The Aquaporin 1 Inhibitor Bacopaside II Reduces Endothelial Cell Migration and Tubulogenesis and Induces Apoptosis

 , , and
, , and 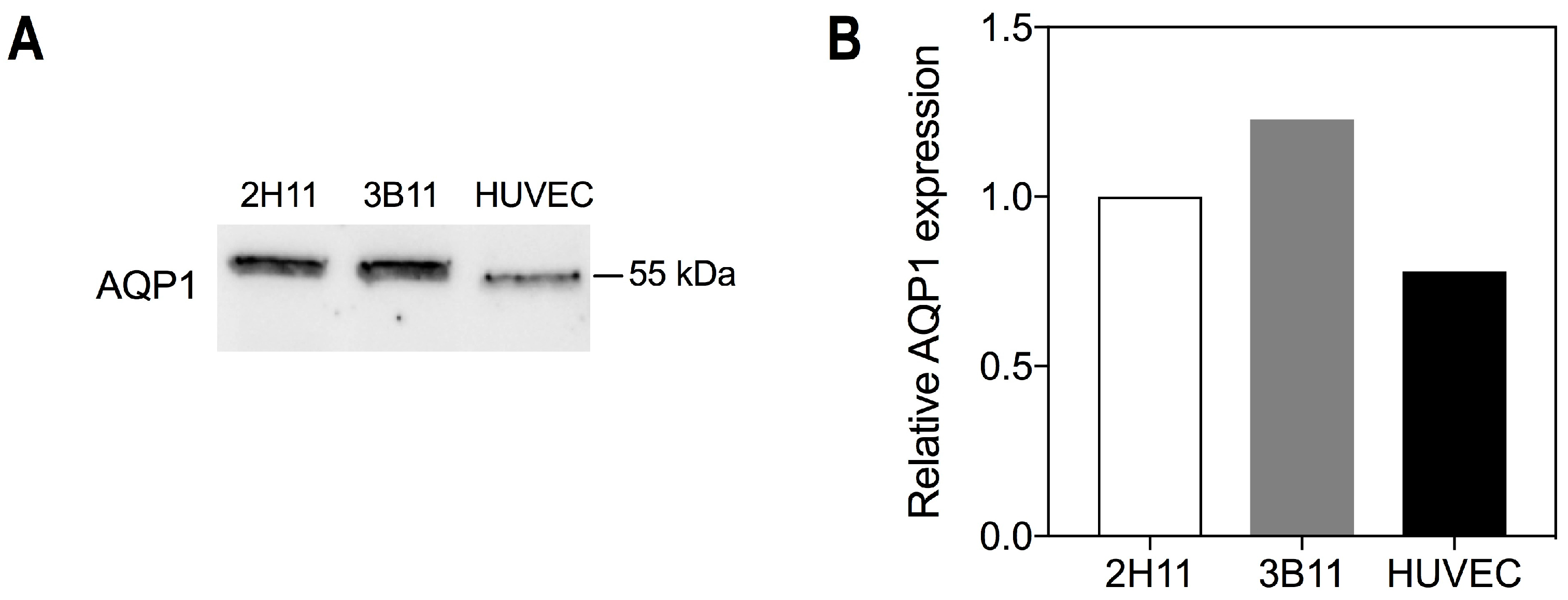{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Endothelial Cells Express the Bacopaside II Target AQP1
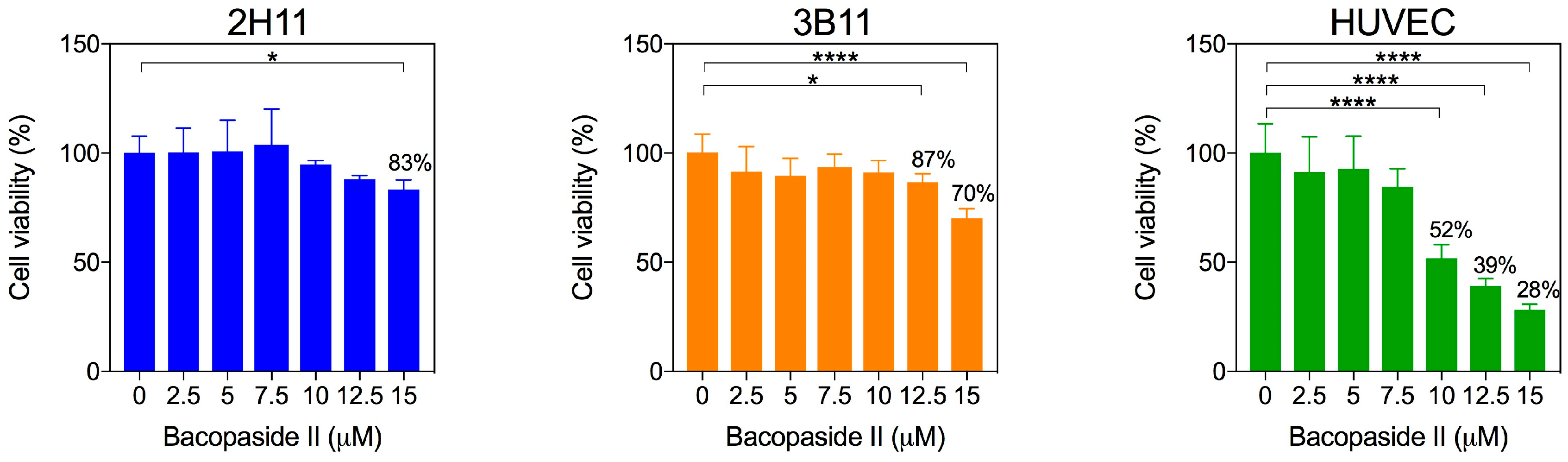
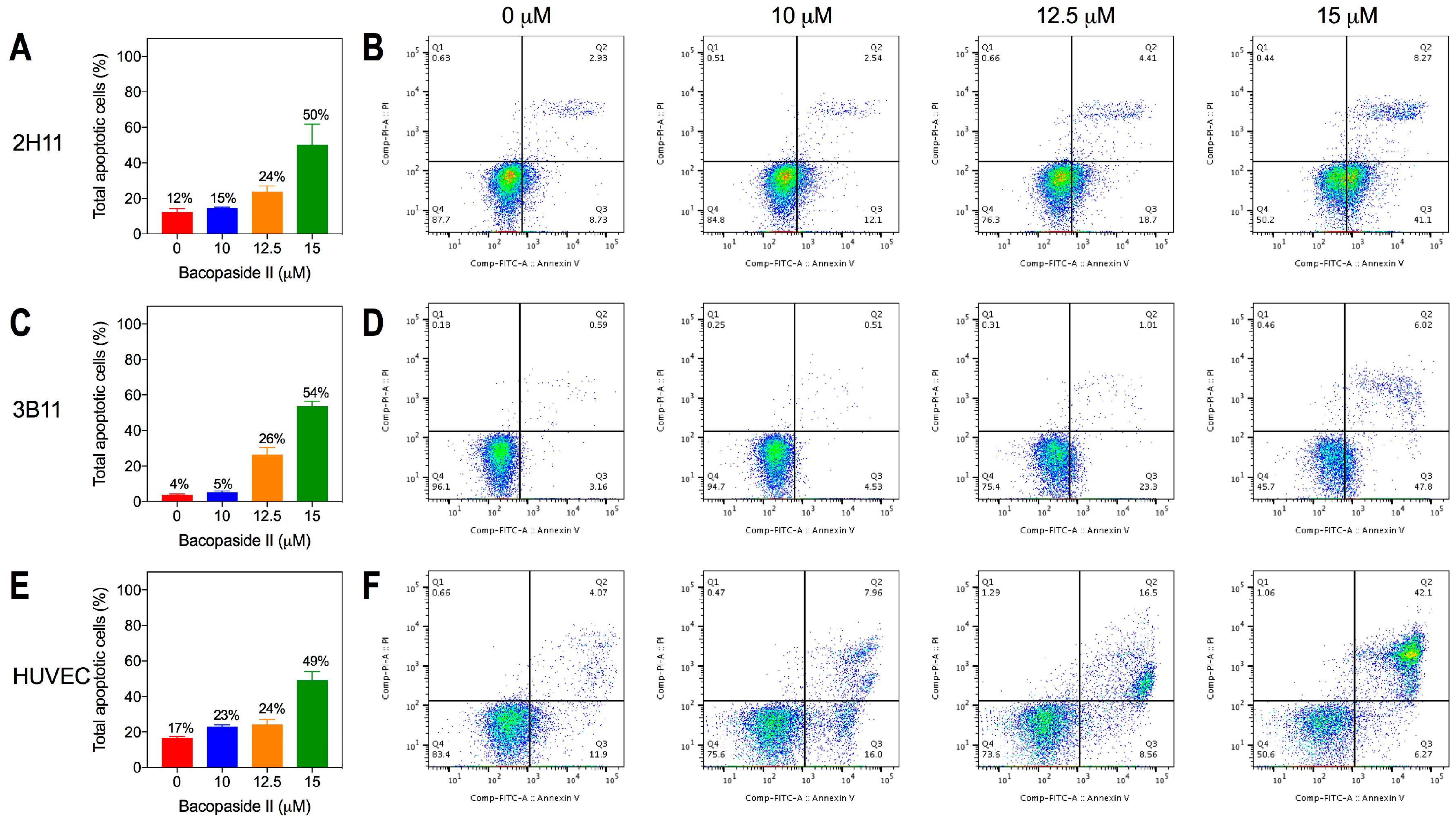
2.2. Bacopaside II Reduced Endothelial Cell Viability and Increased Apoptosis
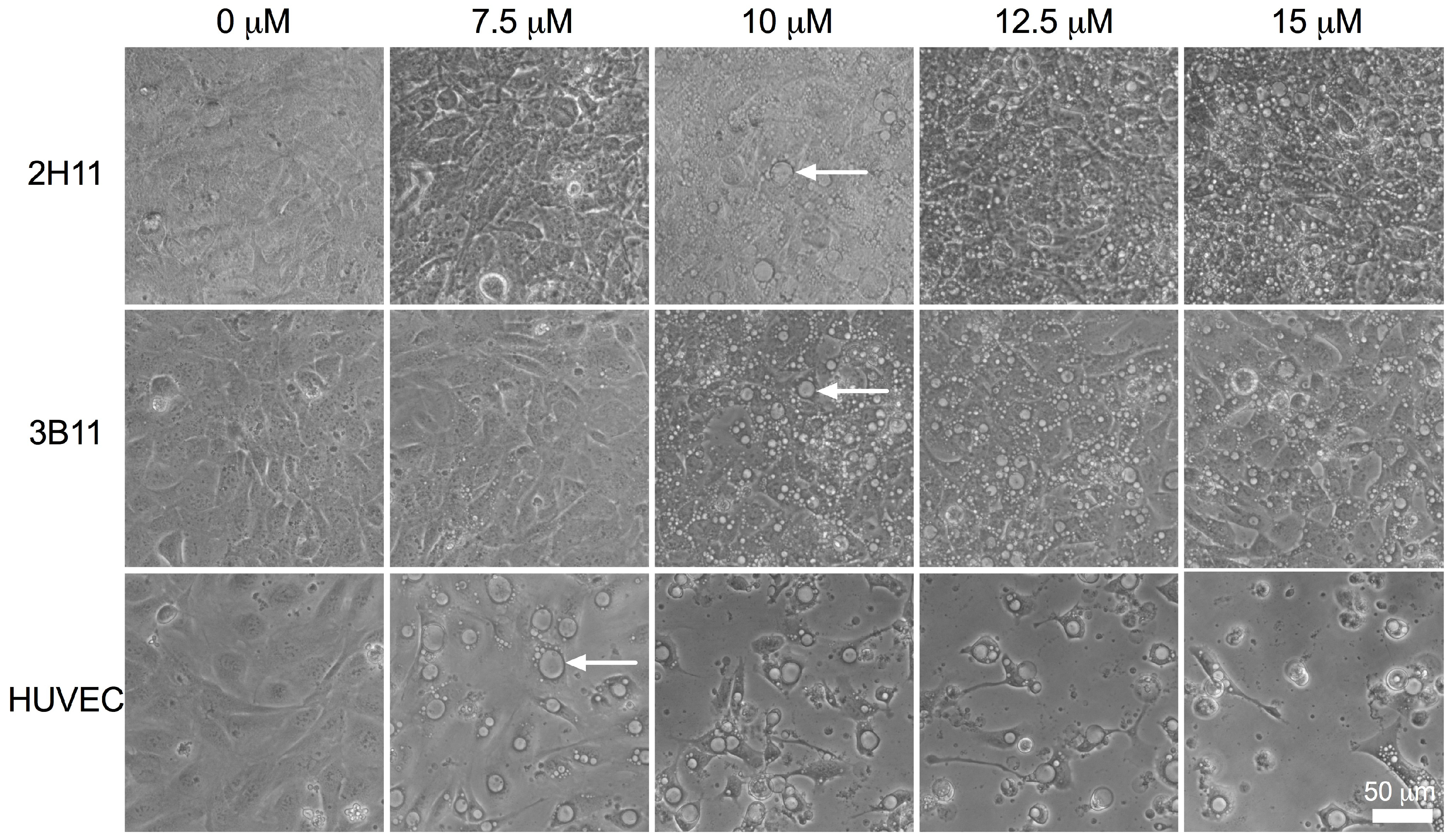
2.3. Endothelial Cell Morphology Was Altered by Bacopaside II
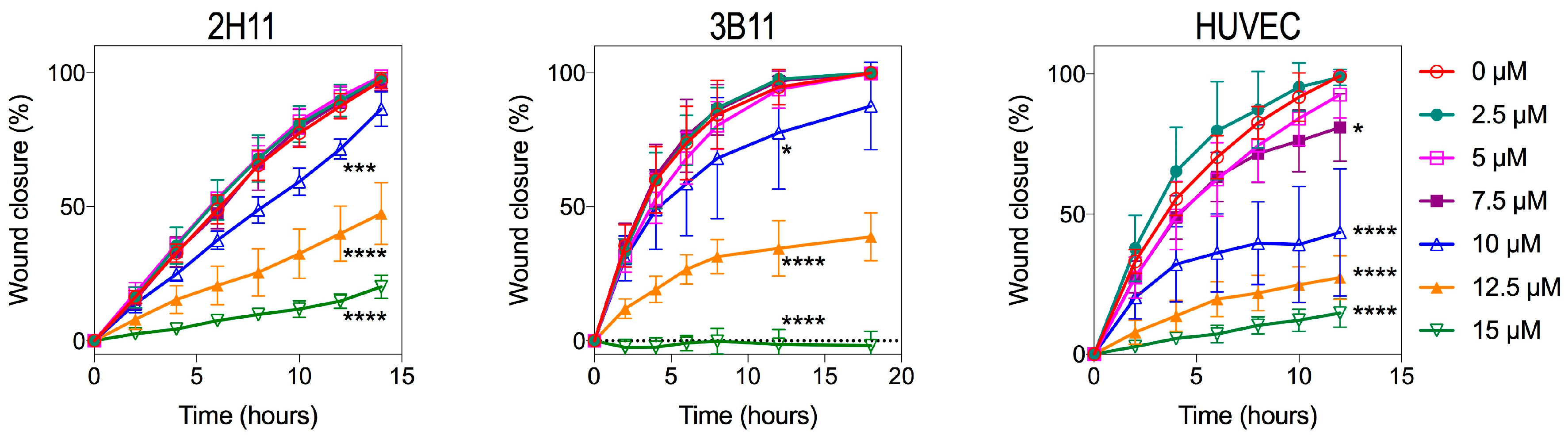
2.4. Endothelial Cell Migration Was Reduced by Bacopaside II
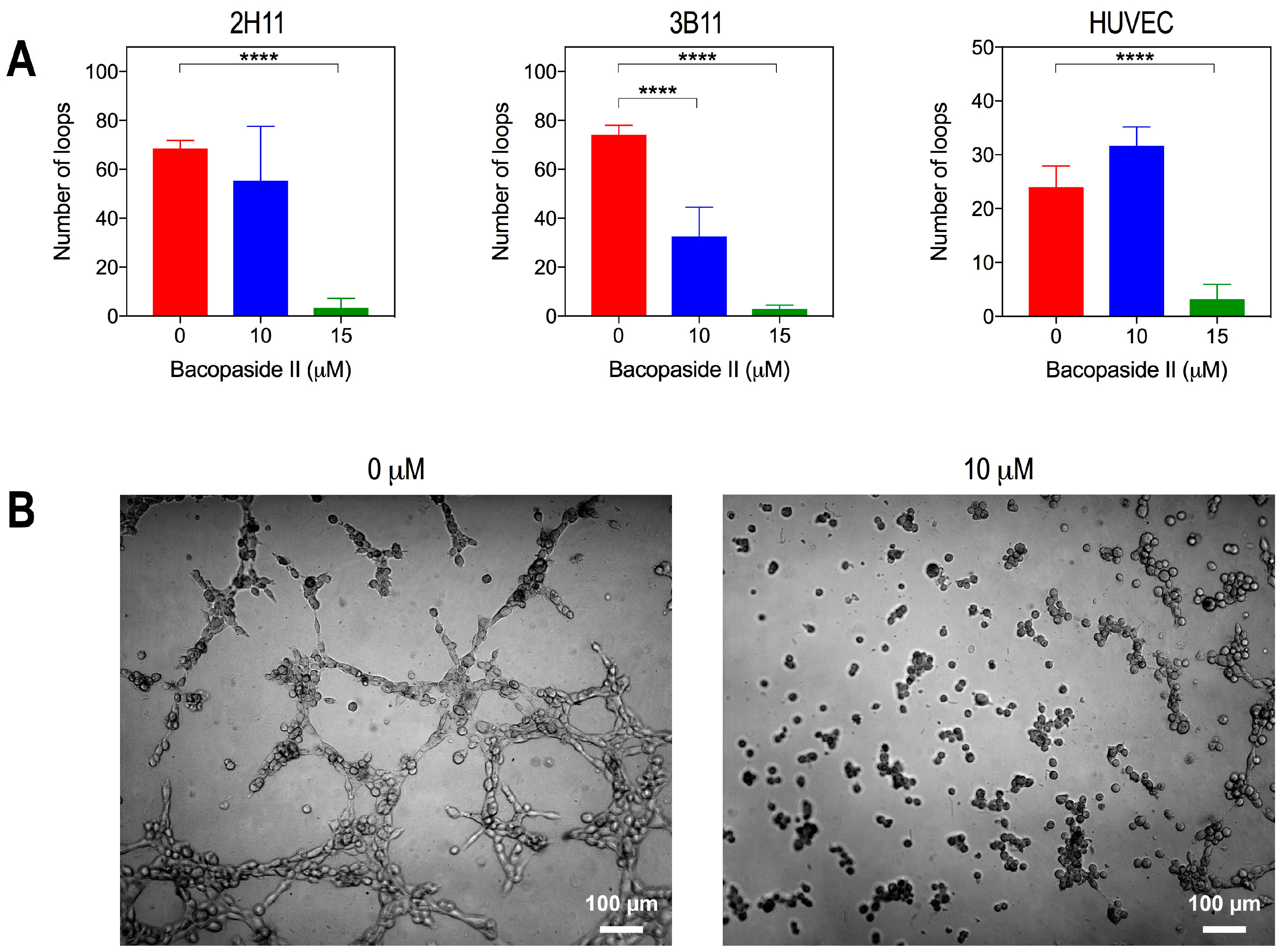
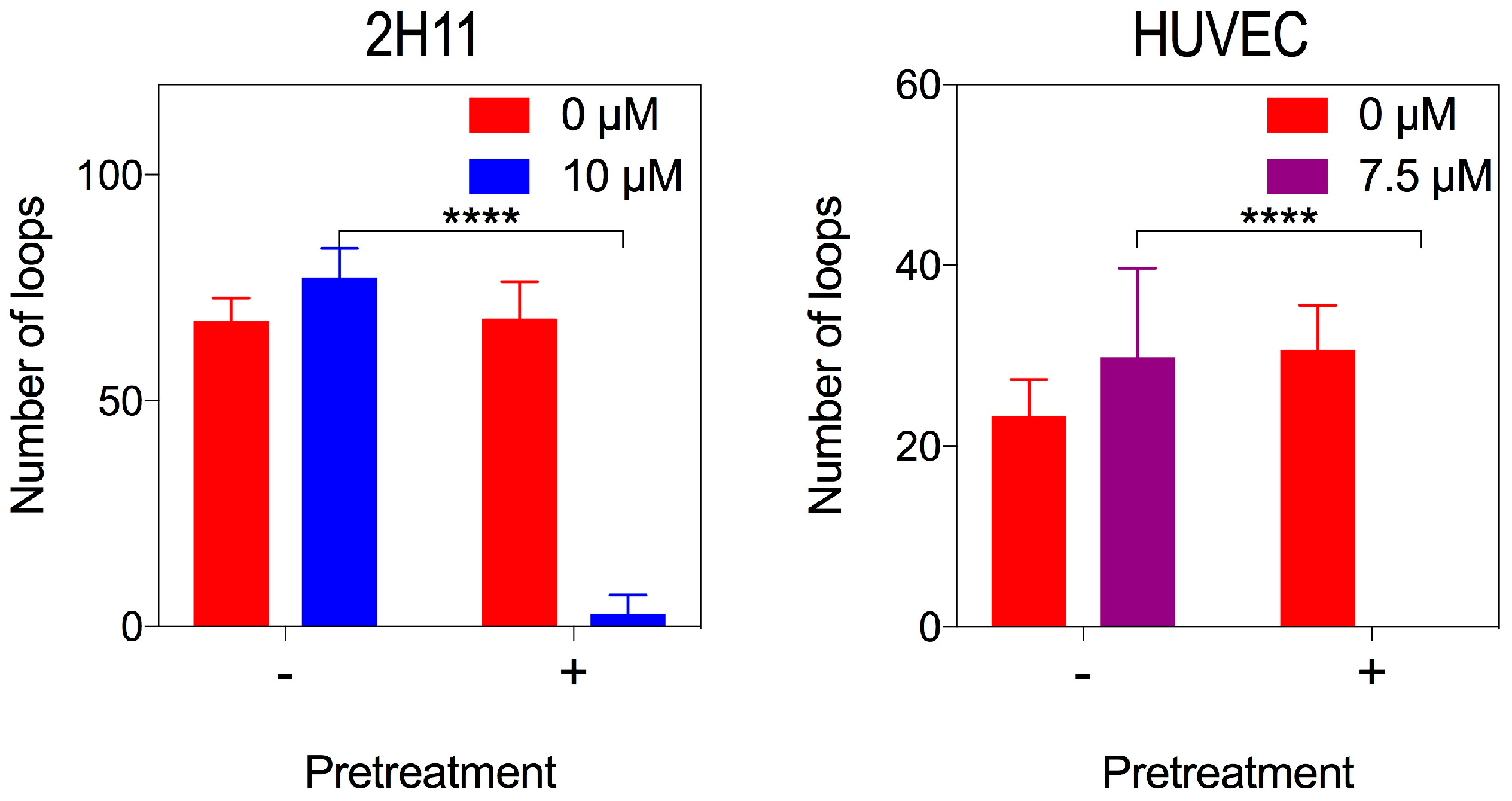
2.5. Bacopaside II Inhibited Endothelial Cell Tube Formation
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Analysis of AQP1 Expression by Western Immunoblot
4.3. MTS Viability Assay
4.4. Apoptosis Assay
4.5. Scratch Wound (Migration) Assay
4.6. Endothelial Tube Formation (Angiogenesis) Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AQP1 | Aquaporin 1 |
| MTS | (3-4,5-Dimethylthiazol-2-yl)-5(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt |
| HUVEC | Human umbilical vein endothelial cell |
| ATCC | American Type Culture Collection |
| FCS | Foetal calf serum |
| VEGF | Vascular endothelial growth factor |
References
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [PubMed]
- Rajabi, M.; Mousa, S.A. The Role of Angiogenesis in Cancer Treatment. Biomedicines 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, M.C.; Saadoun, S.; Verkman, A.S. Aquaporins and cell migration. Eur. J. Physiol. 2008, 456, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Monzani, E.; Shtil, A.A.; la Porta, C.A. The water channels, new druggable targets to combat cancer cell survival, invasiveness and metastasis. Curr. Drug Targets 2007, 8, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Dorward, H.; Yool, A.J.; Smith, E.; Townsend, A.R.; Price, T.J.; Hardingham, J.E. Role of Aquaporin 1 Signalling in Cancer Development and Progression. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Dorward, H.S.; Du, A.; Bruhn, M.A.; Wrin, J.; Pei, J.V.; Evdokiou, A.; Price, T.J.; Yool, A.J.; Hardingham, J.E. Pharmacological blockade of aquaporin-1 water channel by AqB013 restricts migration and invasiveness of colon cancer cells and prevents endothelial tube formation in vitro. J. Exp. Clin. Cancer Res. 2016, 35, 36. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Borrelli, F. Bacopa monniera, a reputed nootropic plant: An overview. Phytomedicine 2005, 12, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.V.; Kourghi, M.; de Ieso, M.L.; Campbell, E.M.; Dorward, H.S.; Hardingham, J.E.; Yool, A.J. Differential Inhibition of Water and Ion Channel Activities of Mammalian Aquaporin-1 by Two Structurally Related Bacopaside Compounds Derived from the Medicinal Plant Bacopa monnieri. Mol. Pharmacol. 2016, 90, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Giantonio, B.J.; Catalano, P.J.; Meropol, N.J.; O’Dwyer, P.J.; Mitchell, E.P.; Alberts, S.R.; Schwartz, M.A.; Benson, A.B., 3rd. Bevacizumab in combination with oxaliplatin, fluorouracil, and leucovorin (FOLFOX4) for previously treated metastatic colorectal cancer: Results from the Eastern Cooperative Oncology Group Study E3200. J. Clin. Oncol. 2007, 25, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Tebbutt, N.C.; Wilson, K.; Gebski, V.J.; Cummins, M.M.; Zannino, D.; van Hazel, G.A.; Robinson, B.; Broad, A.; Ganju, V.; Ackland, S.P.; et al. Capecitabine, bevacizumab, and mitomycin in first-line treatment of metastatic colorectal cancer: Results of the Australasian Gastrointestinal Trials Group Randomized Phase III MAX Study. J. Clin. Oncol. 2010, 28, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Vasudev, N.S.; Reynolds, A.R. Anti-angiogenic therapy for cancer: Current progress, unresolved questions and future directions. Angiogenesis 2014, 17, 471–494. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A.; van Cutsem, E.; Sobrero, A.; Siena, S.; Falcone, A.; Ychou, M.; Humblet, Y.; Bouche, O.; Mineur, L.; Barone, C.; et al. Regorafenib monotherapy for previously treated metastatic colorectal cancer (CORRECT): An international, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 2013, 381, 303–312. [Google Scholar] [CrossRef]
- Bruhn, M.A.; Townsend, A.R.; Khoon Lee, C.; Shivasami, A.; Price, T.J.; Wrin, J.; Arentz, G.; Tebbutt, N.C.; Hocking, C.; Cunningham, D.; et al. Proangiogenic tumor proteins as potential predictive or prognostic biomarkers for bevacizumab therapy in metastatic colorectal cancer. Int. J. Cancer 2014, 135, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Tomida, C.; Nagano, H.; Yamagishi, N.; Uchida, T.; Ohno, A.; Hirasaka, K.; Nikawa, T.; Teshima-Kondo, S. Regorafenib induces adaptive resistance of colorectal cancer cells via inhibition of vascular endothelial growth factor receptor. J. Med. Investig. 2017, 64, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.L.; Preston, G.M.; Spring, F.A.; Anstee, D.J.; Agre, P. Human red cell aquaporin CHIP. I. Molecular characterization of ABH and Colton blood group antigens. J. Clin. Investig. 1994, 94, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Rachumallu, R.; Bhateria, M.; Panduri, J.; Bhatta, R.S. In vitro effects of standardized extract of Bacopa monniera and its five individual active constituents on human P-glycoprotein activity. Xenobiotica 2015, 45, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Angiogenesis inhibitors: A new class of drugs. Cancer Biol. Ther. 2003, 2, S127–S133. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Kunimasa, K.; Kobayashi, T.; Sakamoto, M.; Kaji, K. Propolis suppresses tumor angiogenesis by inducing apoptosis in tube-forming endothelial cells. Biosci. Biotechnol. Biochem. 2008, 72, 2436–2440. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.; Subramanian, I.V.; Kelekar, A.; Ramakrishnan, S. Kringle 5 of human plasminogen, an angiogenesis inhibitor, induces both autophagy and apoptotic death in endothelial cells. Blood 2007, 109, 4793–4802. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.; Nguyen, T.M.; Subramanian, I.V.; Kelekar, A. Autophagy and angiogenesis inhibition. Autophagy 2007, 3, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Saltz, L.B.; Clarke, S.; Diaz-Rubio, E.; Scheithauer, W.; Figer, A.; Wong, R.; Koski, S.; Lichinitser, M.; Yang, T.S.; Rivera, F.; et al. Bevacizumab in combination with oxaliplatin-based chemotherapy as first-line therapy in metastatic colorectal cancer: A randomized phase III study. J. Clin. Oncol. 2008, 26, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.; Lang, I.; Marcuello, E.; Lorusso, V.; Ocvirk, J.; Shin, D.B.; Jonker, D.; Osborne, S.; Andre, N.; Waterkamp, D.; et al. Bevacizumab plus capecitabine versus capecitabine alone in elderly patients with previously untreated metastatic colorectal cancer (AVEX): An open-label, randomised phase 3 trial. Lancet Oncol. 2013, 14, 1077–1085. [Google Scholar] [CrossRef]
- Abdollahi, A.; Folkman, J. Evading tumor evasion: Current concepts and perspectives of anti-angiogenic cancer therapy. Drug Resist. Updates 2010, 13, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Kerbel, R.S. Tumor angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.; Smith, B.L.; Preston, G.M. ABH and Colton blood group antigens on aquaporin-1, the human red cell water channel protein. Transfus. Clin. Biol. 1995, 2, 303–308. [Google Scholar] [CrossRef]
- King, L.S.; Choi, M.; Fernandez, P.C.; Cartron, J.P.; Agre, P. Defective urinary concentrating ability due to a complete deficiency of aquaporin-1. N. Engl. J. Med. 2001, 345, 175–179. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palethorpe, H.M.; Tomita, Y.; Smith, E.; Pei, J.V.; Townsend, A.R.; Price, T.J.; Young, J.P.; Yool, A.J.; Hardingham, J.E. The Aquaporin 1 Inhibitor Bacopaside II Reduces Endothelial Cell Migration and Tubulogenesis and Induces Apoptosis. Int. J. Mol. Sci. 2018, 19, 653. https://doi.org/10.3390/ijms19030653
Palethorpe HM, Tomita Y, Smith E, Pei JV, Townsend AR, Price TJ, Young JP, Yool AJ, Hardingham JE. The Aquaporin 1 Inhibitor Bacopaside II Reduces Endothelial Cell Migration and Tubulogenesis and Induces Apoptosis. International Journal of Molecular Sciences. 2018; 19(3):653. https://doi.org/10.3390/ijms19030653
Chicago/Turabian StylePalethorpe, Helen M., Yoko Tomita, Eric Smith, Jinxin V. Pei, Amanda R. Townsend, Timothy J. Price, Joanne P. Young, Andrea J. Yool, and Jennifer E. Hardingham. 2018. "The Aquaporin 1 Inhibitor Bacopaside II Reduces Endothelial Cell Migration and Tubulogenesis and Induces Apoptosis" International Journal of Molecular Sciences 19, no. 3: 653. https://doi.org/10.3390/ijms19030653
APA StylePalethorpe, H. M., Tomita, Y., Smith, E., Pei, J. V., Townsend, A. R., Price, T. J., Young, J. P., Yool, A. J., & Hardingham, J. E. (2018). The Aquaporin 1 Inhibitor Bacopaside II Reduces Endothelial Cell Migration and Tubulogenesis and Induces Apoptosis. International Journal of Molecular Sciences, 19(3), 653. https://doi.org/10.3390/ijms19030653