Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice


Abstract
:1. Introduction
2. Results
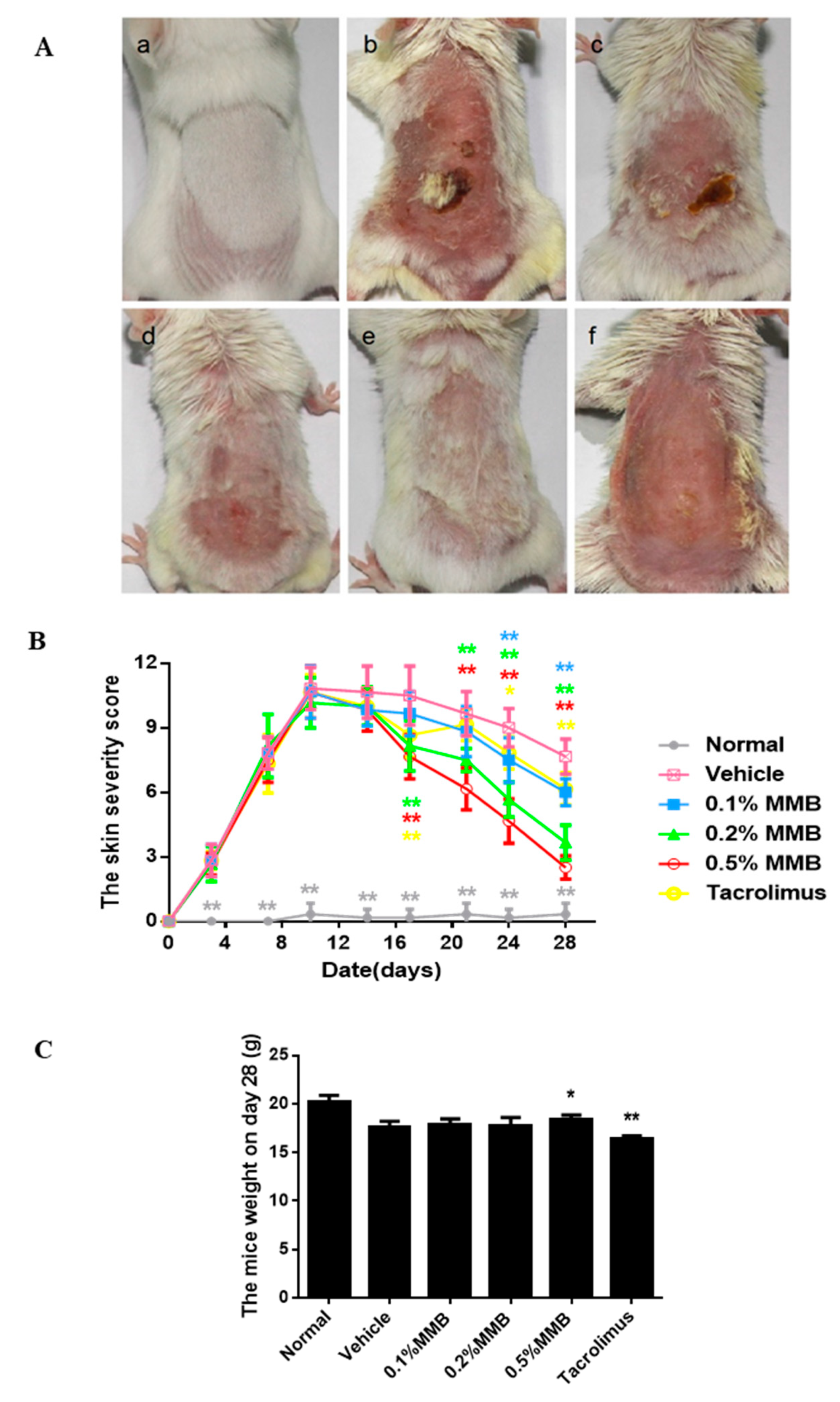
2.1. The Macroscopical Effect of MMB on AD
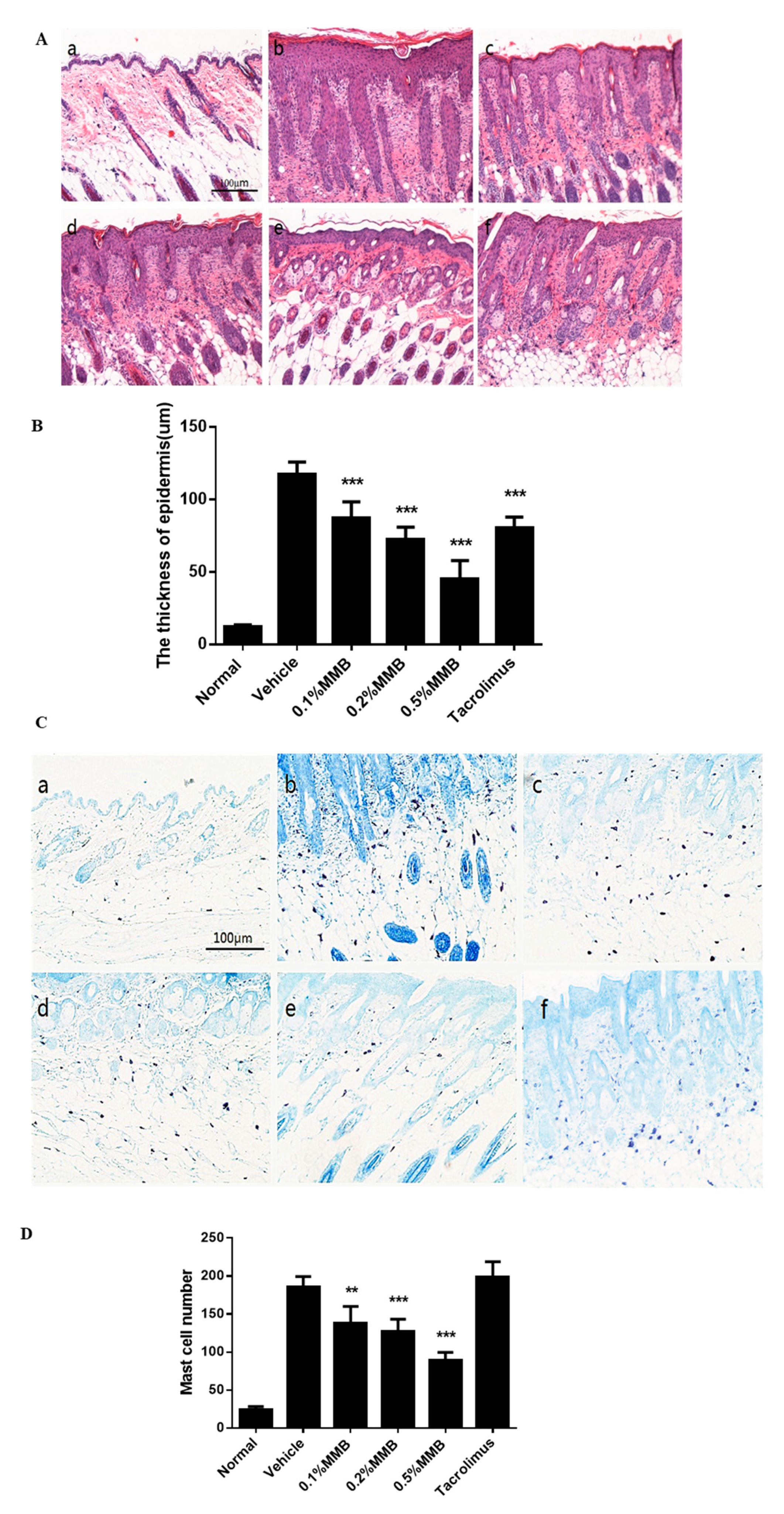
2.2. The Histological Effect of MMB on AD
2.3. MMB Suppresses IgE Generation
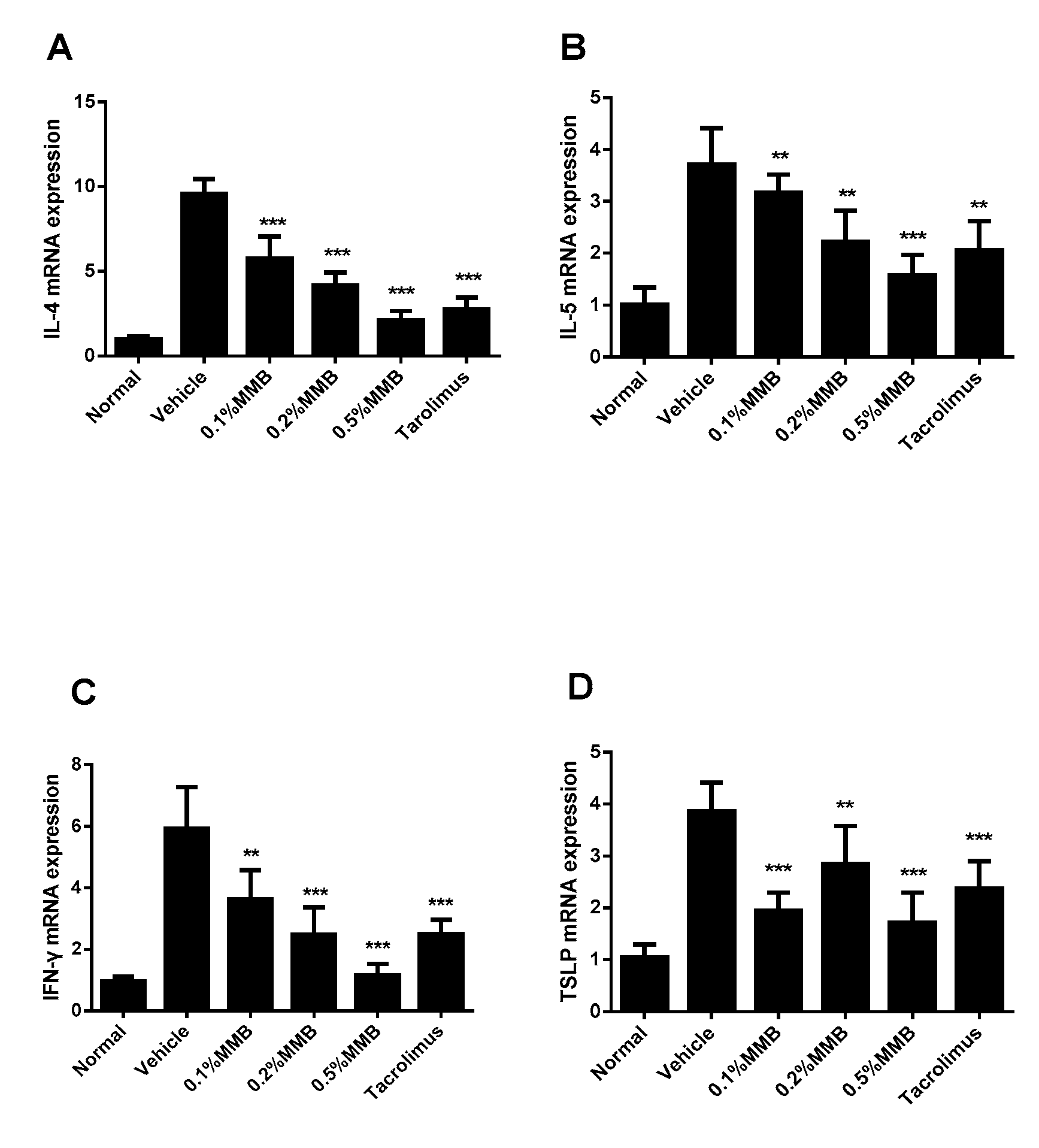
2.4. MMB Suppresses the Increased mRNA Expression of Cytokines in Skin Lesion
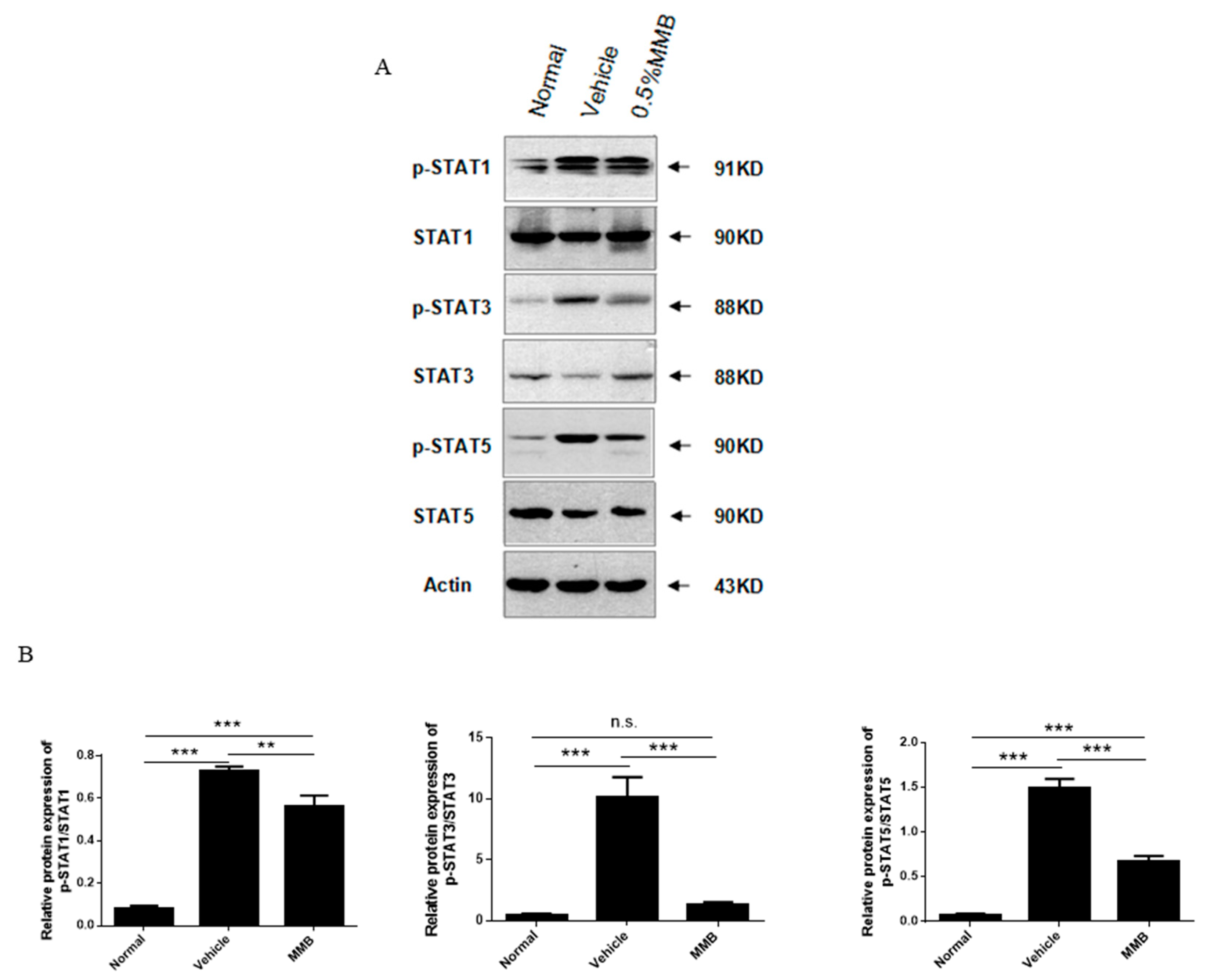
2.5. MMB Suppressed Expression of p-STAT1, p-STAT3, and p-STAT5 Proteins in Skin Lesions
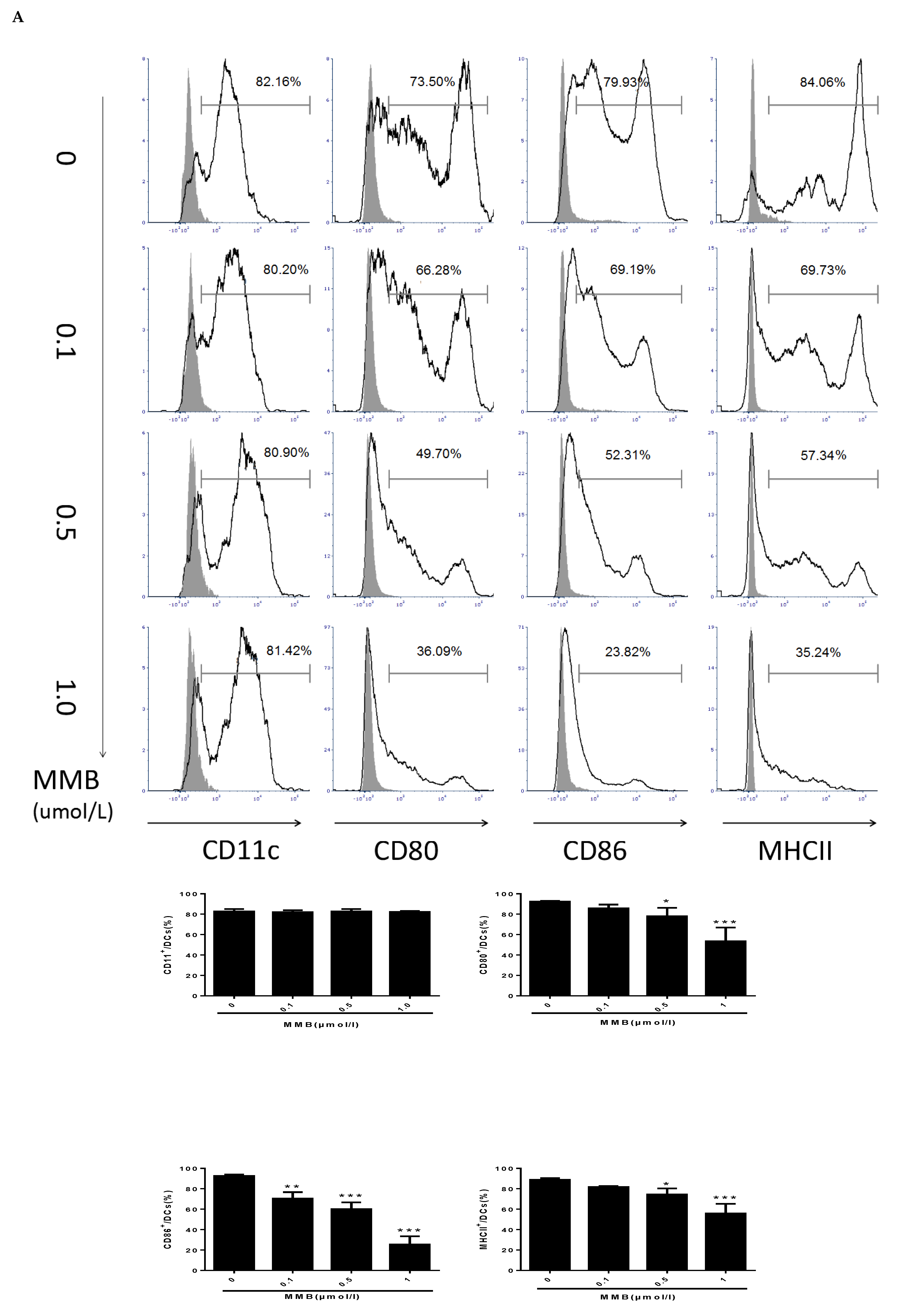
2.6. MMB Suppressed the Costimulatory Molecules, and Downmodulated the OX40L mRNA of TSLP-Stimulated DCs
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Preparation of MMB Ointment
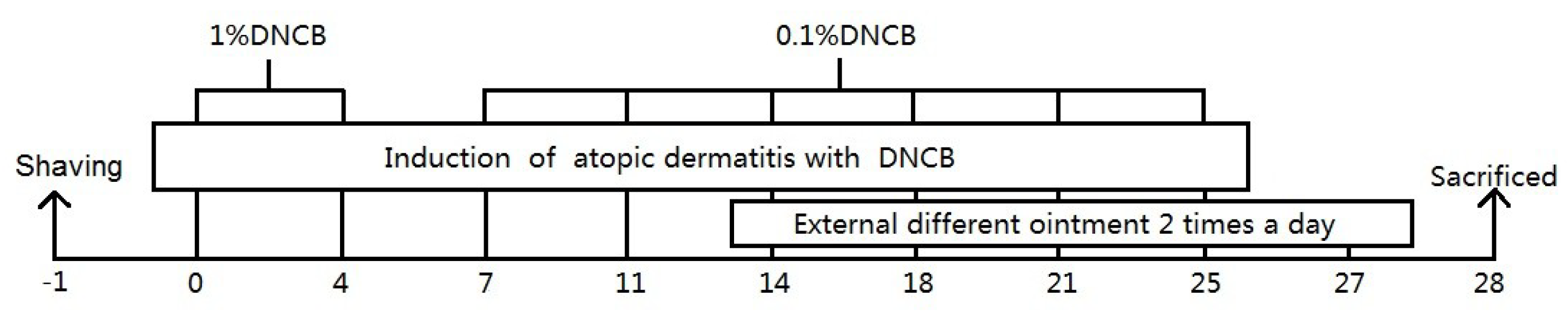
4.4. Establishment and Topical Treatment of AD Model
4.5. Macroscopic Analysis
4.6. Histopathological Analysis
4.7. ELISA Analysis
4.8. Western Blot Analysis
4.9. Preparation of DCs
4.10. Flow Cytometry
4.11. EvaGreen Real-Time Quantitative PCR
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Bieber, T. Atopic Dermatitis. Ann. Dermatol. 2010, 22, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, E.; Neri, I.; Ricci, G.; Baldo, E.; Barone, M.; Fortina, A.B.; Bernardini, R.; Berti, I.; Caffarelli, C.; Calamelli, E.; et al. Consensus conference on clinical management of pediatric atopic dermatitis. Ital. J. Pediatr. 2016, 42, 1–25. [Google Scholar] [CrossRef]
- Draelos, Z.D. Use of topical corticosteroids and topical calcineurin inhibitors for the treatment of atopic dermatitis in thin and sensitive skin areas. Curr. Med. Res. Opin. 2008, 24, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Terao, H.; Moroi, Y.; Koga, T.; Kubota, Y.; Nakayama, J.; Furukawa, F.; Tanaka, Y.; Katayama, I.; Kinukawa, N.; et al. Dosage and adverse effects of topical tacrolimus and steroids in daily management of atopic dermatitis. J. Dermatol. 2004, 31, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Montestorres, A.; Llamasvelasco, M.; Pérezplaza, A.; Solanolópez, G.; Sánchezpérez, J. Biological treatments in atopic dermatitis. J. Clin. Med. 2015, 4, 593–613. [Google Scholar] [CrossRef]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT signaling as a target for inflammatory and autoimmune diseases: Current and future prospects. Drug 2017, 77, 521–546. [Google Scholar] [CrossRef]
- Bao, L.; Zhang, H.; Chan, L.S. The involvement of the JAK-STAT signaling pathway in chronic inflammatory skin disease atopic dermatitis. JAK-STAT 2013, 2, e24137. [Google Scholar] [CrossRef]
- Schwartz, D.M.; Bonelli, M.; Gadina, M.; O’Shea, J.J. Type I/II cytokines, JAKS, and new strategies for treating autoimmune diseases. Nat. Rev. Rheumatol. 2016, 12, 25–36. [Google Scholar] [CrossRef]
- Karina, A.D.M.A.; Reinhart, S.; Eline, D.; Mireille, V.G.; Sofie, D.S.; Jo, L. JAK3 as an emerging target for topical treatment of inflammatory skin diseases. PLoS ONE 2016, 11, e0164080. [Google Scholar]
- Winthrop, K.L. The emerging safety profile of JAK inhibitors in rheumatic disease. Nat. Rev. Rheumatol. 2017, 13, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Pardanani, A.; Lasho, T.; Smith, G.; Burns, C.J.; Fantino, E.; Tefferi, A. CYT387, a selective JAK1/JAK2 inhibitor: In vitro assessment of kinase selectivity and preclinical studies using cell lines and primary cells from polycythemia vera patients. Leukemia 2009, 23, 1441–1445. [Google Scholar] [CrossRef] [PubMed]
- Hanabuchi, S.; Watanabe, N.; Liu, Y.J. TSLP and immune homeostasis. Allergol. Int. 2012, 61, 19. [Google Scholar] [CrossRef] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673. [Google Scholar] [CrossRef] [PubMed]
- Ebner, S.; Nguyen, V.A.; Forstner, M.; Wang, Y.H.; Wolfram, D.; Liu, Y.J.; Romani, N. Thymic stromal lymphopoietin converts human epidermal langerhans cells into antigen-presenting cells that induce proallergic t cells. J. Allergy Clin. Immunol. 2007, 119, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Wang, Y.H.; Duramad, O.; Hori, T.; Delespesse, G.J.; Watanabe, N.; Qin, X.F.; Yao, Z.B.; Cao, W.; Liu, Y.J. Tslp-activated dendritic cells induce an inflammatory t helper type 2 cell response through ox40 ligand. J. Exp. Med. 2005, 202, 1213–1223. [Google Scholar] [CrossRef]
- Nagaki, N.; Shiraishi, N.; Igeta, K.; Itoh, T.; Chikumoto, T.; Nagao, M.; Kim, J.F.; Nagai, H. Inhibition of scratching behavior associated with allergic dermatitis in mice by tacrolimus, but not by dexamethasone. Eur. J. Pharmacol. 2006, 546, 189–196. [Google Scholar] [CrossRef]
- Nagai, H.; Hiyama, H.; Matsuo, A.; Ueda, Y.; Inagaki, N.; Kawada, K. FK-506 and cyclosporin A potentiate the IgE antibody production by contact sensitization with hapten in mice. J. Pharmacol. Exp. Ther. 1997, 283, 321–327. [Google Scholar]
- O’Shea, J.J.; Plenge, R. JAK and STAT signaling molecules in immunoregulation and immune-mediated disease. Immunity 2012, 36, 542. [Google Scholar] [CrossRef]
- Zhou, Q.; Mrowietz, U.; Rostami-Yazdi, M. Oxidative stress in the pathogenesis of psoriasis. Free Radic. Biol. Med. 2009, 47, 891–905. [Google Scholar] [CrossRef]
- Amano, W.; Nakajima, S.; Kunugi, H.; Numata, Y.; Kitoh, A.; Egawa, G.; Dainichi, T.; Honda, T.; Otsuka, A.; Kimoto, Y.; et al. The janus kinase inhibitor JTE-052 improves skin barrier function through suppressing signal transducer and activator of transcription 3 signaling. J. Allergy Clin. Immunol. 2015, 136, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.K.; Falanga, Y.T.; Depcrynski, A.; Fernando, J.; Ryan, JJ. Mast cell homeostasis and the JAK–STAT pathway. Genes. Immun. 2010, 11, 599. [Google Scholar] [CrossRef]
- Furue, M.; Chiba, T.; Tsuji, G.; Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Kadono, T. Atopic dermatitis: Immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol. Int. 2017, 66, 398–403. [Google Scholar] [CrossRef]
- Bonilla, F.A. Stat3 mutations in the hyper-ige syndrome. N. Engl. J. Med. 2007, 357, 1608–1619. [Google Scholar] [CrossRef]
- Nomura, T.; Kabashima, K. Advances in atopic dermatitis in 2015. J. Allergy. Clin. Immunol. 2016, 138, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Welsch, K.; Holstein, J.; Laurence, A.; Ghoreschi, K. Targeting JAK/STAT signalling in inflammatory skin diseases with small molecule inhibitors. Eur. J. Immunol. 2017, 47, 1096. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Shi, V.Y.; Chan, L.S. IL-4 regulates chemokine CCL26 in keratinocytes through the JAK1, 2/Stat6 signal transduction pathway: Implication for atopic dermatitis. Mol. Immunol. 2012, 50, 91–97. [Google Scholar] [CrossRef]
- Alam, R.; Pazdrak, K.; Stafford, S.; Forsythe, P. The interleukin-5/receptor interaction activates lyn and jak2 tyrosine kinases and propagates signals via the ras-raf-1-map kinase and the JAK-STAT pathways in eosinophils. Int. Arch. Allergy Immunol. 1995, 107, 226–227. [Google Scholar] [CrossRef] [PubMed]
- Bito, T.; Roy, S.; Sen, C.K.; Shirakawa, T.; Gotoh, A.; Ueda, M.; Ichihashi, M.; Packer, L. Flavonoids differentially regulate ifnγ-induced icam-1 expression in human keratinocytes: Molecular mechanisms of action. FEBS Lett. 2002, 520, 145–152. [Google Scholar] [CrossRef]
- Liu, Y.J.; Soumelis, V.; Watanabe, N.; Ito, T.; Wang, Y.H.; Malefyt, R.D.W.; Omori, M.; Zhou, B.; Ziegler, S.F. Tslp: An epithelial cell cytokine that regulates t cell differentiation by conditioning dendritic cell maturation. Annu. Rev. Immunol. 2007, 25, 193–219. [Google Scholar] [CrossRef]
- Pierre, P.; Turley, S.J.; Gatti, E.; Hull, M.; Meltzer, J.; Mirza, A.; Inaba, K.; Steinman, R.M.; Mellman, I. Developmental regulation of MHC class II transport in mouse dendritic cells. Nature 1997, 388, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Van Rijt, L.S.; Vos, N.; Willart, M.; Kleinjan, A.; Coyle, A.J.; Hoogsteden, H.C.; Lambrecht, B.N. Essential role of dendritic cell CD80/CD86 costimulation in the induction, but not reactivation, of TH2 effector responses in a mouse model of asthma. J. Allergy Clin. Immunol. 2004, 114, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Arima, K.; Watanabe, N.; Hanabuchi, S.; Chang, M.; Sun, S.C.; Liu, Y.J. Distinct signal codes generate dendritic cell functional plasticity. Sci. Signal. 2010, 3, ra4. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.; So, T.; Duan, W.; Soroosh, P. The significance of OX40 and OX40L to T-cell biology and immune disease. Immunol. Rev. 2010, 229, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Wohlmann, A.; Sebastian, K.; Borowski, A.; Krause, S.; Friedrich, K. Signal transduction by the atopy-associated human thymic stromal lymphopoietin (TSLP) receptor depends on janus kinase function. Biol. Chem. 2010, 391, 181–186. [Google Scholar] [CrossRef]
- Yu, X.; Li, H.; Ren, X. Signaling cascades initiated by tslp-mediated signals in different cell types. Cell Immunol. 2012, 279, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Jeong, E.S.; Heo, S.H.; Seo, J.H.; Jeong, D.G.; Choi, Y.K. A novel model for human atopic dermatitis: Application of repeated DNCB patch in BALB/c mice, in comparison with NC/NGA mice. Lab. Anim. Res. 2010, 26, 95. [Google Scholar] [CrossRef]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.; Rössner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Upstream Primer Sequences(5′-3′) | Downstream Primer Sequences(5′-3′) |
|---|---|---|
| OX40L | ACGCTAAGGCTGGTGGTCTCT | TCCTCACATCTGGTAACTGCTCCT |
| IL-4 | CACGGATGCGACAAAAATCAC | CGAAAAGCCCGAAAGAGTCTCT |
| IL-5 | TCCTCCTGCCTCCTCTTCCTGAA | TGTGATCCTCCTGCGTCCATCTG |
| IFN-γ | CCATCAGCAACAACATAAGCGTCA | CCGAATCAGCAGCGACTCCTT |
| TSLP | CTGCCATGATGAGGTGGTCTGAA | TCTGCTCACGAATTGTACTGTCCT |
| β-actin | GAGATTACTGCTCTGGCTCCTA | GGACTCATCGTACTCCTGCTTG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W.; Huang, W.; Chen, L.; Jin, M.; Wang, Q.; Gao, Z.; Jin, Z. Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice. Int. J. Mol. Sci. 2018, 19, 3973. https://doi.org/10.3390/ijms19123973
Jin W, Huang W, Chen L, Jin M, Wang Q, Gao Z, Jin Z. Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice. International Journal of Molecular Sciences. 2018; 19(12):3973. https://doi.org/10.3390/ijms19123973
Chicago/Turabian StyleJin, Wenyu, Wei Huang, Liqing Chen, Mingji Jin, Qiming Wang, Zhonggao Gao, and Zhehu Jin. 2018. "Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice" International Journal of Molecular Sciences 19, no. 12: 3973. https://doi.org/10.3390/ijms19123973
APA StyleJin, W., Huang, W., Chen, L., Jin, M., Wang, Q., Gao, Z., & Jin, Z. (2018). Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice. International Journal of Molecular Sciences, 19(12), 3973. https://doi.org/10.3390/ijms19123973