2007, 237, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Brazaitis, P. Maxillary Regeneration in a Marsh Crocodile, Crocodylus palustris. J. Herpetol. 1981, 15, 360. [Google Scholar] [CrossRef]
- Ghosh, S.; Thorogood, P.; Ferretti, P. Regenerative capability of upper and lower jaws in the newt. Int. J. Dev. Biol. 1994, 38, 479–490. [Google Scholar] [PubMed]
- Wang, X.; He, H.; Tang, W.; Zhang, X.A.; Hua, X.; Yan, J. Two Origins of Blastemal Progenitors Define Blastemal Regeneration of Zebrafish Lower Jaw. PLoS ONE 2012, 7, e45380. [Google Scholar] [CrossRef] [PubMed]
- Finch, R.A. The influence of the nerve on lower jaw regeneration in the adult newt, Triturus viridescens. J. Morphol. 1969, 129, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.J.; Stagg, M.W. Regeneration in lower jaws of newts after excision of the intermandibular regions. J. Exp. Zool. 1958, 137, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, M.; Foxe, J.J.; Pedrosa, E.; Hrabovsky, A.; Carroll, R.; Zheng, D.; and Lachman, H.M. Transcriptome Comparison of Human Neurons Generated Using Induced Pluripotent Stem Cells Derived from Dental Pulp and Skin Fibroblasts. PLoS ONE 2013, 8, e75682. [Google Scholar] [CrossRef] [PubMed]
- Tank, P.W. The Effect of Nonlimb Tissues on Forelimb Regeneration in the Axolotl, Ambystoma-Mexicanum. J. Exp. Zool. 1987, 244, 409–423. [Google Scholar] [CrossRef]
- Kim, K.; Zhao, R.; Doi, A.; Ng, K.; Unternaehrer, J.; Cahan, P.; Huo, H.; Loh, Y.H.; Aryee, M.J.; Lensche, M.W.; et al. Donor cell type can influence the epigenome and differentiation potential of human induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 1117–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goss, R.J.; Stagg, M.W. Regeneration of lower jaws in adult newts. J. Morphol. 1958, 102, 289–309. [Google Scholar] [CrossRef]
- Yntema, C.L. Regeneration in sparsely innervated and aneurogenic forelimbs of Amblystoma larvae. J. Exp. Zool. 1959, 140, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, Y.S.; Oh, H.S.; Yang, K.H.; Kim, E.C.; Chi, J.G. Prenatal Development of the Human Mandible. Anat. Rec. 2001, 263, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Vazques, J.F.; Merida-Velasco, J.R.; Merida-Velasco, J.A.; Sanchez-Montesinos, I.; Espin-Ferra, J.; Jimenez-Collado, J. Development of Meckel’s Cartilage in the Symphyseal Region in Man. Dev. Biol. 1997, 249, 249–254. [Google Scholar] [CrossRef]
- Hovorakova, M.; Lesot, H.; Peterka, M.; Peterkova, R. Early development of the human dentition revisited. J. Anat. 2018, 233, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Lismaa, S.E.; Kaidonis, X.; Nicks, A.M.; Bogush, N.; Kikuchi, K.; Naqvi, N.; Harvey, R.P.; Husain, A.; Graham, R.M. Comparative regenerative mechanisms across different mammalian tissues. npj Regen. Med. 2018, 3, 6. [Google Scholar]
- Graver, H.T. Origin of the dental lamina in the regenerating salamander jaw. J. Exp. Zool. 1974, 189, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Thorogood, P.; Ferretti, P. Regeneration of lower and upper jaws in urodeles is differentially affected by retinoic acid. Int. J. Dev. Biol. 1996, 40, 1161–1170. [Google Scholar] [PubMed]
- Dwaraka, V.B.; Smith, J.J.; Woodcock, M.R.; Voss, S.R. Comparative transcriptomics of limb regeneration_ Identification of conserved expression changes among three species of Ambystoma. Genomics 2018. [Google Scholar] [CrossRef]
- McCusker, C.; Bryant, S.V.; Gardiner, D.M. The axolotl limb blastema: Cellular and molecular mechanisms driving blastema formation and limb regeneration in tetrapods. Regeneration 2015, 2, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Singer, M. On the Nature of the Neurotrophic Phenomenon in Urodele Limb Regeneration. Integr. Comp. Biol. 1978, 18, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Mullen, L.M.; Bryant, S.V.; Torok, M.A.; Blumberg, B.; Gardiner, D.M. Nerve dependency of regeneration: The role of Distal-less and FGF signaling in amphibian limb regeneration. Development 1996, 122, 3487–3497. [Google Scholar] [PubMed]
- McCusker, C.D.; Athippozhy, A.; Diaz-Castillo, C.; Fowlkes, C.; Gardiner, D.M.; Voss, S.R. Positional plasticity in regenerating Amybstoma mexicanum limbs is associated with cell proliferation and pathways of cellular differentiation. BMC Dev. Biol. 2015, 15, 45. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.R.; Wolpert, L. Nerves and angiogenesis in amphibian limb regeneration. Nature 1975, 257, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Voss, S.R.; Palumbo, A.; Nagarajan, R.; Gardiner, D.M.; Muneoka, K.; Stromberg, A.J.; Athippozhy, A.T. Gene expression during the first 28 days of axolotl limb regeneration I: Experimental design and global analysis of gene expression. Regeneration 2015, 2, 120–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, X.; Lyu, K.; Gai, G.; Wang, G.; Fan, C.; Zhang, X.A.; Yan, J. Time Point-Based Integrative Analyses of Deep-Transcriptome Identify Four Signal Pathways in Blastemal Regeneration of Zebrafish Lower Jaw. Stem Cells 2015, 33, 806–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kragl, M.; Knapp, D.; Nacu, E.; Khattak, S.; Maden, M.; Epperlein, H.H.; Tanaka, E.M. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature 2009, 460, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Kragl, M.; Roensch, K.; Nusslein, I.; Tazaki, A.; Taniguchi, Y.; Tarui, H.; Hayashi, T.; Agata, K.; Tanaka, E.M. Muscle and connective tissue progenitor populations show distinct Twist1 and Twist3 expression profiles during axolotl limb regeneration. Dev. Biol. 2013, 373, 196–204. [Google Scholar] [CrossRef] [PubMed]
- McCusker, C.D.; Diaz-Castillo, C.; Sosnik, J.; Phan, A.Q.; Gardiner, D.M. Cartilage and bone cells do not participate in skeletal regeneration in Ambystoma mexicanum limbs. Dev. Biol. 2016, 416, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Muneoka, K.; Fox, W.F.; Bryant, S.V. Cellular contribution from dermis and cartilage to the regenerating limb blastema in axolotls. Dev. Biol. 1986, 116, 256–260. [Google Scholar] [CrossRef]
- Nacu, E.; Glausch, M.; Le, H.Q.; Damanik, F.F.R.; Schuez, M.; Knapp, D.; Khattak, S.; Richter, T.; Tanaka, E.M. Connective tissue cells, but not muscle cells, are involved in establishing the proximo-distal outcome of limb regeneration in the axolotl. Dev. Stem Cells 2013, 140. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Schindler, S.; Giovannone, D.; de Millo Terrazzani, A.; Mariani, F.V.; Crump, J.G. Ihha induces hybrid cartilage-bone cells during zebrafish jawbone regeneration. Development 2016, 143, 2066–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, C.S. The histogenesis of the regenerating fore limb of larval Amblystoma after exarticulation of the humerus. J. Morphol. 1938, 62, 219–241. [Google Scholar] [CrossRef]
- Charbonneau, A.M.; Roy, S.; Tran, S.D. Oral-Facial Tissue Reconstruction in the Regenerative Axolotl. J. Exp. Zool. (Mol. Dev. Evol.) 2017, 326, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Padhi, B.K.; Joly, L.; Tellis, P.; Sith, A.; Nanjappa, P.; Chevrete, M.; Ekker, M.; Akimenko, M.A. Screen for genes differentially expressed during regeneration of the zebrafish caudal fin. Dev. Dyn. 2004, 231, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaghan, J.R.; Athippozhy, A.; Seifert, A.W.; Putta, S.; Stromberg, A.J.; Maden, M.; Gardiner, D.M.; Voss, S.R. Gene expression patterns specific to the regenerating limb of the Mexican axolotl. Biol. Open 2012, 1, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.; Tsun, Z.-Y.; Belmonte, J.C.I. A histone demethylase is necessary for regeneration in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 19889–19894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinn, J.L.; Bondre, C.; Gladstone, H.B.; Brown, P.O.; Chang, H.Y. Anatomic Demarcation by Positional Variation in Fibroblast Gene Expression Programs. PLoS Genet. 2006, 2, e119. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Mallo, M.S. Reassessing the Role of Hox Genes during Vertebrate Development and Evolution. Trends Genet. 2018, 34, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Chi, J.T.; Dudoit, S.; Bondre, C.; van de Rijn, M.; Botstein, D.; Brown, P.O. Diversity, topographic differentiation, and positional memory in human fibroblasts. Proc. Natl. Acad. Sci. USA 2002, 99, 12877–12882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, T.; Bryant, S.V.; Gardiner, D.M. A stepwise model system for limb regeneration. Dev. Biol. 2004, 270, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Wang, J.K.; Allen, N.; Brugmann, S.A.; Mikels, A.J.; Liu, H.; Ridky, T.W.; Stadler, H.S.; Nusse, R.; Helms, J.A.; et al. A dermal HOX transcriptional program regulates site-specific epidermal fate. Genes Dev. 2008, 22, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhouailly, D.; Rogers, G.E.; Sengel, P. The specification of feather and scale protein synthesis in epidermal-dermal recombinations. Dev. Biol. 1978, 65, 58–68. [Google Scholar] [CrossRef]
- Phan, A.Q.; Lee, J.; Oei, M.; Flath, C.; Hwe, C.; Mariano, R.; Vu, T.; Shu, C.; Dinh, A.; Simkin, J.; et al. Positional information in axolotl and mouse limb extracellular matrix is mediated via heparan sulfate and fibroblast growth factor during limb regeneration in the axolotl (Ambystoma mexicanum). Regeneration 2015, 2, 182–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCusker, C.D.; Gardiner, D.M. Positional Information Is Reprogrammed in Blastema Cells of the Regenerating Limb of the Axolotl (Ambystoma mexicanum). PLoS ONE 2013, 8, e77064. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.G. Regeneration of the urodele forelimb after reversal of its proximo-distal axis. J. Morphol. 1955, 96, 265–281. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Blumberg, B.; Komine, Y.; Bryant, S.V. Regulation of HoxA expression in developing and regenerating axolotl limbs. Development 1995, 121, 1731–1741. [Google Scholar] [PubMed]
- Carlson, M.R.; Komine, Y.; Bryant, S.V.; Gardiner, D.M. Expression of Hoxb13 and Hoxc10 in developing and regenerating Axolotl limbs and tails. Dev. Biol. 2001, 229, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Khan, P.A.; Tsilfidis, C.; Liversage, R.A. Hox C6 expression during development and regeneration of forelimbs in larval Notophthalmus viridescens. Dev. Genes Evol. 1999, 209, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.S.; Robitaille, A.m.; Wang, Y.; Ray, C.A.; Thummel, R.; Gu, H.; Djukovic, D.; Raftery, D.; Berndt, J.D.; Moon, R.T. Transcriptomic, proteomic, and metabolomic landscape of positional memory in the caudal fin of zebrafish. Proc. Natl. Acad. Sci. USA 2017, 114, E717–E726. [Google Scholar] [CrossRef] [PubMed]
- Leucht, P.; Kim, J.B.; Amasha, R.; James, A.W.; Girod, S.; Helms, J.A. Embryonic origin and Hox status determine progenitor cell fate during adult bone regeneration. Development 2008, 135, 2845–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Lee, J.; Yun, H.-S.; Shin, H.-I.; Park, E.K. Comparison of bone regeneration rate in flat and long bone defects: Calvarial and tibial bone. Tissue Eng. Regen. Med. 2013, 10, 336–340. [Google Scholar] [CrossRef]
- Pritchard, J.J. Repair of fractures of the parietal bone in rats. J. Anat. 1946, 80, 1–8. [Google Scholar]
- Hall, B.K.; Jacobson, H.N. The repair of fractured membrane bones in the newly hatched chick. Anat. Rec. 1975, 181, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.K.; Jacobson, H.N. Repair of Fractured Lower Jaws in the Spotted Salamander: Do Amphibians Form Secondary Cartilage? J. Exp. Zool. 1985. [Google Scholar] [CrossRef]
- Moss, M.L. Studies of the acellular bone of teleost fish. II. Response to fracture under normal an acalcemic conditions. Cells Tissues Organs 1962, 48, 46–60. [Google Scholar] [CrossRef]
- Gardiner, D.M. Regenerative Engineering and Developmental Biology; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- McCusker, C.; Lehrberg, J.; Gardiner, D. Position-specific induction of ectopic limbs in non-regenerating blastemas on axolotl forelimbs. Regeneration 2014, 1, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hay, E.D. The fine structure of differentiating muscle in the salamander tail. Cell Tissue Res. 1963, 59, 6–34. [Google Scholar] [CrossRef]
- Sosnik, J.; Vieira, W.A.; Webster, K.A.; Siegfried, K.R.; McCusker, C.D. A new and improved algorithm for the quantification of chromatin condensation from microscopic data shows decreased chromatin condensation in regenerating axolotl limb cells. PLoS ONE 2017, 12, e0185292. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, W.; Zang, T.; Zou, Y.; Fang, S.; Smith, D.K.; Bachoo, R.; and Zhang, C.L. In vivo reprogramming of astrocytes to neuroblasts in the adult brain. Nat. Cell Biol. 2013, 15, 1164–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torper, O.; Pfisterer, U.; Wolf, D.A.; Pereira, M.; Lau, S.; Jakobsson, J.; Bjorklund, A.; Grealish, S.; Parmar, M. Generation of induced neurons via direct conversion in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 7038–7043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, A.; Sumiyoshi, K.; Lopez-Juarez, A.; Howard, J.; Sakthivel, B.; Aronow, B.; Campell, K.; Nakafuku, M. Environmental impact on direct neuronal reprogramming in vivo in the adult brain. Nat. Commun. 2013, 4, 2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rais, Y.; Zviran, A.; Geula, S.; Gafni, O.; Chomsky, E.; Viukov, S.; Alfatah, A.; Caspi, I.; Krupalnik, V.; Zerbid, M.; et al. Deterministic direct reprogramming of somatic cells to pluripotency. Nature 2013, 502, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.; Mosteiro, L.; Pantoja, C.; Canamero, M.; Rayon, T.; Ors, I.; Grana, O.; Megias, D.; Dominguez, O.; Martinez, D.; et al. Reprogramming in vivo produces teratomas and iPS cells with totipotency features. Nature 2013, 502, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Stadtfeld, M.; Maherali, N.; Borkent, M.; Hochedlinger, K. A reprogrammable mouse strain from gene-targeted embryonic stem cells. Nat. Methods 2010, 7, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Kurita, M.; Araoka, T.; Hishida, T.; O’Keefe, D.D.; Takahashi, Y.; Sakamoto, A.; Sakurai, M.; Suzuki, K.; Wu, J.; Yamamoto, M.; et al. In vivo reprogramming of wound-resident cells generates skin epithelial tissue. Nature 2018, 561, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.R.; Graves, S.E.; Bain, G.I. Synthetic bone graft substitutes. ANZ J. Surg. 2001, 71, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Neovius, E.; Engstrand, T. Craniofacial reconstruction with bone and biomaterials: Review over the last 11 years. Br. J. Plast. Surg. 2010, 63, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.T.; Rosenbaum, A.J. Bone grafts, bone substitutes and orthobiologics. Organogenesis 2014, 8, 114–124. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Mammalian | Avian | Urodele | Teleost | |
|---|---|---|---|---|---|
| Regeneration | Capacity | No | No | Yes | Yes |
| Mechanism | n/a | n/a | Epimorphic regeneration | Epimorphic regeneration | |
| Pattern relative to original | n/a | n/a | Juvenile = identical (based on w/m), Adult = different (based on w/m, µCT) | Different (based on w/m) | |
| Integration with stump | n/a | n/a | Juvenile = fully integrated (based on w/m) Adult = incomplete (based on w/m, hst) | Incomplete (based on w/m, hst) | |
| Fracture healing | Capacity | Yes | Yes | Yes | Yes |
| Mechanism | Secondary ossification | Secondary ossification | Absence of secondary ossification | Absence of secondary ossification | |
| Pattern relative to original | Poor (based on hst, µCT) | Poor (based on hst) | Poor (based on hst) | Poor (based on hst) | |
| Integration with stump | Poor (based on hst, µCT) | Poor (based on hst) | Poor (based on hst) | Poor (based on hst) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, W.A.; McCusker, C.D. Regenerative Models for the Integration and Regeneration of Head Skeletal Tissues. Int. J. Mol. Sci. 2018, 19, 3752. https://doi.org/10.3390/ijms19123752
Vieira WA, McCusker CD. Regenerative Models for the Integration and Regeneration of Head Skeletal Tissues. International Journal of Molecular Sciences. 2018; 19(12):3752. https://doi.org/10.3390/ijms19123752
Chicago/Turabian StyleVieira, Warren A., and Catherine D. McCusker. 2018. "Regenerative Models for the Integration and Regeneration of Head Skeletal Tissues" International Journal of Molecular Sciences 19, no. 12: 3752. https://doi.org/10.3390/ijms19123752
APA StyleVieira, W. A., & McCusker, C. D. (2018). Regenerative Models for the Integration and Regeneration of Head Skeletal Tissues. International Journal of Molecular Sciences, 19(12), 3752. https://doi.org/10.3390/ijms19123752