Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen siRNAs on the Differentiation Index

 , and
, and
Abstract
:1. Introduction
2. Result
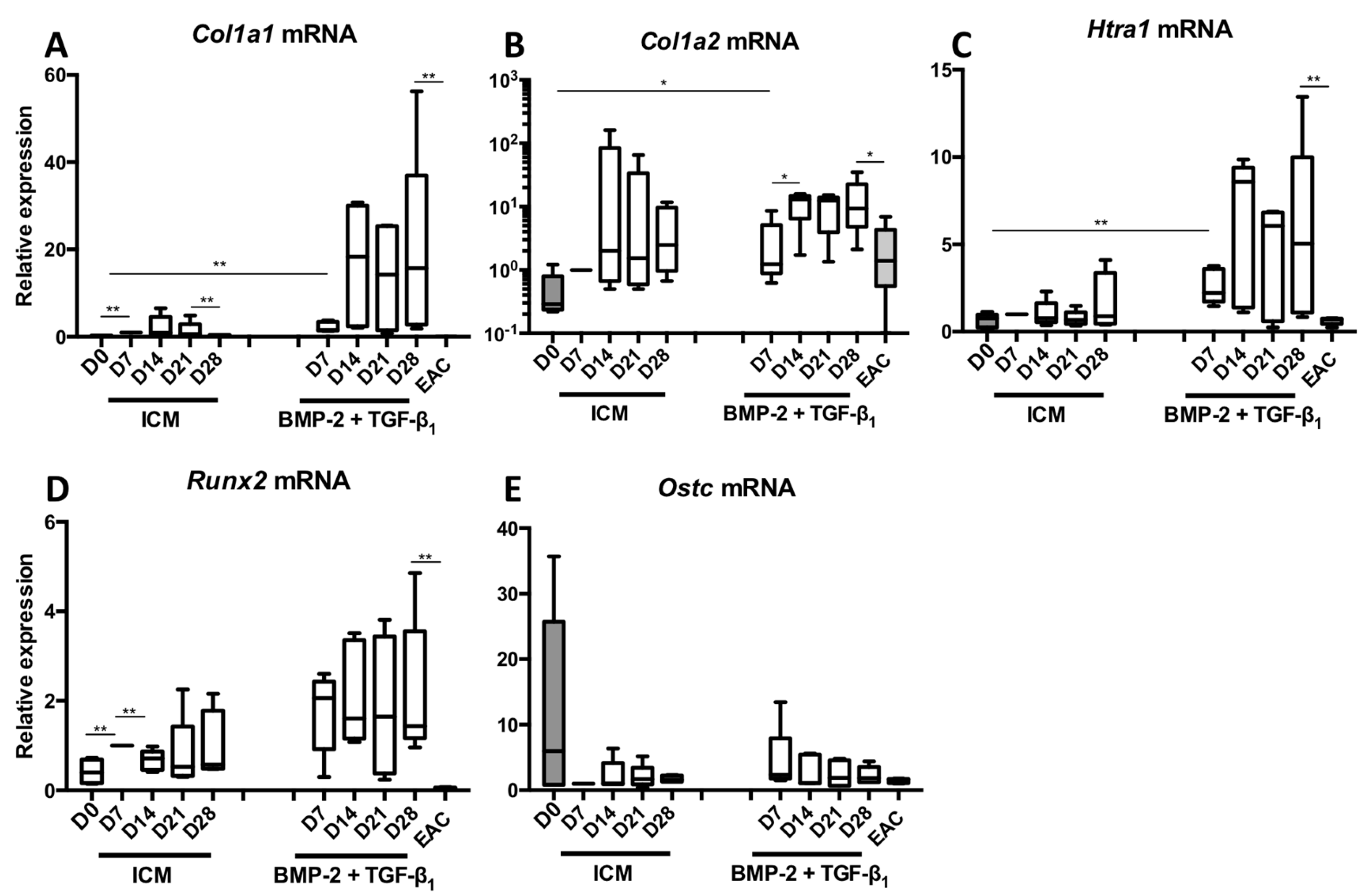
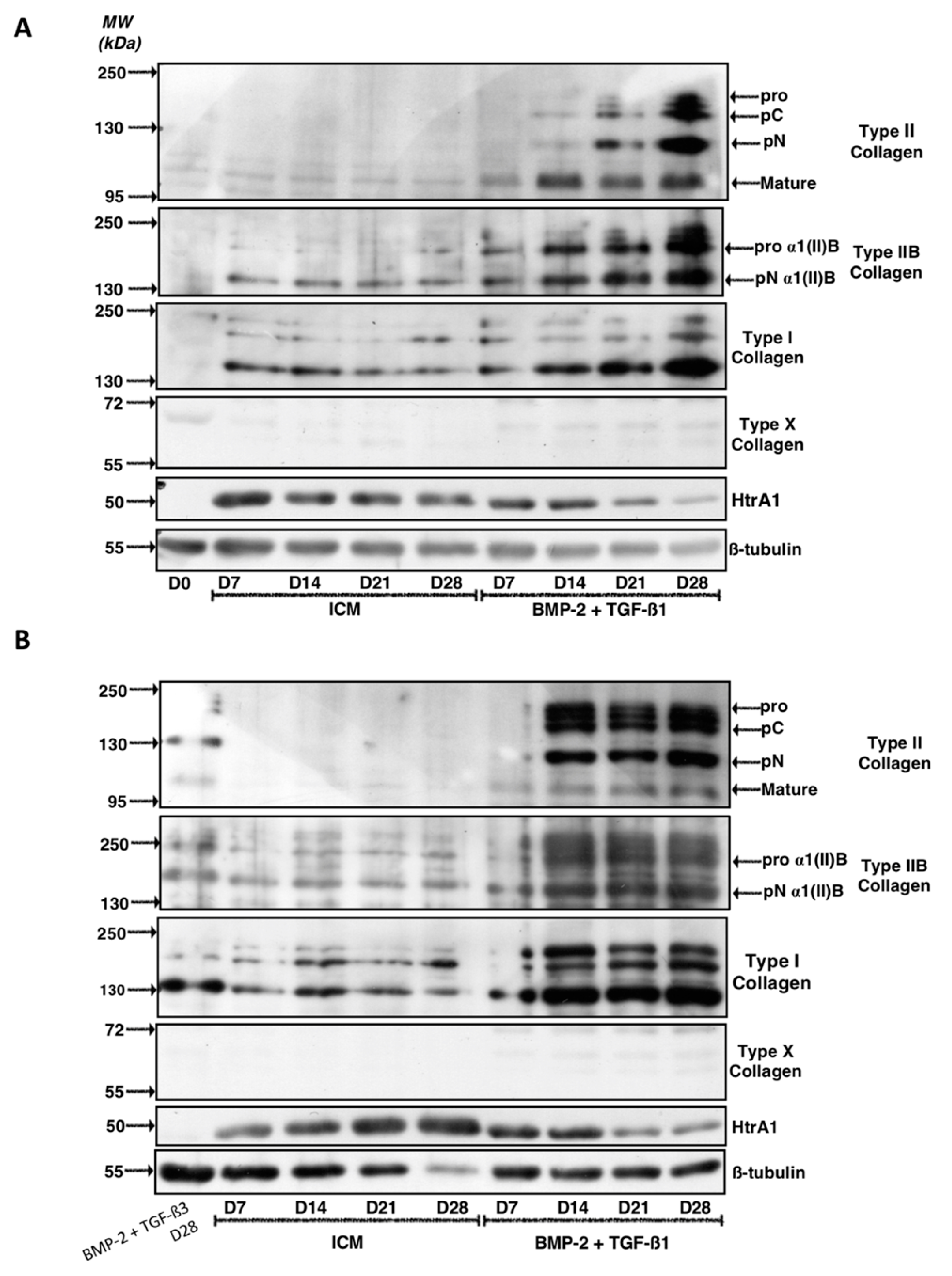
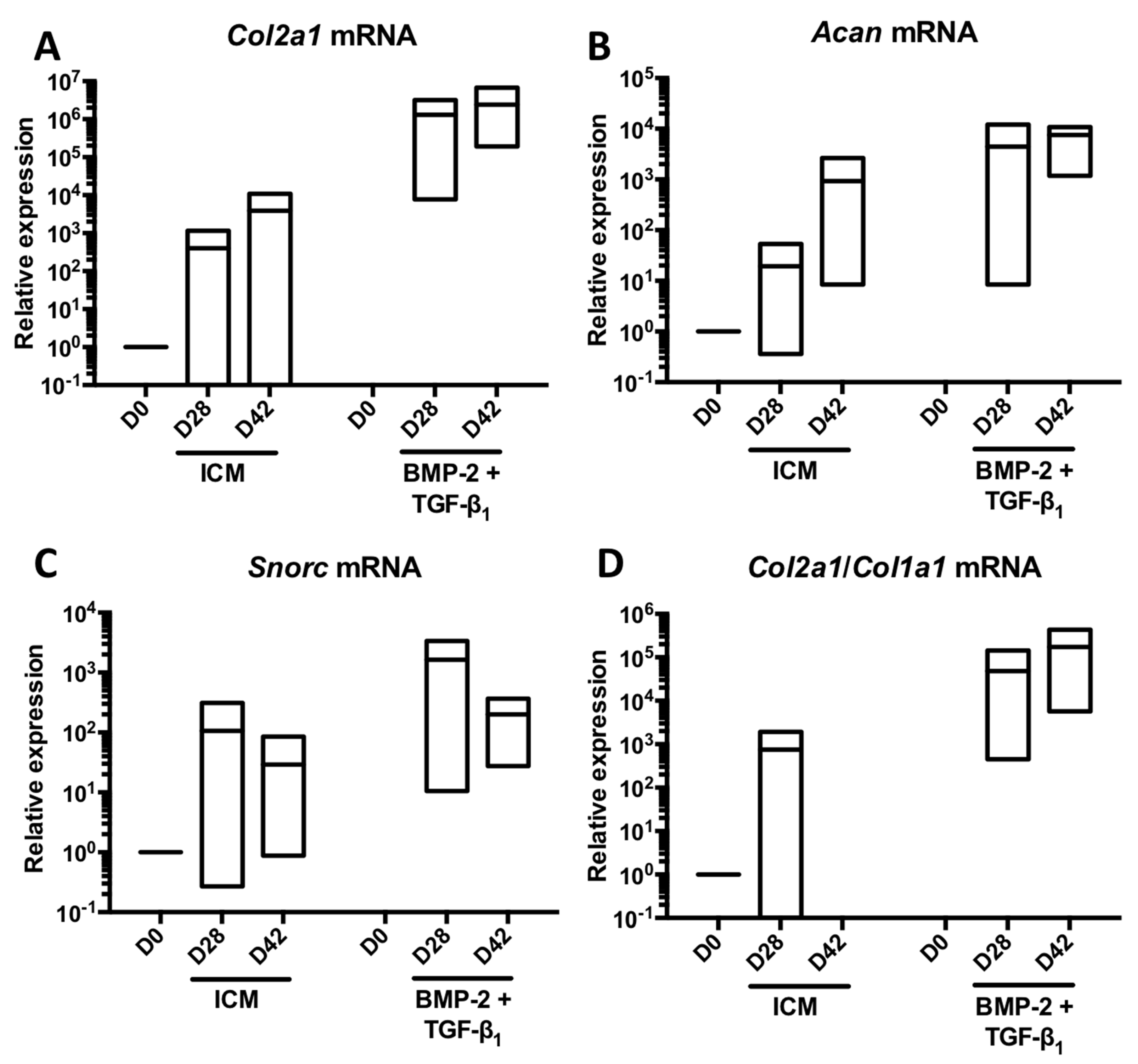
2.1. Kinetic Study of the BMP-2 and TGF-β1 Effect on the Phenotypic Status of Chondrocytes Derived from BM-MSCs
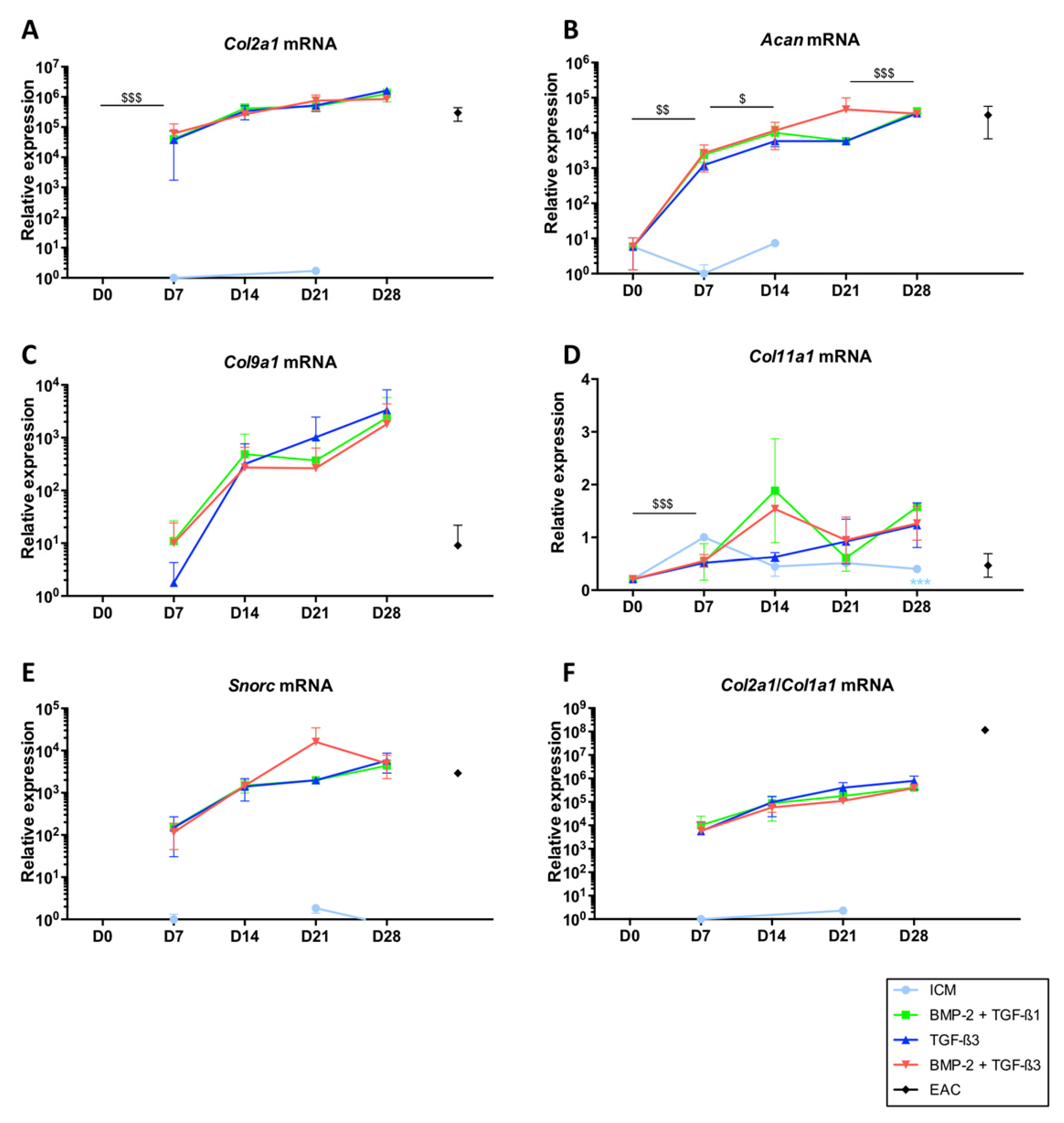
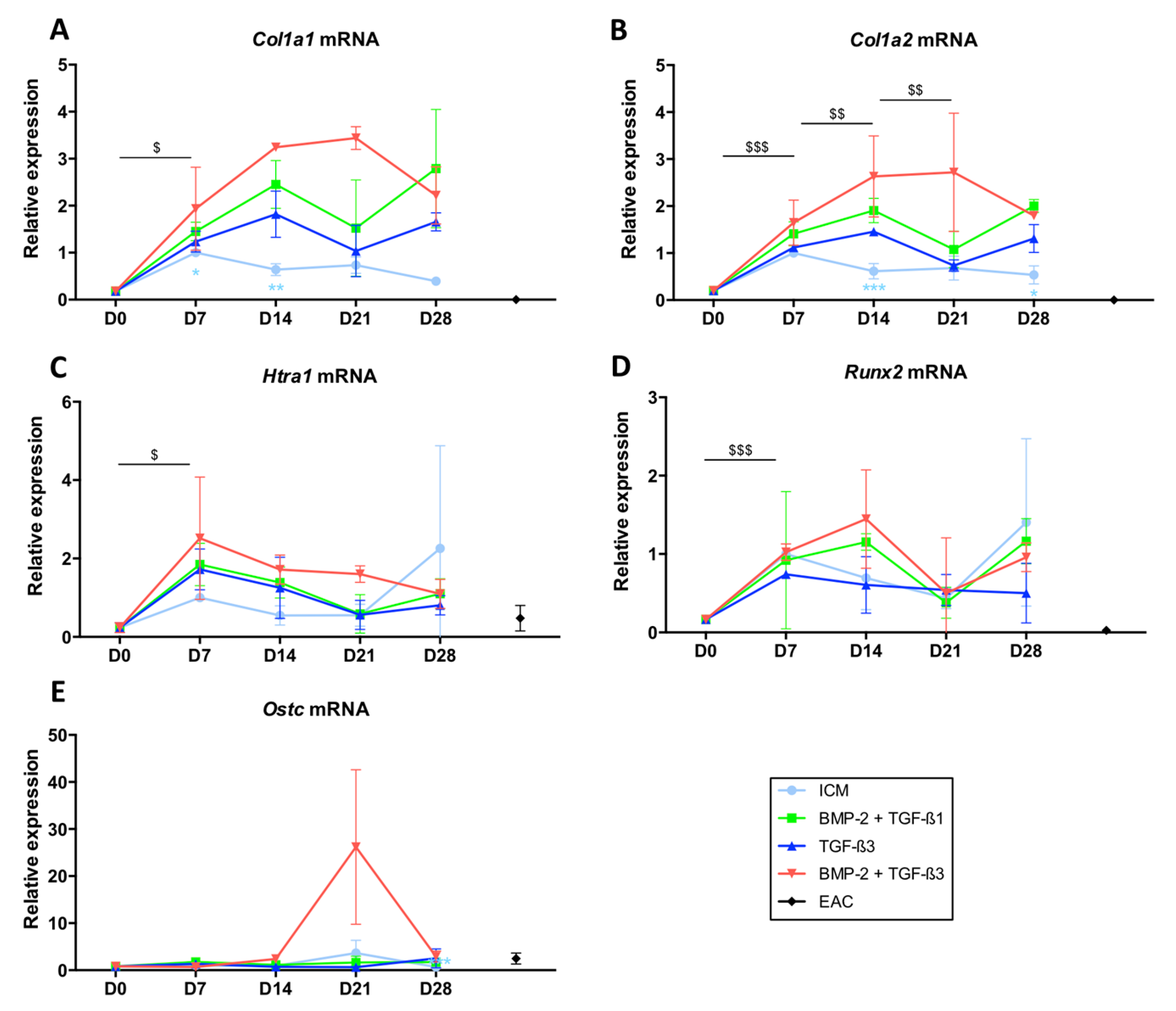
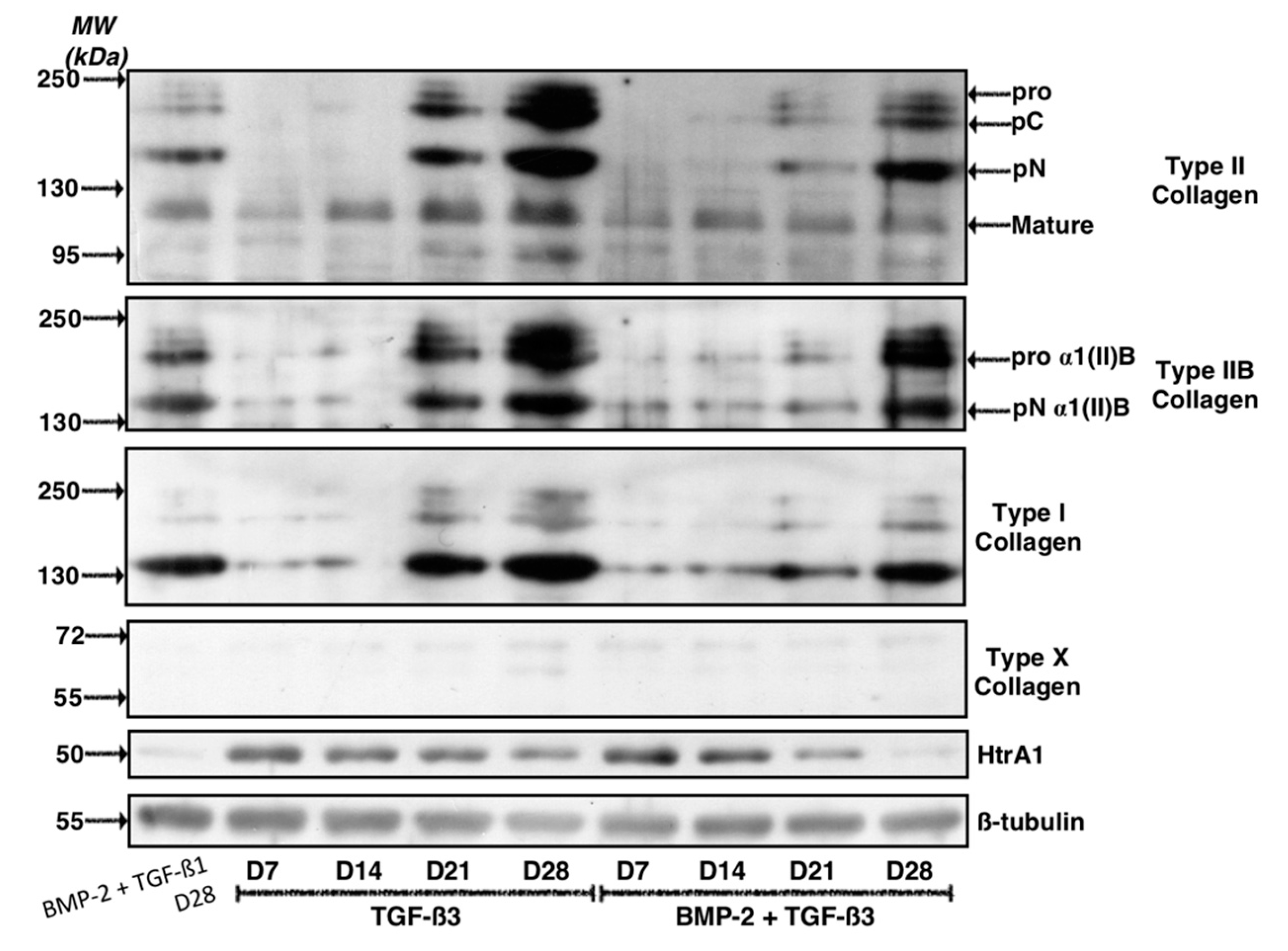
2.2. TGF-β3 Effect on the Differentiation Status of the Chondrocytes Derived from BM-MSCs
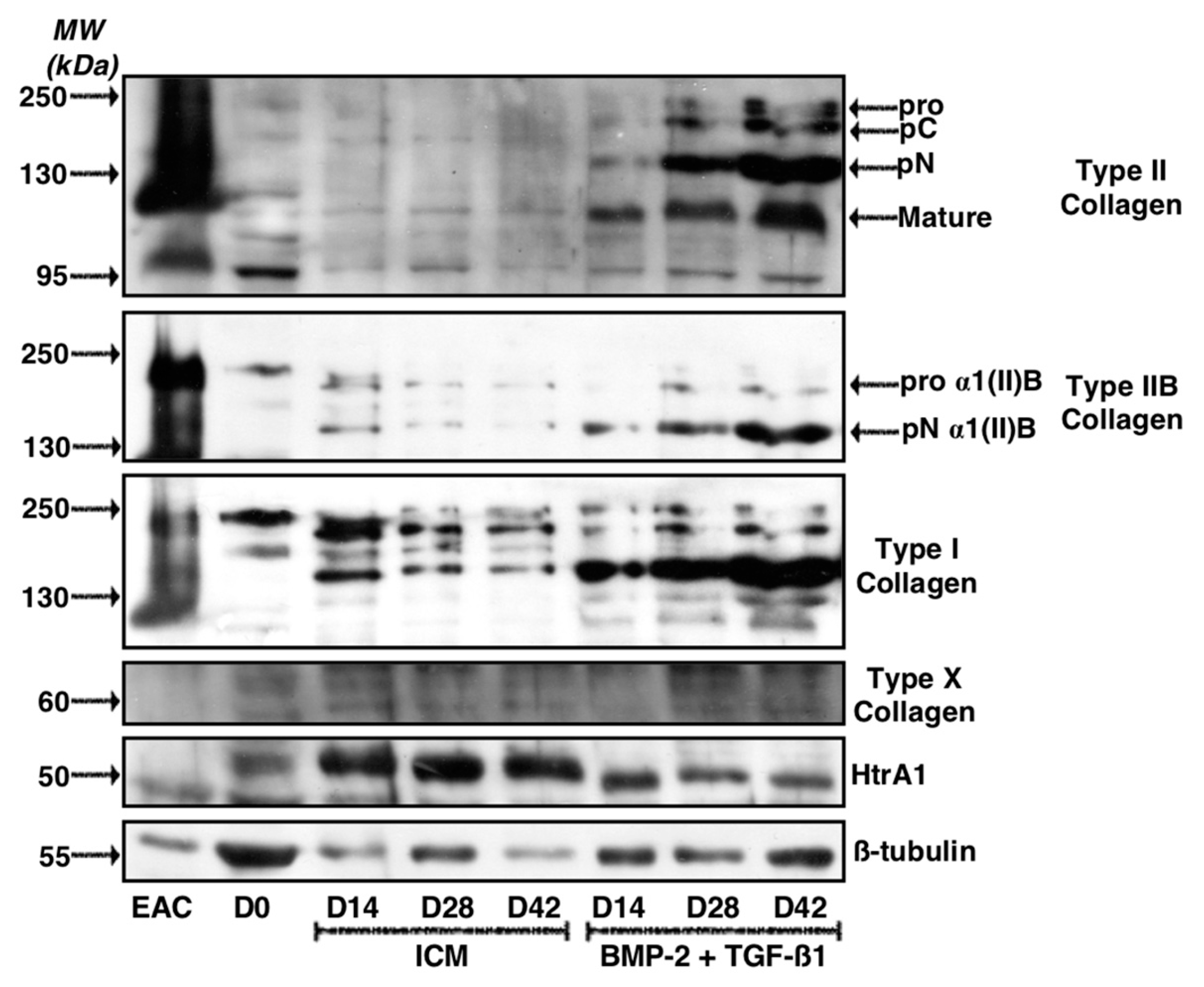
2.3. Effect of Long-Term Culture on the Phenotypic Status of the Chondrocytes Derived from BM-MSCs
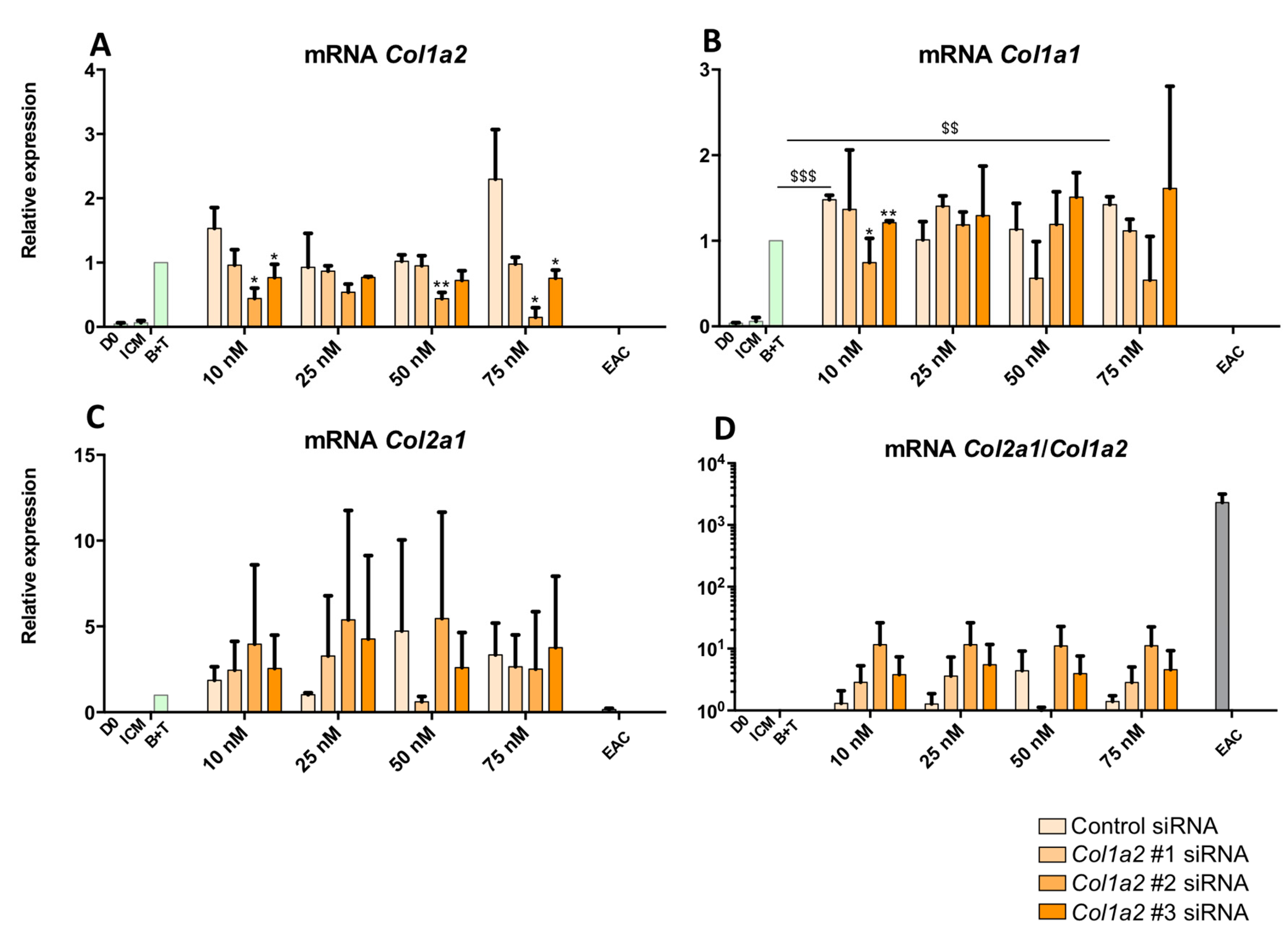
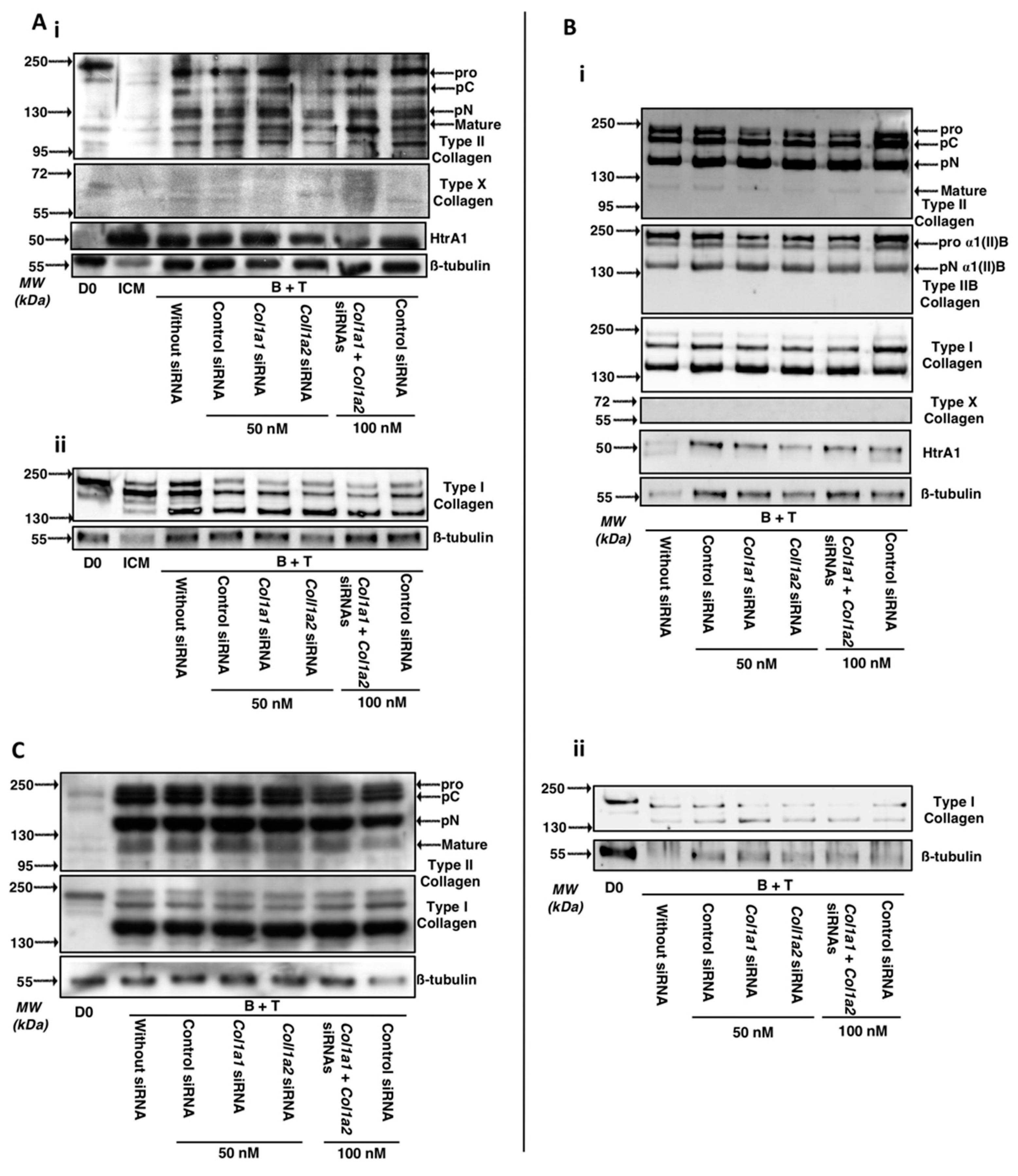
2.4. Effect of Knockdown with Col1a1 and/or Col1a2 siRNAs on the Quality of the ECM Synthesized by Chondrocytes Derived from MSCs
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Cell Culture
4.2. Chondrogenic Differentiation in 3D
4.3. RNA Interference-Knockdown of Col1a1 and/or Col1a2 mRNAs
4.4. RNA Isolation and RT-qPCR
4.5. Western Blots
4.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Aigner, T.; Stöve, J. Collagens—Major component of the physiological cartilage matrix, major target of cartilage degeneration, major tool in cartilage repair. Adv. Drug Deliv. Rev. 2003, 55, 1569–1593. [Google Scholar] [CrossRef] [PubMed]
- Sandell, L.J.; Morris, N.; Robbins, J.R.; Goldring, M.B. Alternatively spliced type II procollagen mRNAs define distinct populations of cells during vertebral development: Differential expression of the amino-propeptide. J. Cell Biol. 1991, 114, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Sandell, L.J.; Nalin, A.M.; Reife, R.A. Alternative splice form of type II procollagen mRNA (IIA) is predominant in skeletal precursors and non-cartilaginous tissues during early mouse development. Dev. Dyn. 1994, 199, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Perrier, E.; Ronzière, M.-C.; Bareille, R.; Pinzano, A.; Mallein-Gerin, F.; Freyria, A.M. Analysis of collagen expression during chondrogenic induction of human bone marrow mesenchymal stem cells. Biotechnol. Lett. 2011, 33, 2091–2101. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, J.; Taipaleenmäki, H.; Roering, P.; Takatalo, M.; Harkness, L.; Sandholm, J.; Uusitalo-Järvinen, H.; Kassem, M.; Kiviranta, I.; Laitala-Leinonen, T.; et al. Snorc is a novel cartilage specific small membrane proteoglycan expressed in differentiating and articular chondrocytes. Osteoarthr. Cartil. 2011, 19, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Benya, P.D.; Padilla, S.R.; Nimni, M.E. Independent regulation of collagen types by chondrocytes during the loss of differentiated function in culture. Cell 1978, 15, 1313–1321. [Google Scholar] [CrossRef]
- Lee, D.A.; Bentley, G.; Archer, C.W. The control of cell division in articular chondrocytes. Osteoarthr. Cartil. 1993, 1, 137–146. [Google Scholar] [CrossRef]
- Gelse, K. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberland, A.; Wang, E.; Jones, A.R.; Collins-Racie, L.A.; LaVallie, E.R.; Huang, Y.; Liu, L.; Morris, E.A.; Flannery, C.R.; Yang, Z. Identification of a novel HtrA1-susceptible cleavage site in human aggrecan: Evidence for the involvement of HtrA1 in aggrecan proteolysis in vivo. J. Biol. Chem. 2009, 284, 27352–27359. [Google Scholar] [CrossRef] [PubMed]
- Oka, C.; Tsujimoto, R.; Kajikawa, M.; Koshiba-Takeuchi, K.; Ina, J.; Yano, M.; Tsuchiya, A.; Ueta, Y.; Soma, A.; Kanda, H.; et al. HtrA1 serine protease inhibits signaling mediated by Tgfbeta family proteins. Development 2004, 131, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.-M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage tissue engineering: Molecular control of chondrocyte differentiation for proper cartilage matrix reconstruction. Biochim. Biophys. Acta 2014, 1840, 2414–2440. [Google Scholar] [CrossRef] [PubMed]
- Aigner, T.; Cook, J.L.; Gerwin, N.; Glasson, S.S.; Laverty, S.; Little, C.B.; McIlwraith, W.; Kraus, V.B. Histopathology atlas of animal model systems—Overview of guiding principles. Osteoarthr. Cartil. 2010, 18, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Malda, J.; Benders, K.E.M.; Klein, T.J.; de Grauw, J.C.; Kik, M.J.L.; Hutmacher, D.W.; Saris, D.B.F.; van Weeren, P.R.; Dhert, W.J.A. Comparative study of depth-dependent characteristics of equine and human osteochondral tissue from the medial and lateral femoral condyles. Osteoarthr. Cartil. 2012, 20, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.R.; Reid, S.; Morris, R.S. Profiling the New Zealand thoroughbred racing industry. 2. Conditions interfering with training and racing. N. Z. Vet. J. 2005, 53, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Brittberg, M.; Lindahl, A.; Nilsson, A.; Ohlsson, C.; Isaksson, O.; Peterson, L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N. Engl. J. Med. 1994, 331, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Brittberg, M. Autologous chondrocyte transplantation. Clin. Orthop. Relat. Res. 1999, 367, S147–S155. [Google Scholar] [CrossRef]
- Danišovič, L.; Boháč, M.; Zamborský, R.; Oravcová, L.; Provazníková, Z.; Csöbönyeiová, M.; Varga, I. Comparative analysis of mesenchymal stromal cells from different tissue sources in respect to articular cartilage tissue engineering. Gen. Physiol. Biophys. 2016, 35, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Delorme, B.; Ringe, J.; Pontikoglou, C.; Gaillard, J.; Langonné, A.; Sensebé, L.; Noël, D.; Jorgensen, C.; Häupl, T.; Charbord, P. Specific lineage-priming of bone marrow mesenchymal stem cells provides the molecular framework for their plasticity. Stem Cells 2009, 27, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Smith, A. Self-renewal of teratocarcinoma and embryonic stem cells. Oncogene 2004, 23, 7150–7160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlovits, S.; Aldrian, S.; Tichy, B.; Albrecht, C.; Nürnberger, S. Biomaterial for autologous chondrocyte transplantation. Orthopäde 2009, 38, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.K.; Huey, D.J.; Hu, J.C.; Athanasiou, K.A. TGF-β1, GDF-5, and BMP-2 stimulation induces chondrogenesis in expanded human articular chondrocytes and marrow-derived stromal cells. Stem Cells 2015, 33, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Kafienah, W.; Mistry, S.; Dickinson, S.C.; Sims, T.J.; Learmonth, I.; Hollander, A.P. Three-dimensional cartilage tissue engineering using adult stem cells from osteoarthritis patients. Arthritis Rheumatol. 2007, 56, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Ronzière, M.C.M.; Perrier, E.E.; Mallein-Gerin, F.F.; Freyria, A.-M. Chondrogenic potential of bone marrow- and adipose tissue-derived adult human mesenchymal stem cells. Biomed. Mater. Eng. 2010, 20, 145–158. [Google Scholar] [PubMed]
- Legendre, F.; Ollitrault, D.; Hervieu, M.; Baugé, C.; Maneix, L.; Goux, D.; Chajra, H.; Mallein-Gerin, F.; Boumediene, K.; Galera, P.; et al. Enhanced hyaline cartilage matrix synthesis in collagen sponge scaffolds by using siRNA to stabilize chondrocytes phenotype cultured with bone morphogenetic protein-2 under hypoxia. Tissue Eng. Part C Methods 2013, 19, 550–567. [Google Scholar] [CrossRef] [PubMed]
- Legendre, F.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Gruchy, N.; Mallein-Gerin, F.; Leclercq, S.; Demoor, M.; Galéra, P. Enhanced chondrogenesis of bone marrow derived-stem cells by using a combinatory cell therapy strategy with BMP-2/TGF-β1, hypoxia and COL1A1/HtrA1 siRNAs. Sci. Rep. 2017, 13, 3406. [Google Scholar] [CrossRef] [PubMed]
- Ollitrault, D.; Legendre, F.; Drougard, C.; Briand, M.; Benateau, H.; Goux, D.; Chajra, H.; Poulain, L.; Hartmann, D.; Vivien, D.; et al. BMP-2, Hypoxia, and COL1A1/HtrA1 siRNAs favor neo-cartilage hyaline matrix formation in chondrocytes. Tissue Eng. Part C Methods 2015, 21, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Branly, T.; Bertoni, L.; Contentin, R.; Rakic, R.; Gomez-Leduc, T.; Desancé, M.; Hervieu, M.; Legendre, F.; Jacquet, S.; Audigié, F.; et al. Characterization and use of equine bone marrow mesenchymal stem cells in equine cartilage engineering. Study of their hyaline cartilage forming potential when cultured under hypoxia within a biomaterial in the presence of BMP-2 and TGF-β1. Stem Cell Rev. Rep. 2017, 13, 611–630. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, R.L.; Ni, M.; Meretoja, V.V.; Kasper, F.K.; Mikos, A.G. TGF-β3-induced chondrogenesis in co-cultures of chondrocytes and mesenchymal stem cells on biodegradable scaffolds. Biomaterials 2014, 35, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.D.; Kimmerling, E.P.; Cairns, D.M.; Kaplan, D.L. Silk as a biomaterial to support long-term three-dimensional tissue cultures. ACS Appl. Mater. Interfaces 2016, 8, 21861–21868. [Google Scholar] [CrossRef] [PubMed]
- Adamson, E.D.; Ayers, S.E. The localization and synthesis of some collagen types in developing mouse embryos. Cell 1979, 16, 953–965. [Google Scholar] [CrossRef]
- Barsh, G.S.; Byers, P.H. Reduced secretion of structurally abnormal type I procollagen in a form of osteogenesis imperfecta. Proc. Natl. Acad. Sci. USA 1981, 78, 5142–5146. [Google Scholar] [CrossRef] [PubMed]
- De Wet, W.J.; Chu, M.L.; Prockop, D.J. The mRNAs for the pro-alpha 1(I) and pro-alpha 2(I) chains of type I. procollagen are translated at the same rate in normal human fibroblasts and in fibroblasts from two variants of osteogenesis imperfecta with altered steady state ratios of the two mRNAs. J. Biol. Chem. 1983, 258, 14385–14389. [Google Scholar] [PubMed]
- Galéra, P.; Park, R.W.; Ducy, P.; Mattéi, M.G.; Karsenty, G. c-Krox binds to several sites in the promoter of both mouse type, I. Collagen genes. Structure/function study and developmental expression analysis. J. Biol. Chem. 1996, 271, 21331–21339. [Google Scholar] [CrossRef] [PubMed]
- Rosová, I.; Dao, M.; Capoccia, B.; Link, D.; Nolta, J.A. Hypoxic preconditioning results in increased motility and improved therapeutic potential of human mesenchymal stem cells. Stem Cells 2008, 26, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Grayson, W.L.; Zhao, F.; Bunnell, B.; Ma, T. Hypoxia enhances proliferation and tissue formation of human mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2007, 358, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Kanichai, M.; Ferguson, D.; Prendergast, P.J.; Campbell, V.A. Hypoxia promotes chondrogenesis in rat mesenchymal stem cells: A role for AKT and hypoxia-inducible factor (HIF)-1α. J. Cell. Physiol. 2008, 216, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Schipani, E.; Ryan, H.E.; Didrickson, S.; Kobayashi, T.; Knight, M.; Johnson, R.S. Hypoxia in cartilage: HIF-1α is essential for chondrocyte growth arrest and survival. Genes Dev. 2001, 15, 2865–2876. [Google Scholar] [PubMed]
- Markway, B.D.; Cho, H.; Johnstone, B. Hypoxia promotes redifferentiation and suppresses markers of hypertrophy and degeneration in both healthy and osteoarthritic chondrocytes. Arthritis Res. Ther. 2013, 15, R92. [Google Scholar] [CrossRef] [PubMed]
- Brittberg, M. Cell carriers as the next generation of cell therapy for cartilage repair: A review of the matrix-induced autologous chondrocyte implantation procedure. Am. J. Sports Med. 2010, 38, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Ciuffreda, M.C.; Malpasso, G.; Musarò, P.; Turco, V.; Gnecchi, M. Protocols for in vitro differentiation of human mesenchymal stem cells into osteogenic, chondrogenic and adipogenic lineages. Methods Mol. Biol. 2016, 1416, 149–158. [Google Scholar] [PubMed]
- Yang, W.; Both, S.K.; van Osch, G.J.V.M.; Wang, Y.; Jansen, J.A.; Yang, F. Effects of in vitro chondrogenic priming time of bone-marrow-derived mesenchymal stromal cells on in vivo endochondral bone formation. Acta Biomater. 2015, 13, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.J.; Fisher, M.B.; Huang, A.H.; Shin, J.I.; Farrell, K.M.; Mauck, R.L. Functional properties of bone marrow-derived MSC-based engineered cartilage are unstable with very long-term in vitro culture. J. Biomech. 2014, 47, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Ryu, J.-S.; Yeo, J.E.; Choi, Y.J.; Kim, Y.S.; Ko, K.; Koh, Y.-G. Overexpression of TGF-β1 enhances chondrogenic differentiation and proliferation of human synovium-derived stem cells. Biochem. Biophys. Res. Commun. 2014, 450, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Leduc, T.; Hervieu, M.; Legendre, F.; Bouyoucef, M.; Gruchy, N.; Poulain, L.; de Vienne, C.; Herlicoviez, M.; Demoor, M.; Galera, P. Chondrogenic commitment of human umbilical cord blood-derived mesenchymal stem cells in collagen matrices for cartilage engineering. Sci. Rep. 2016, 6, 32786. [Google Scholar] [CrossRef] [PubMed]
- Barile, F.A.; Siddiqi, Z.E.; Ripley-Rouzier, C.; Bienkowski, R.S. Effects of puromycin and hydroxynorvaline on net production and intracellular degradation of collagen in human fetal lung fibroblasts. Arch. Biochem. Biophys. 1989, 270, 294–301. [Google Scholar] [CrossRef]
- Gotkin, M.G.; Ripley, C.R.; Lamande, S.R.; Bateman, J.F.; Bienkowski, R.S. Intracellular trafficking and degradation of unassociated proα2 chains of collagen type I. Exp. Cell Res. 2004, 296, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Veitia, R.A.; Bottani, S.; Birchler, J.A. Cellular reactions to gene dosage imbalance: Genomic, transcriptomic and proteomic effects. Trends Genet. 2008, 24, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.P.; Montecinos, L.; Ríos, S.; Reyes, P.; Martínez, J. Mesenchymal stem cells from osteoporotic patients produce a type, I. collagen-deficient extracellular matrix favoring adipogenic differentiation. J. Cell. Biochem. 2000, 79, 557–565. [Google Scholar] [CrossRef]
- Essex, S.; Navarro, G.; Sabhachandani, P.; Chordia, A.; Trivedi, M.; Movassaghian, S.; Torchilin, V.P. Phospholipid-modified PEI-based nanocarriers for in vivo siRNA therapeutics against multidrug-resistant tumors. Gene Ther. 2015, 22, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, J.; Lin, S.; Wu, T.; Huang, H.; Zhang, K.; Sun, Y.; Yeung, K.W.K.; Li, G.; Bian, L. Nanocarrier-mediated codelivery of small molecular drugs and siRNA to enhance chondrogenic differentiation and suppress hypertrophy of human mesenchymal stem cells. Adv. Funct. Mater. 2016, 26, 2463–2472. [Google Scholar] [CrossRef]
- Aubert-Foucher, E.; Mayer, N.; Pasdeloup, M.; Pagnon, A.; Hartmann, D.; Mallein-Gerin, F. A unique tool to selectively detect the chondrogenic IIB form of human type II procollagen protein. Matrix Biol. 2014, 34, 80–88. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) (F: Foward; R: Reverse) |
|---|---|
| β-actin | F: AGGCACCAGGGCGTGAT |
| R: CTCTTGCTCTGGGCCTCGT | |
| Acan | F: TGTCAACAACAATGCCCAAGAC |
| R: CTTCTTCCGCCCAAAGGTCC | |
| Col1a1 | F: TGCCGTGACCTCAAGATGTG |
| R: CGTCTCCATGTTGCAGAAGA | |
| Col1a2 | F: CCAGAGTGGAGCAGCGGTTA |
| R: GGGATGTTTTCAGGTTGAGCC | |
| Col2a1 | F: GGCAATAGCAGGTTCACGTACA |
| R: CGATAACAGTCTTGCCCCACTT | |
| Col9a1 | F: CCAAGAGGCCCAATCGACAT |
| R: GGGGAAGTCCGTTATCCTGG | |
| Col10a1 | F: GCACCCCAGTAATGTACACCTATG |
| R: GAGCCACACCTGGTCATTTTC | |
| Col11a1 | F: TTGCTGATGGGAAGTGGCAT |
| R: GCTGCTTTGGGGTCACCTAT | |
| Htra1 | F: GGACTTCATGTTTCCCTCAA |
| R: GTTCTGCTGAACAAGCAACA | |
| Ostc | F: AGAGTCTGGCAGAGGTGCAG |
| R: TCGTCACAGTCTGGGTTGAG | |
| Runx-2 | F: GCAGTTCCCAAGCATTTCAT |
| R: CACTCTGGCTTTGGGAAGAG | |
| Snorc | F: TTTACCAGCTCAGTCCTCGG |
| R: CAGACAGAGAGCCATCCTGG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branly, T.; Contentin, R.; Desancé, M.; Jacquel, T.; Bertoni, L.; Jacquet, S.; Mallein-Gerin, F.; Denoix, J.-M.; Audigié, F.; Demoor, M.; et al. Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen siRNAs on the Differentiation Index. Int. J. Mol. Sci. 2018, 19, 435. https://doi.org/10.3390/ijms19020435
Branly T, Contentin R, Desancé M, Jacquel T, Bertoni L, Jacquet S, Mallein-Gerin F, Denoix J-M, Audigié F, Demoor M, et al. Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen siRNAs on the Differentiation Index. International Journal of Molecular Sciences. 2018; 19(2):435. https://doi.org/10.3390/ijms19020435
Chicago/Turabian StyleBranly, Thomas, Romain Contentin, Mélanie Desancé, Thibaud Jacquel, Lélia Bertoni, Sandrine Jacquet, Frédéric Mallein-Gerin, Jean-Marie Denoix, Fabrice Audigié, Magali Demoor, and et al. 2018. "Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen siRNAs on the Differentiation Index" International Journal of Molecular Sciences 19, no. 2: 435. https://doi.org/10.3390/ijms19020435
APA StyleBranly, T., Contentin, R., Desancé, M., Jacquel, T., Bertoni, L., Jacquet, S., Mallein-Gerin, F., Denoix, J. -M., Audigié, F., Demoor, M., & Galéra, P. (2018). Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen siRNAs on the Differentiation Index. International Journal of Molecular Sciences, 19(2), 435. https://doi.org/10.3390/ijms19020435