α-Melanocyte-Stimulating Hormone Attenuates Neovascularization by Inducing Nitric Oxide Deficiency via MC-Rs/PKA/NF-κB Signaling


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
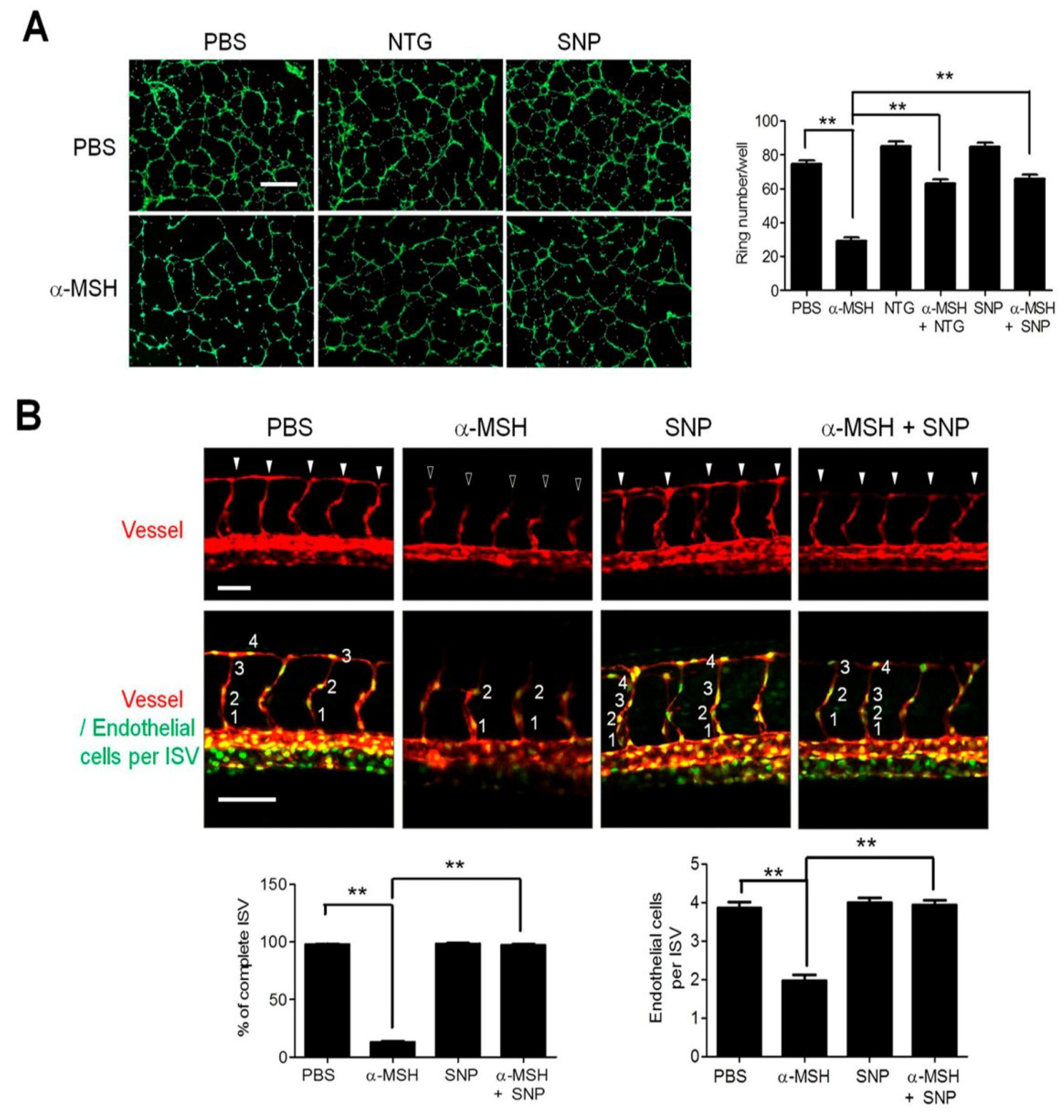
2.1. Nitric Oxide (NO) Deficiency Participated in α-Melanocyte-Stimulating Hormone (α-MSH)-Induced Angiogenesis Inhibition
2.2. l-Arginine (l-Arg) Supply Mitigated the α-MSH-Induced Neovascularization Blockade in Zebrafish
2.3. NO/Soluble Guanylyl Cyclase (sGC) Signaling Modulates α-MSH-Induced Angiogenesis Inhibition
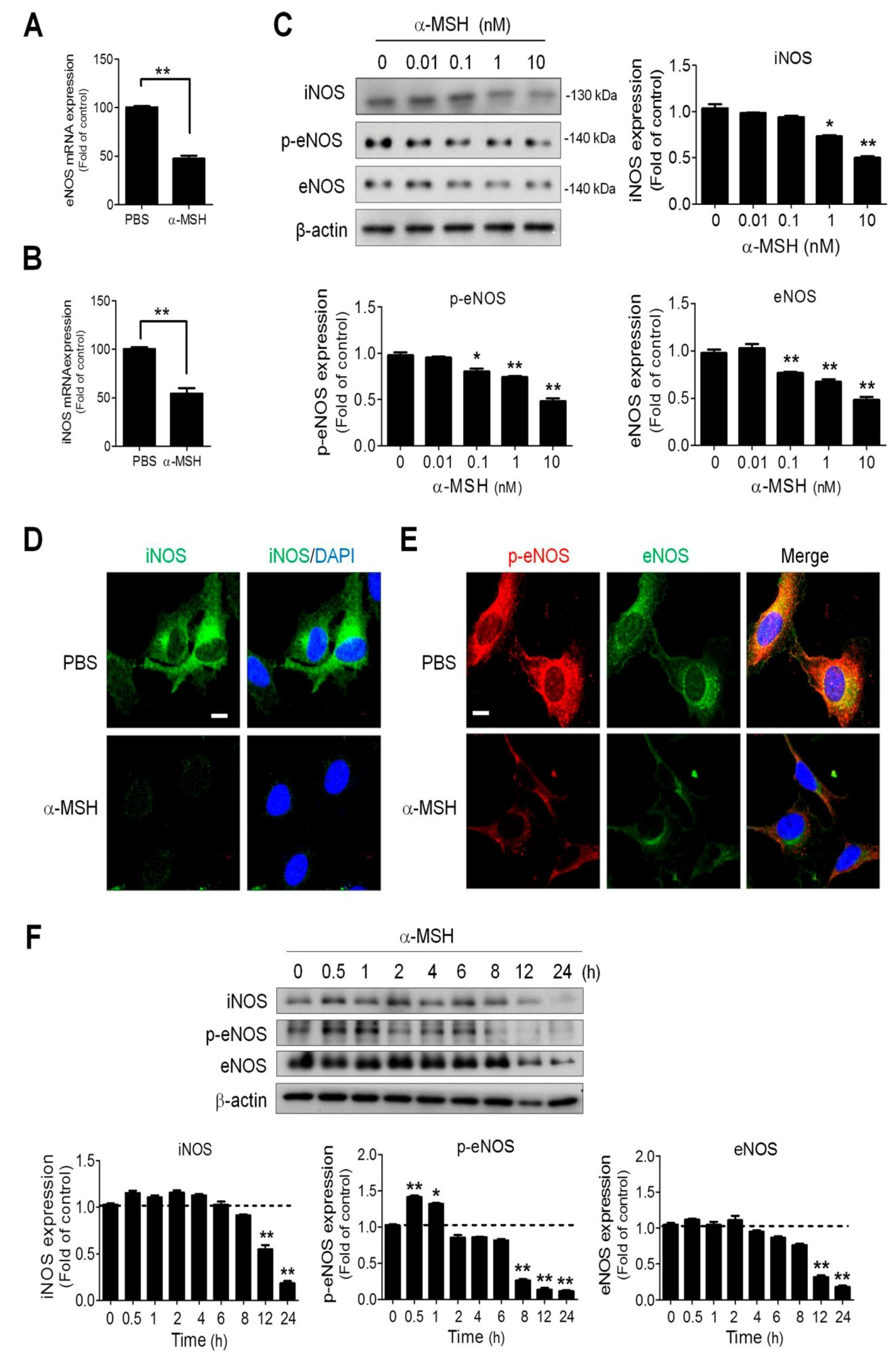
2.4. α-MSH Reduced Endothelial Endothelial and Inducible Nitric Oxide Synthase (eNOS and iNOS) Expression in a Dose-Dependent Manner
2.5. α-MSH Inhibited the Basal NF-κB Activities in Endothelial Cells by Elevating IκB and Reducing p65/p50/p105 Expression
2.6. MC1-R and MC2-R Contributed to the α-MSH-Induced Reduction of eNOS and iNOS Signaling
2.7. Pharmaceutical Protein Kinase A Inhibition by H89 Abolished the α-MSH-Induced NO Deficiency and Inhibition of NO/NOS Signaling
3. Discussion
4. Materials and Methods
4.1. Peptides and Antibodies
4.2. Endothelial Cells Cultures
4.3. Nitric Oxide Measurement
4.4. Boyden Chamber Assay
4.5. Tube Formation Assay
4.6. Aortic Ring Assay
4.7. Angiogenesis in Zebrafish Model
4.8. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
4.9. Western Blot Analysis
4.10. Immunofluorescence Assay
4.11. MC1-R and MC2-R Gene Silencing Assay
4.12. NF-κB Activity Assay
4.13. Animal Use and Care Ethics
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-MSH | α-melanocyte-stimulating hormone |
| EGFP | Enhanced Green Fluorescent Protein |
| eNOS | endothelial nitric oxide synthase |
| HPA | hypothalamus/pituitary/adrenal gland |
| HUVEC | human umbilical vein endothelial cell |
| iNOS | inducible nitric oxide synthase |
| ISV | intersegmental vessels |
| l-Arg | l-arginine |
| MAPK | mitogen-activated kinase |
| MC1-R | melanocortin-1 receptor |
| MC2-R | melanocortin-2 receptor |
| NF-κB | nuclear factor kappa B |
| NTG | nitroglycerin |
| PKA | protein kinase A |
| POMC | proopiomelanocortin |
| sGC | soluble guanylyl cyclase |
| SD | standard deviation |
| SNP | nitroprusside |
| VEGF | vascular endothelial growth factor |
| VEGFR2 | vascular endothelial growth factor receptor-2 |
References
- Brzoska, T.; Luger, T.A.; Maaser, C.; Abels, C.; Bohm, M. α-melanocyte-stimulating hormone and related tripeptides: Biochemistry, antiinflammatory and protective effects in vitro and in vivo, and future perspectives for the treatment of immune-mediated inflammatory diseases. Endocr. Rev. 2008, 29, 581–602. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.T.; Lee, D.; Koo, K.; Lee, J.; Yoon, H.S.; Choi, Y.M.; Kwon, T.R.; Kim, B.J. Superoxide dismutase 1 inhibits α-melanocyte stimulating hormone and ultraviolet B-induced melanogenesis in murine skin. Ann. Dermatol. 2014, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Cyr, N.E.; Steger, J.S.; Toorie, A.M.; Yang, J.Z.; Stuart, R.; Nillni, E.A. Central Sirt1 regulates body weight and energy expenditure along with the POMC-derived peptide α-MSH and the processing enzyme CPE production in diet-induced obese male rats. Endocrinology 2015, 156, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Lopez, A.D. Measuring the global burden of disease. N. Engl. J. Med. 2013, 369, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Nabel, E.G.; Braunwald, E. A tale of coronary artery disease and myocardial infarction. N. Engl. J. Med. 2012, 366, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Gatti, S.; Colombo, G.; Buffa, R.; Turcatti, F.; Garofalo, L.; Carboni, N.; Ferla, L.; Fassati, L.R.; Lipton, J.M.; Catania, A. α-Melanocyte-stimulating hormone protects the allograft in experimental heart transplantation. Transplantation 2002, 74, 1678–1684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, L.; Liu, X.; Jiang, Y.; Zhang, X.; Li, X.; Zhang, Y. α-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS ONE 2014, 9, e93433. [Google Scholar] [CrossRef] [PubMed]
- Edling, A.E.; Gomes, D.; Weeden, T.; Dzuris, J.; Stefano, J.; Pan, C.; Williams, J.; Kaplan, J.; Perricone, M.A. Immunosuppressive activity of a novel peptide analog of α-melanocyte stimulating hormone (α-MSH) in experimental autoimmune uveitis. J. Neuroimmunol. 2011, 236, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fabrikant, J.; Touloei, K.; Brown, S.M. A review and update on melanocyte stimulating hormone therapy: Afamelanotide. J. Drugs Dermatol. 2013, 12, 775–779. [Google Scholar] [PubMed]
- Harms, J.; Lautenschlager, S.; Minder, C.E.; Minder, E.I. An α-melanocyte-stimulating hormone analogue in erythropoietic protoporphyria. N. Engl. J. Med. 2009, 360, 306–307. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, D.; Bitto, A.; Galantucci, M.; Zaffe, D.; Ottani, A.; Irrera, N.; Neri, L.; Cavallini, G.M.; Altavilla, D.; Botticelli, A.R.; et al. Melanocortins protect against progression of Alzheimer’s disease in triple-transgenic mice by targeting multiple pathophysiological pathways. Neurobiol. Aging 2014, 35, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, D.; Galantucci, M.; Neri, L.; Canalini, F.; Calevro, A.; Bitto, A.; Ottani, A.; Vandini, E.; Sena, P.; Sandrini, M.; et al. Melanocortins protect against brain damage and counteract cognitive decline in a transgenic mouse model of moderate Alzheimers disease. Eur. J. Pharmacol. 2014, 740, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Savos, A.V.; Gee, J.M.; Zierath, D.; Becker, K.J. α-MSH: A potential neuroprotective and immunomodulatory agent for the treatment of stroke. J. Cereb. Blood Flow Metabol. 2011, 31, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Holloway, P.M.; Smith, H.K.; Renshaw, D.; Flower, R.J.; Getting, S.J.; Gavins, F.N. Targeting the melanocortin receptor system for anti-stroke therapy. Trends Pharmacol. Sci. 2011, 32, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.S.; Tsai, H.E.; Weng, W.T.; Liu, L.F.; Weng, C.H.; Chuang, M.R.; Lam, H.C.; Wu, C.S.; Tee, R.; Wen, Z.H.; et al. Systemic pro-opiomelanocortin expression induces melanogenic differentiation and inhibits tumor angiogenesis in established mouse melanoma. Hum. Gene Ther. 2011, 22, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.E.; Liu, L.F.; Dusting, G.J.; Weng, W.T.; Chen, S.C.; Kung, M.L.; Tee, R.; Liu, G.S.; Tai, M.H. Pro-opiomelanocortin gene delivery suppresses the growth of established Lewis lung carcinoma through a melanocortin-1 receptor-independent pathway. J. Gene Med. 2012, 14, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.E.; Liu, G.S.; Kung, M.L.; Liu, L.F.; Wu, J.C.; Tang, C.H.; Huang, C.H.; Chen, S.C.; Lam, H.C.; Wu, C.S.; et al. Downregulation of hepatoma-derived growth factor contributes to retarded lung metastasis via inhibition of epithelial-mesenchymal transition by systemic POMC gene delivery in melanoma. Mol. Cancer Ther. 2013, 12, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; Diano, S. α-Melanocyte stimulating hormone: Production and degradation. J. Mol. Med. 2010, 88, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- An, J.J.; Rhee, Y.; Kim, S.H.; Kim, D.M.; Han, D.H.; Hwang, J.H.; Jin, Y.J.; Cha, B.S.; Baik, J.H.; Lee, W.T.; et al. Peripheral effect of α-melanocyte-stimulating hormone on fatty acid oxidation in skeletal muscle. J. Biol. Chem. 2007, 282, 2862–2870. [Google Scholar] [CrossRef] [PubMed]
- Galusca, B.; Prevost, G.; Germain, N.; Dubuc, I.; Ling, Y.; Anouar, Y.; Estour, B.; Chartrel, N. Neuropeptide Y and α-MSH circadian levels in two populations with low body weight: Anorexia nervosa and constitutional thinness. PLoS ONE 2015, 10, e0122040. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.T.; Huang, S.C.; Ma, Y.L.; Chan, H.H.; Lin, S.W.; Wu, J.C.; Wu, C.Y.; Wen, Z.H.; Wang, E.M.; Wu, C.L.; et al. α-Melanocyte-stimulating hormone inhibits angiogenesis through attenuation of VEGF/VEGFR2 signaling pathway. Biochim. Biophys. Acta 2014, 1840, 1850–1860. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Shimokawa, H.; Tang, E.H.; Feletou, M. Endothelial dysfunction and vascular disease. Acta Physiol. 2009, 196, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benazzi, C.; Al-Dissi, A.; Chau, C.H.; Figg, W.D.; Sarli, G.; de Oliveira, J.T.; Gartner, F. Angiogenesis in spontaneous tumors and implications for comparative tumor biology. Sci. World J. 2014, 2014, 919570. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, S.K.; Chaudhary, M.; Bagde, S.; Gadbail, A.R.; Joshi, V. Nitric oxide and cancer: A review. World J. Surg. Oncol. 2013, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Michel, T.; Vanhoutte, P.M. Cellular signaling and NO production. Pflug. Arch. 2010, 459, 807–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Nitrite and nitrate measurement by Griess reagent in human plasma: Evaluation of interferences and standardization. Methods Enzymol. 2008, 440, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Li, R.F.; Wu, T.Y.; Mou, Y.Z.; Wang, Y.S.; Chen, C.L.; Wu, C.Y. Nr2f1b control venous specification and angiogenic patterning during zebrafish vascular development. J. Biomed. Sci. 2015, 22, 104. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.R.; Nie, S.D.; Shan, W.; Jiang, D.J.; Shi, R.Z.; Zhou, Z.; Guo, R.; Zhang, Z.; Li, Y.J. Decrease in endogenous CGRP release in nitroglycerin tolerance: Role of ALDH-2. Eur. J. Pharmacol. 2007, 571, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortch-Carnes, J.; Tosini, G. Melatonin receptor agonist-induced reduction of SNP-released nitric oxide and cGMP production in isolated human non-pigmented ciliary epithelial cells. Exp. Eye Res. 2013, 107, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiyama, T.; Campbell, I.L.; Furukawa, S.; Catania, A.; Lipton, J.M. Autocrine α-melanocyte-stimulating hormone inhibits NF-κB activation in human glioma. J. Neurosci. Res. 1999, 58, 684–689. [Google Scholar] [CrossRef]
- Caruso, C.; Carniglia, L.; Durand, D.; Gonzalez, P.V.; Scimonelli, T.N.; Lasaga, M. Melanocortin 4 receptor activation induces brain-derived neurotrophic factor expression in rat astrocytes through cyclic AMP-protein kinase A pathway. Mol. Cell. Endocrinol. 2011, 348, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Cho, D.H.; Kim, H.S.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Park, H.J. Expression of the corticotropin-releasing hormone-proopiomelanocortin axis in the various clinical types of psoriasis. Exp. Dermatol. 2007, 16, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Kolgazi, M.; Arbak, S.; Alican, I. The effect of α-melanocyte stimulating hormone on gentamicin-induced acute nephrotoxicity in rats. J. Appl. Toxicol. 2007, 27, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.C.; Kuo, S.M.; Chuang, M.J.; Keng, H.M.; Lin, P.R.; Liu, G.S.; Hsu, C.M.; Howng, S.L.; Tai, M.H. Blockade of endothelin-1 release contributes to the anti-angiogenic effect by pro-opiomelanocortin overexpression in endothelial cells. Exp. Biol. Med. 2006, 231, 782–788. [Google Scholar] [CrossRef]
- Shen, P.C.; Shiau, A.L.; Jou, I.M.; Lee, C.H.; Tai, M.H.; Juan, H.Y.; Lin, P.R.; Liu, G.S.; Wu, C.L.; Hsieh, J.L. Inhibition of cartilage damage by pro-opiomelanocortin prohormone overexpression in a rat model of osteoarthritis. Exp. Biol. Med. 2011, 236, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.H.; Weng, W.T.; Lo, W.C.; Chan, J.Y.; Lin, C.J.; Lam, H.C.; Tseng, C.J. Role of nitric oxide in α-melanocyte-stimulating hormone-induced hypotension in the nucleus tractus solitarii of the spontaneously hypertensive rats. J. Pharmacol. Exp. Ther. 2007, 321, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Rinne, P.; Nordlund, W.; Heinonen, I.; Penttinen, A.M.; Saraste, A.; Ruohonen, S.T.; Makela, S.; Vahatalo, L.; Kaipio, K.; Cai, M.; et al. α-Melanocyte-stimulating hormone regulates vascular NO availability and protects against endothelial dysfunction. Cardiovasc. Res. 2012, 97, 360–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bir, S.C.; Xiong, Y.; Kevil, C.G.; Luo, J. Emerging role of PKA/eNOS pathway in therapeutic angiogenesis for ischaemic tissue diseases. Cardiovasc. Res. 2012, 95, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corander, M.P.; Fenech, M.; Coll, A.P. Science of self-preservation: How melanocortin action in the brain modulates body weight, blood pressure, and ischemic damage. Circulation 2009, 120, 2260–2268. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Laorden, B.L.; Sanchez-Mas, J.; Martinez-Alonso, E.; Martinez-Menarguez, J.A.; Garcia-Borron, J.C.; Jimenez-Cervantes, C. Dimerization of the human melanocortin 1 receptor: Functional consequences and dominant-negative effects. J. Investig. Dermatol. 2006, 126, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.M.; Lin, C.Y.; Lam, H.C.; Lin, P.R.; Chan, H.H.; Tseng, J.C.; Sun, C.K.; Hsu, T.F.; Wu, C.C.; Yang, C.Y.; et al. PTEN overexpression attenuates angiogenic processes of endothelial cells by blockade of endothelin-1/endothelin B receptor signaling. Atherosclerosis 2010, 221, 341–349. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, W.-T.; Wu, C.-S.; Wang, F.-S.; Wu, C.-Y.; Ma, Y.-L.; Chan, H.-H.; Wu, D.-C.; Wu, J.-C.; Chu, T.-H.; Huang, S.-C.; et al. α-Melanocyte-Stimulating Hormone Attenuates Neovascularization by Inducing Nitric Oxide Deficiency via MC-Rs/PKA/NF-κB Signaling. Int. J. Mol. Sci. 2018, 19, 3823. https://doi.org/10.3390/ijms19123823
Weng W-T, Wu C-S, Wang F-S, Wu C-Y, Ma Y-L, Chan H-H, Wu D-C, Wu J-C, Chu T-H, Huang S-C, et al. α-Melanocyte-Stimulating Hormone Attenuates Neovascularization by Inducing Nitric Oxide Deficiency via MC-Rs/PKA/NF-κB Signaling. International Journal of Molecular Sciences. 2018; 19(12):3823. https://doi.org/10.3390/ijms19123823
Chicago/Turabian StyleWeng, Wen-Tsan, Chieh-Shan Wu, Feng-Sheng Wang, Chang-Yi Wu, Yi-Ling Ma, Hoi-Hung Chan, Den-Chiung Wu, Jian-Ching Wu, Tian-Huei Chu, Shih-Chung Huang, and et al. 2018. "α-Melanocyte-Stimulating Hormone Attenuates Neovascularization by Inducing Nitric Oxide Deficiency via MC-Rs/PKA/NF-κB Signaling" International Journal of Molecular Sciences 19, no. 12: 3823. https://doi.org/10.3390/ijms19123823
APA StyleWeng, W.-T., Wu, C.-S., Wang, F.-S., Wu, C.-Y., Ma, Y.-L., Chan, H.-H., Wu, D.-C., Wu, J.-C., Chu, T.-H., Huang, S.-C., & Tai, M.-H. (2018). α-Melanocyte-Stimulating Hormone Attenuates Neovascularization by Inducing Nitric Oxide Deficiency via MC-Rs/PKA/NF-κB Signaling. International Journal of Molecular Sciences, 19(12), 3823. https://doi.org/10.3390/ijms19123823