The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Frequency of CMV-Specific Cells
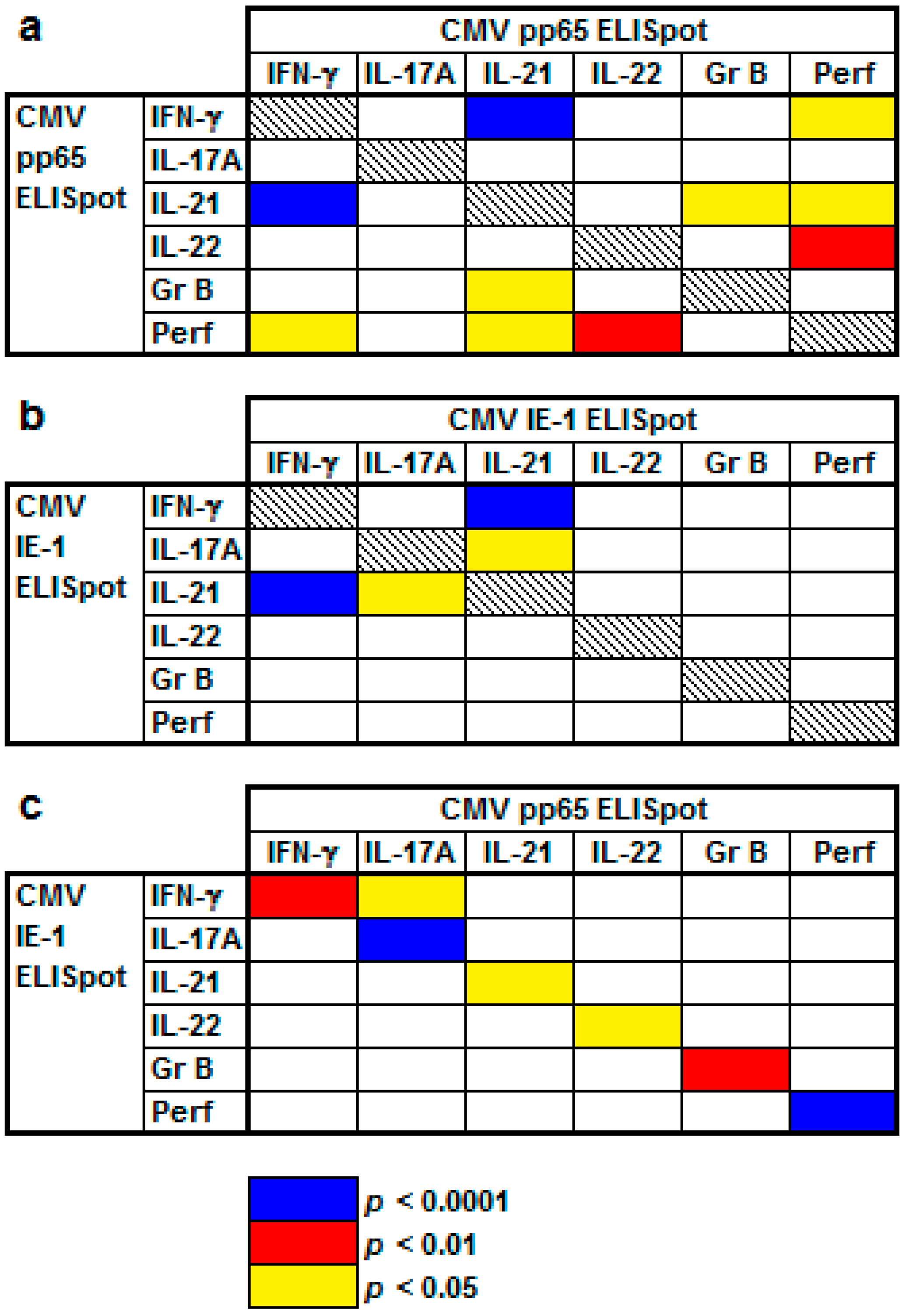
2.2. Correlation of Results to Various CMV-Specific ELISpot Assays
2.3. Correlation between Results to Flow Cytometry and CMV-Specific ELISpot Assays
2.4. Correlation of Clinical Parameters and ELISpot Results
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. ELISpot Assay
4.3. Flow Cytometry
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CMV | cytomegalovirus |
| eGFR | estimated glomerular filtration rate |
| HLA | human leukocyte antigen |
| IE-1 | immediate early antigen 1 |
| IFN | interferon |
| IL | interleukin |
| Ig | immunoglobulin |
| MMF | mycophenolate mofetil |
| mTOR | mechanistic (mammalian) Target of Rapamycin |
| pp65 | phosphoprotein 65 |
| SEM | standard error of the mean |
| SFC | spot forming cells |
References
- Azevedo, L.S.; Pierrotti, L.C.; Abdala, E.; Costa, S.F.; Strabelli, T.M.; Campos, S.V.; Ramos, J.F.; Latif, A.Z.; Litvinov, N.; Maluf, N.Z.; et al. Cytomegalovirus infection in transplant recipients. Clinics 2015, 70, 515–523. [Google Scholar] [CrossRef]
- Martin-Gandul, C.; Mueller, N.J.; Pascual, M.; Manuel, O. The Impact of Infection on Chronic Allograft Dysfunction and Allograft Survival After Solid Organ Transplantation. Am. J. Transpl. 2015, 15, 3024–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godard, B.; Gazagne, A.; Gey, A.; Baptiste, M.; Vingert, B.; Pegaz-Fiornet, B.; Strompf, L.; Fridman, W.H.; Glotz, D.; Tartour, E. Optimization of an elispot assay to detect cytomegalovirus-specific CD8+ T lymphocytes. Hum. Immunol. 2004, 65, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Czerkinsky, C.; Andersson, G.; Ekre, H.P.; Nilsson, L.A.; Klareskog, L.; Ouchterlony, O. Reverse ELISPOT assay for clonal analysis of cytokine production. I. Enumeration of gamma-interferon-secreting cells. J. Immunol. Methods 1988, 110, 29–36. [Google Scholar] [CrossRef]
- Lindemann, M.; Witzke, O.; Lütkes, P.; Fiedler, M.; Kreuzfelder, E.; Philipp, T.; Roggendorf, M.; Grosse-Wilde, H. ELISpot assay as a sensitive tool to detect cellular immunity following influenza vaccination in kidney transplant recipients. Clin. Immunol. 2006, 120, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Gliga, S.; Korth, J.; Krawczyk, A.; Wilde, B.; Horn, P.A.; Witzke, O.; Lindemann, M.; Fiedler, M. T-Track-CMV and QuantiFERON-CMV assays for prediction of protection from CMV reactivation in kidney transplant recipients. J. Clin. Virol. 2018, 105, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Banas, B.; Steubl, D.; Renders, L.; Chittka, D.; Banas, M.C.; Wekerle, T.; Koch, M.; Witzke, O.; Muhlfeld, A.; Sommerer, C.; et al. Clinical validation of a novel enzyme-linked immunosorbent spot assay-based in vitro diagnostic assay to monitor cytomegalovirus-specific cell-mediated immunity in kidney transplant recipients: A multicenter, longitudinal, prospective, observational study. Transpl. Int. 2018, 31, 436–450. [Google Scholar] [CrossRef]
- Walker, S.; Fazou, C.; Crough, T.; Holdsworth, R.; Kiely, P.; Veale, M.; Bell, S.; Gailbraith, A.; McNeil, K.; Jones, S.; et al. Ex vivo monitoring of human cytomegalovirus-specific CD8+ T-cell responses using QuantiFERON-CMV. Transpl. Infect. Dis. 2007, 9, 165–170. [Google Scholar] [CrossRef]
- Reuschel, E.; Barabas, S.; Zeman, F.; Bendfeldt, H.; Rascle, A.; Deml, L.; Seelbach-Goebel, B. Functional impairment of CMV-reactive cellular immunity during pregnancy. J. Med. Virol. 2017, 89, 324–331. [Google Scholar] [CrossRef]
- Barabas, S.; Spindler, T.; Kiener, R.; Tonar, C.; Lugner, T.; Batzilla, J.; Bendfeldt, H.; Rascle, A.; Asbach, B.; Wagner, R.; et al. An optimized IFN-gamma ELISpot assay for the sensitive and standardized monitoring of CMV protein-reactive effector cells of cell-mediated immunity. BMC Immunol. 2017, 18, 14. [Google Scholar] [CrossRef]
- Banas, B.; Boger, C.A.; Luckhoff, G.; Kruger, B.; Barabas, S.; Batzilla, J.; Schemmerer, M.; Kostler, J.; Bendfeldt, H.; Rascle, A.; et al. Validation of T-Track(R) CMV to assess the functionality of cytomegalovirus-reactive cell-mediated immunity in hemodialysis patients. BMC Immunol. 2017, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.; Yaghobi, R.; Karimi, M.H.; Darbouy, M.; Azarpira, N.; Geramizadeh, B.; Malek-Hosseini, S.A.; Nikeghbalian, S. IL-17 mRNA expression and cytomegalovirus infection in liver transplant patients. Exp. Clin. Transpl. 2015, 13 (Suppl. 1), 83–89. [Google Scholar]
- Agrawal, A.; Su, H.; Chen, J.; Osann, K.; Agrawal, S.; Gupta, S. Increased IL-21 secretion by aged CD4+T cells is associated with prolonged STAT-4 activation and CMV seropositivity. Aging 2012, 4, 648–659. [Google Scholar] [CrossRef] [Green Version]
- Oral, H.B.; Kotenko, S.V.; Yilmaz, M.; Mani, O.; Zumkehr, J.; Blaser, K.; Akdis, C.A.; Akdis, M. Regulation of T cells and cytokines by the interleukin-10 (IL-10)-family cytokines IL-19, IL-20, IL-22, IL-24 and IL-26. Eur. J. Immunol. 2006, 36, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Soccal, P.M.; Doyle, R.L.; Jani, A.; Chang, S.; Akindipe, O.A.; Poirier, C.; Pavlakis, M. Quantification of cytotoxic T-cell gene transcripts in human lung transplantation. Transplantation 2000, 69, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Ott, P.A.; Berner, B.R.; Herzog, B.A.; Guerkov, R.; Yonkers, N.L.; Durinovic-Bello, I.; Tary-Lehmann, M.; Lehmann, P.V.; Anthony, D.D. CD28 costimulation enhances the sensitivity of the ELISPOT assay for detection of antigen-specific memory effector CD4 and CD8 cell populations in human diseases. J. Immunol. Methods 2004, 285, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Laurence, A.; Elias, K.M.; O’Shea, J.J. IL-21 is produced by Th17 cells and drives IL-17 production in a STAT3-dependent manner. J. Biol. Chem. 2007, 282, 34605–34610. [Google Scholar] [CrossRef]
- Van Leeuwen, E.M.; Gamadia, L.E.; Baars, P.A.; Remmerswaal, E.B.; ten Berge, I.J.; van Lier, R.A. Proliferation requirements of cytomegalovirus-specific, effector-type human CD8+ T cells. J. Immunol. 2002, 169, 5838–5843. [Google Scholar] [CrossRef]
- Solana, R.; Tarazona, R.; Aiello, A.E.; Akbar, A.N.; Appay, V.; Beswick, M.; Bosch, J.A.; Campos, C.; Cantisan, S.; Cicin-Sain, L.; et al. CMV and Immunosenescence: From basics to clinics. Immun. Ageing 2012, 9, 23. [Google Scholar] [CrossRef]
- Fletcher, J.M.; Vukmanovic-Stejic, M.; Dunne, P.J.; Birch, K.E.; Cook, J.E.; Jackson, S.E.; Salmon, M.; Rustin, M.H.; Akbar, A.N. Cytomegalovirus-specific CD4+ T cells in healthy carriers are continuously driven to replicative exhaustion. J. Immunol. 2005, 175, 8218–8225. [Google Scholar] [CrossRef]
- Pourgheysari, B.; Khan, N.; Best, D.; Bruton, R.; Nayak, L.; Moss, P.A. The cytomegalovirus-specific CD4+ T-cell response expands with age and markedly alters the CD4+ T-cell repertoire. J. Virol. 2007, 81, 7759–7765. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Shariff, N.; Cobbold, M.; Bruton, R.; Ainsworth, J.A.; Sinclair, A.J.; Nayak, L.; Moss, P.A. Cytomegalovirus seropositivity drives the CD8 T cell repertoire toward greater clonality in healthy elderly individuals. J. Immunol. 2002, 169, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Baan, C.C.; Balk, A.H.; Dijke, I.E.; Korevaar, S.S.; Peeters, A.M.; de Kuiper, R.P.; Klepper, M.; Zondervan, P.E.; Maat, L.A.; Weimar, W. Interleukin-21: An interleukin-2 dependent player in rejection processes. Transplantation 2007, 83, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.; Hirsch, H.; Cusini, A.; van Delden, C.; Manuel, O.; Meylan, P.; Boggian, K.; Mueller, N.J.; Dickenmann, M.; Members of Swiss Transplant Cohort Study. Cytomegalovirus serology and replication remain associated with solid organ graft rejection and graft loss in the era of prophylactic treatment. Transplantation 2014, 98, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Stankovic, S.; Baxter, A.G. Raising the NKT cell family. Nat. Immunol. 2010, 11, 197–206. [Google Scholar] [CrossRef]
- Barabas, S.; Gary, R.; Bauer, T.; Lindner, J.; Lindner, P.; Weinberger, B.; Jilg, W.; Wolf, H.; Deml, L. Urea-mediated cross-presentation of soluble Epstein-Barr virus BZLF1 protein. PLoS Pathog. 2008, 4, e1000198. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.K.; Rujkijyanont, P.; Neale, G.; Yang, J.; Bari, R.; Das Gupta, N.; Holladay, M.; Rooney, B.; Leung, W. Multiplex and genome-wide analyses reveal distinctive properties of KIR+ and CD56+ T cells in human blood. J. Immunol. 2013, 191, 1625–1636. [Google Scholar] [CrossRef]
- Michel, M.L.; Mendes-da-Cruz, D.; Keller, A.C.; Lochner, M.; Schneider, E.; Dy, M.; Eberl, G.; Leite-de-Moraes, M.C. Critical role of ROR-gammat in a new thymic pathway leading to IL-17-producing invariant NKT cell differentiation. Proc. Natl. Acad. Sci. USA 2008, 105, 19845–19850. [Google Scholar] [CrossRef]
- Torina, A.; Guggino, G.; La Manna, M.P.; Sireci, G. The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 440. [Google Scholar] [CrossRef]
- Kotton, C.N.; Kumar, D.; Caliendo, A.M.; Huprikar, S.; Chou, S.; Danziger-Isakov, L.; Humar, A.; The Transplantation Society International CMV Consensus Group. The Third International Consensus Guidelines on the Management of Cytomegalovirus in Solid-organ Transplantation. Transplantation 2018, 102, 900–931. [Google Scholar] [CrossRef]
- Hocker, B.; Zencke, S.; Pape, L.; Krupka, K.; Koster, L.; Fichtner, A.; Dello Strologo, L.; Guzzo, I.; Topaloglu, R.; Kranz, B.; et al. Impact of Everolimus and Low-Dose Cyclosporin on Cytomegalovirus Replication and Disease in Pediatric Renal Transplantation. Am. J. Transpl. 2016, 16, 921–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrassy, J.; Hoffmann, V.S.; Rentsch, M.; Stangl, M.; Habicht, A.; Meiser, B.; Fischereder, M.; Jauch, K.W.; Guba, M. Is cytomegalovirus prophylaxis dispensable in patients receiving an mTOR inhibitor-based immunosuppression? a systematic review and meta-analysis. Transplantation 2012, 94, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Tam, N.; Deng, R.; Chen, P.; Li, H.; Wu, L. Everolimus-based calcineurin-inhibitor sparing regimens for kidney transplant recipients: A systematic review and meta-analysis. Int. Urol. Nephrol. 2014, 46, 2035–2044. [Google Scholar] [CrossRef] [PubMed]
- Villacres, M.C.; Longmate, J.; Auge, C.; Diamond, D.J. Predominant type 1 CMV-specific memory T-helper response in humans: Evidence for gender differences in cytokine secretion. Hum. Immunol. 2004, 65, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Bernin, H.; Fehling, H.; Marggraff, C.; Tannich, E.; Lotter, H. The cytokine profile of human NKT cells and PBMCs is dependent on donor sex and stimulus. Med. Microbiol. Immunol. 2016, 205, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Benedetto, S.; Derhovanessian, E.; Steinhagen-Thiessen, E.; Goldeck, D.; Muller, L.; Pawelec, G. Impact of age, sex and CMV-infection on peripheral T cell phenotypes: Results from the Berlin BASE-II Study. Biogerontology 2015, 16, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Shearer, G.M.; Norman, J.E.; Pinto, L.A.; Marincola, F.M.; Prasad, A.; Waclawiw, M.A.; Csako, G.; Quyyumi, A.A.; Epstein, S.E. Host response to cytomegalovirus infection as a determinant of susceptibility to coronary artery disease: Sex-based differences in inflammation and type of immune response. Circulation 2000, 102, 2491–2496. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable 1 | Variable 2 | p Value |
|---|---|---|
| Detectable CMV viral load 0 | IFN-γ IE-1 ELISpot | 0.03 |
| Symptomatic CMV infection 0 | IL-22 IE-1 ELISpot | 0.02 |
| Detectable CMV viral load 0 | eGFR 1 | 0.001 |
| Detectable CMV viral load 0 | Max. eGFR 2 | 0.009 |
| Detectable CMV viral load 2 | Dosage of prednisone 1 | 0.02 |
| Symptomatic CMV infection 2 | Dosage of prednisone 1 | 0.008 |
| Patient sex | IL-21 pp65 ELISpot | 0.005 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindemann, M.; Korth, J.; Sun, M.; Xu, S.; Struve, C.; Werner, K.; Dornieden, T.; Horn, P.A.; Witzke, O.; Wilde, B. The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients. Int. J. Mol. Sci. 2018, 19, 3945. https://doi.org/10.3390/ijms19123945
Lindemann M, Korth J, Sun M, Xu S, Struve C, Werner K, Dornieden T, Horn PA, Witzke O, Wilde B. The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients. International Journal of Molecular Sciences. 2018; 19(12):3945. https://doi.org/10.3390/ijms19123945
Chicago/Turabian StyleLindemann, Monika, Johannes Korth, Ming Sun, Shilei Xu, Christoph Struve, Kai Werner, Theresa Dornieden, Peter A. Horn, Oliver Witzke, and Benjamin Wilde. 2018. "The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients" International Journal of Molecular Sciences 19, no. 12: 3945. https://doi.org/10.3390/ijms19123945
APA StyleLindemann, M., Korth, J., Sun, M., Xu, S., Struve, C., Werner, K., Dornieden, T., Horn, P. A., Witzke, O., & Wilde, B. (2018). The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients. International Journal of Molecular Sciences, 19(12), 3945. https://doi.org/10.3390/ijms19123945