Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.)

Abstract
:1. Introduction
2. Results
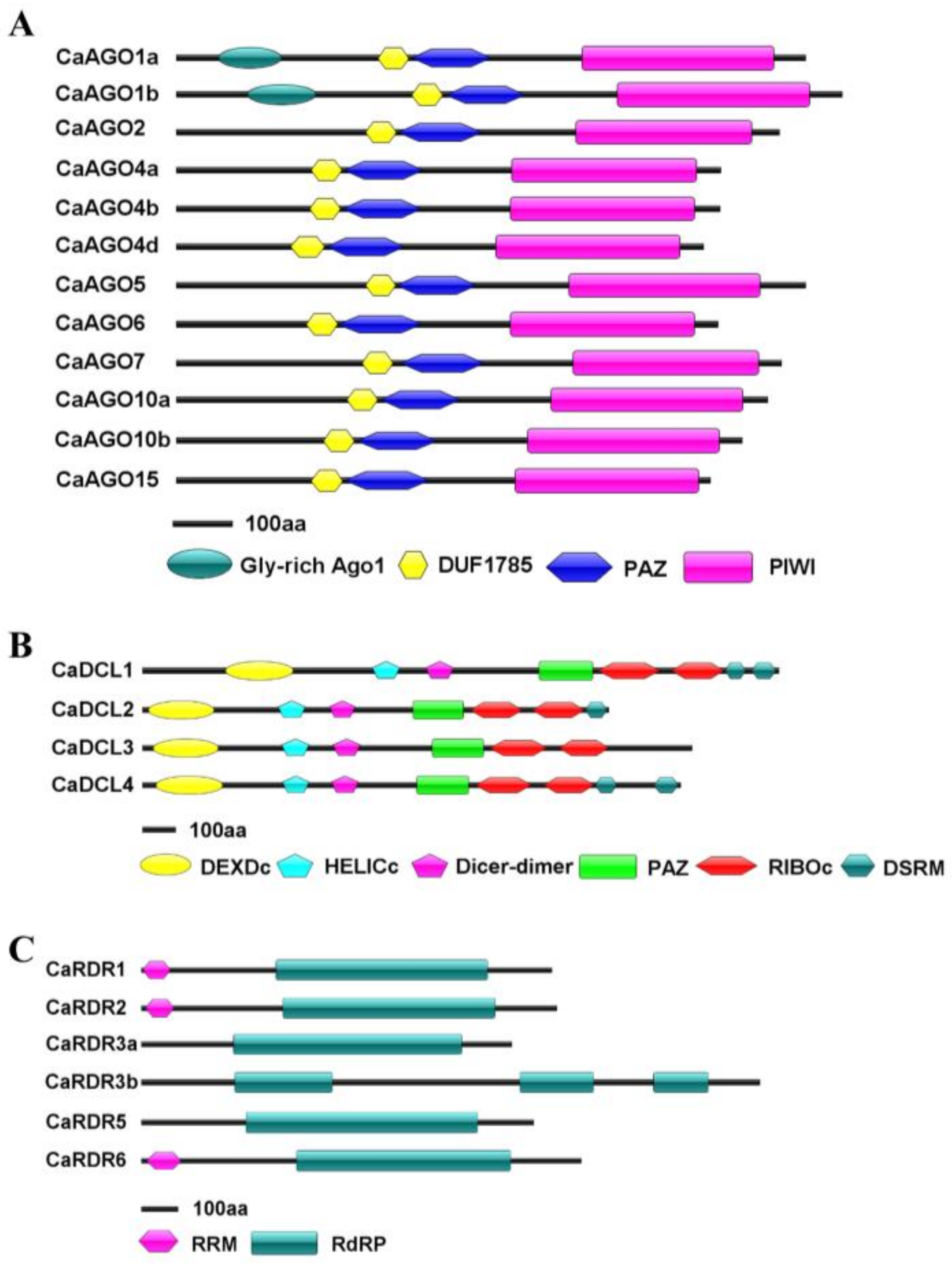
2.1. Identification and Structural Analysis of CaAGO, CaDCL and CaRDR Genes
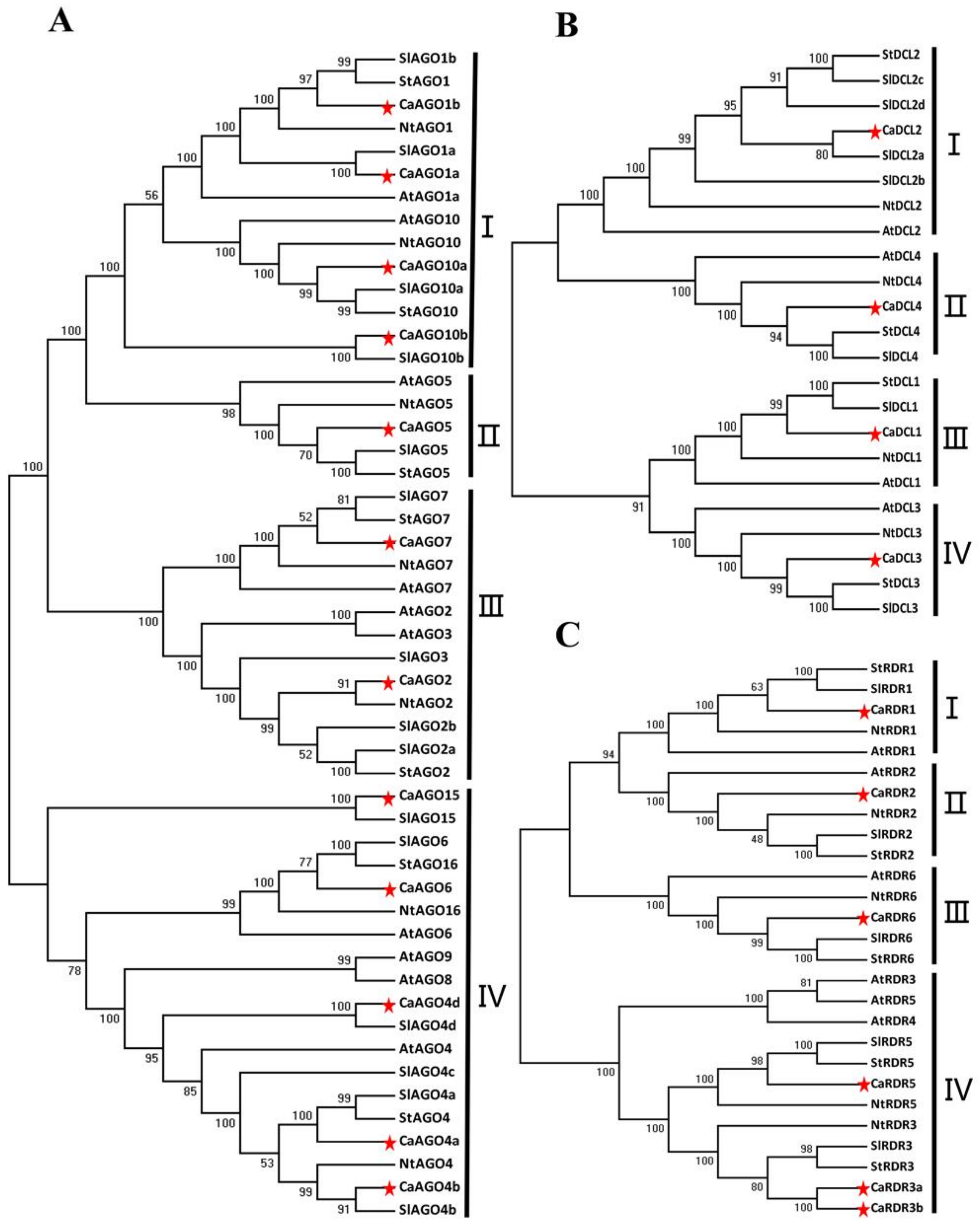
2.2. Phylogenetic Analysis of CaAGO, CaDCL and CaRDR Genes
2.3. Expression Pattern of CaAGOs, CaDCLs and CaRDRs in Various Organs
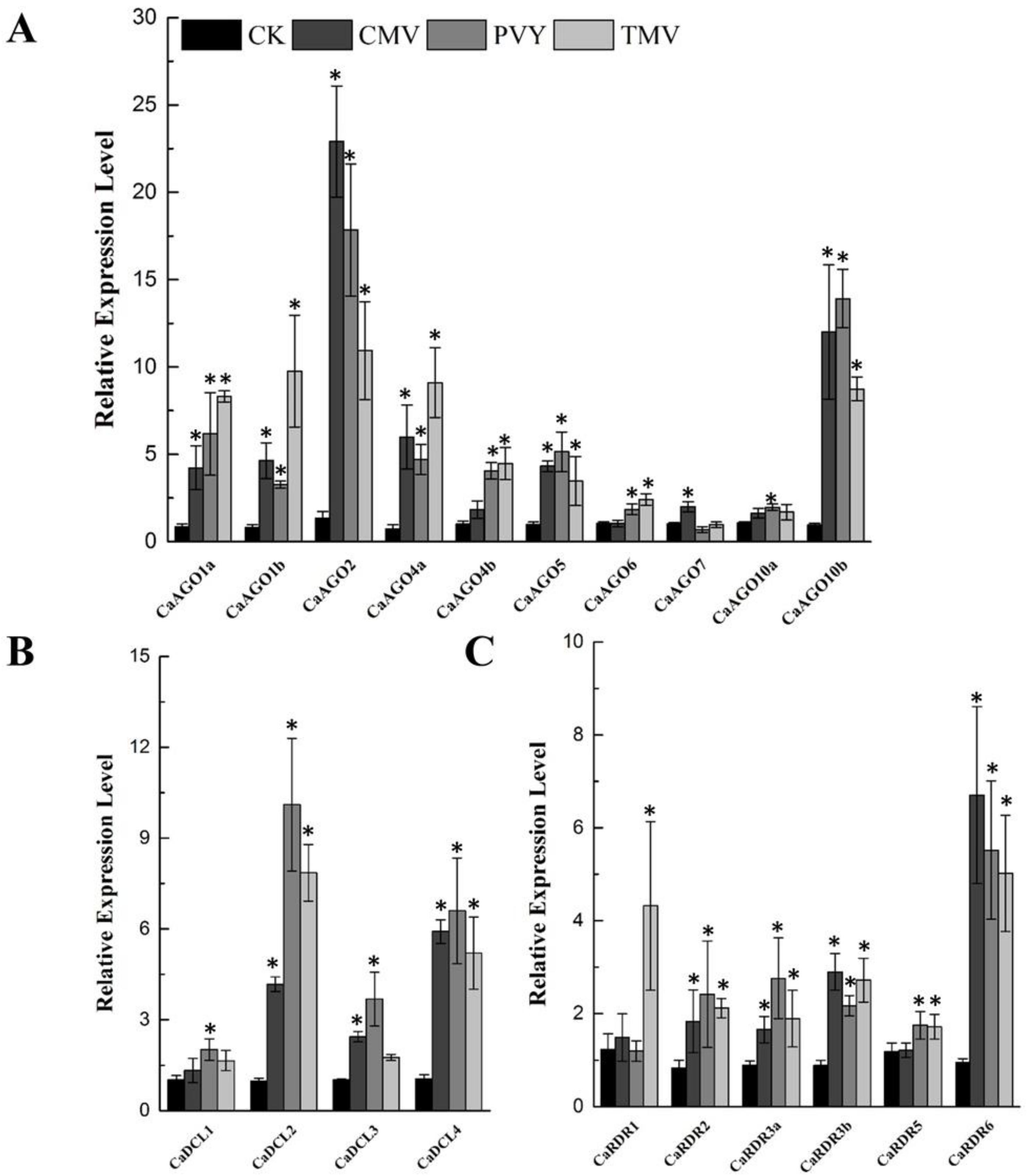
2.4. Biotic Stress Induces Expression of CaAGO, CaDCL and CaRDR Genes
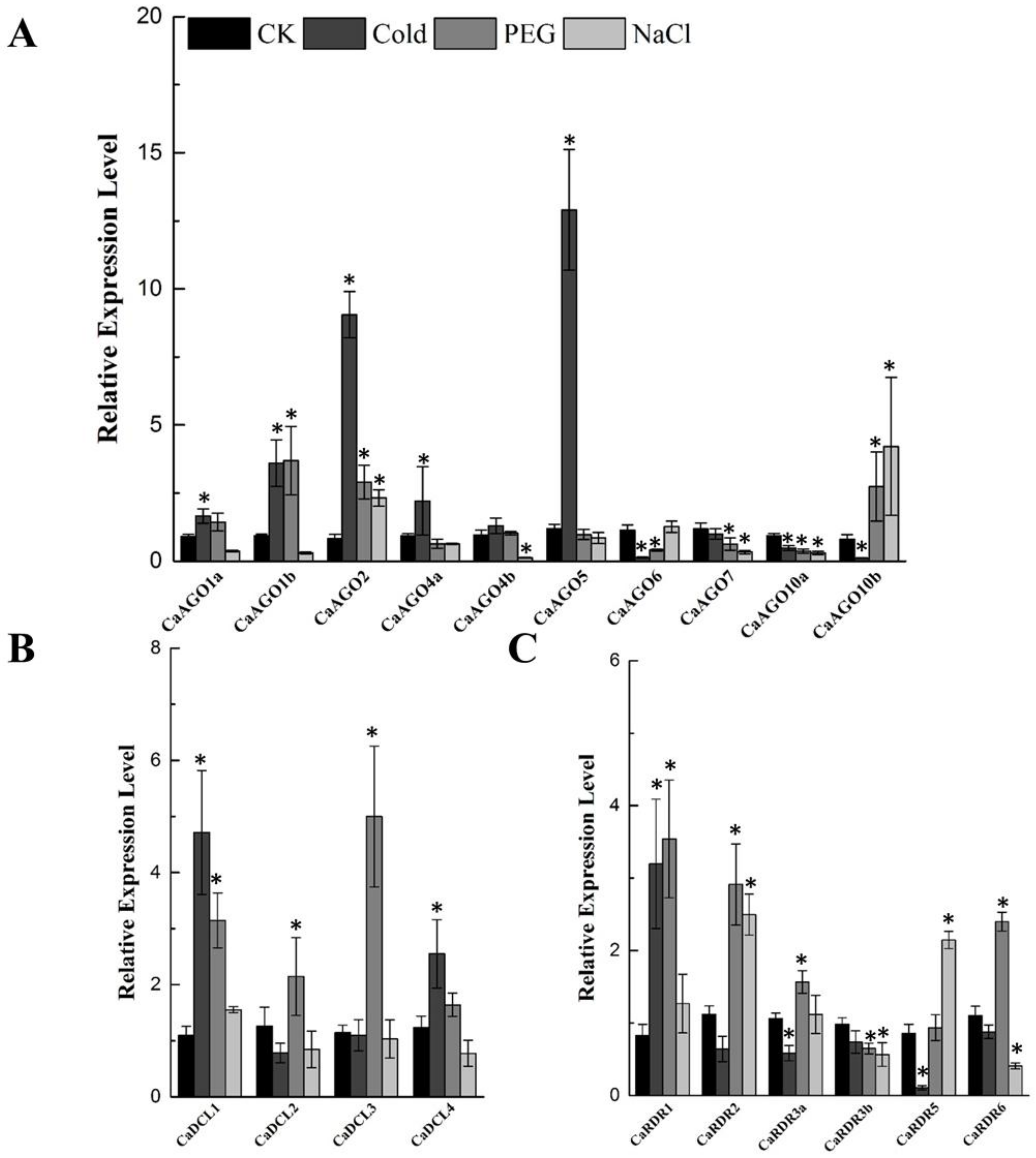
2.5. Abiotic Stress Induces CaAGO, CaDCL and CaRDR Expression
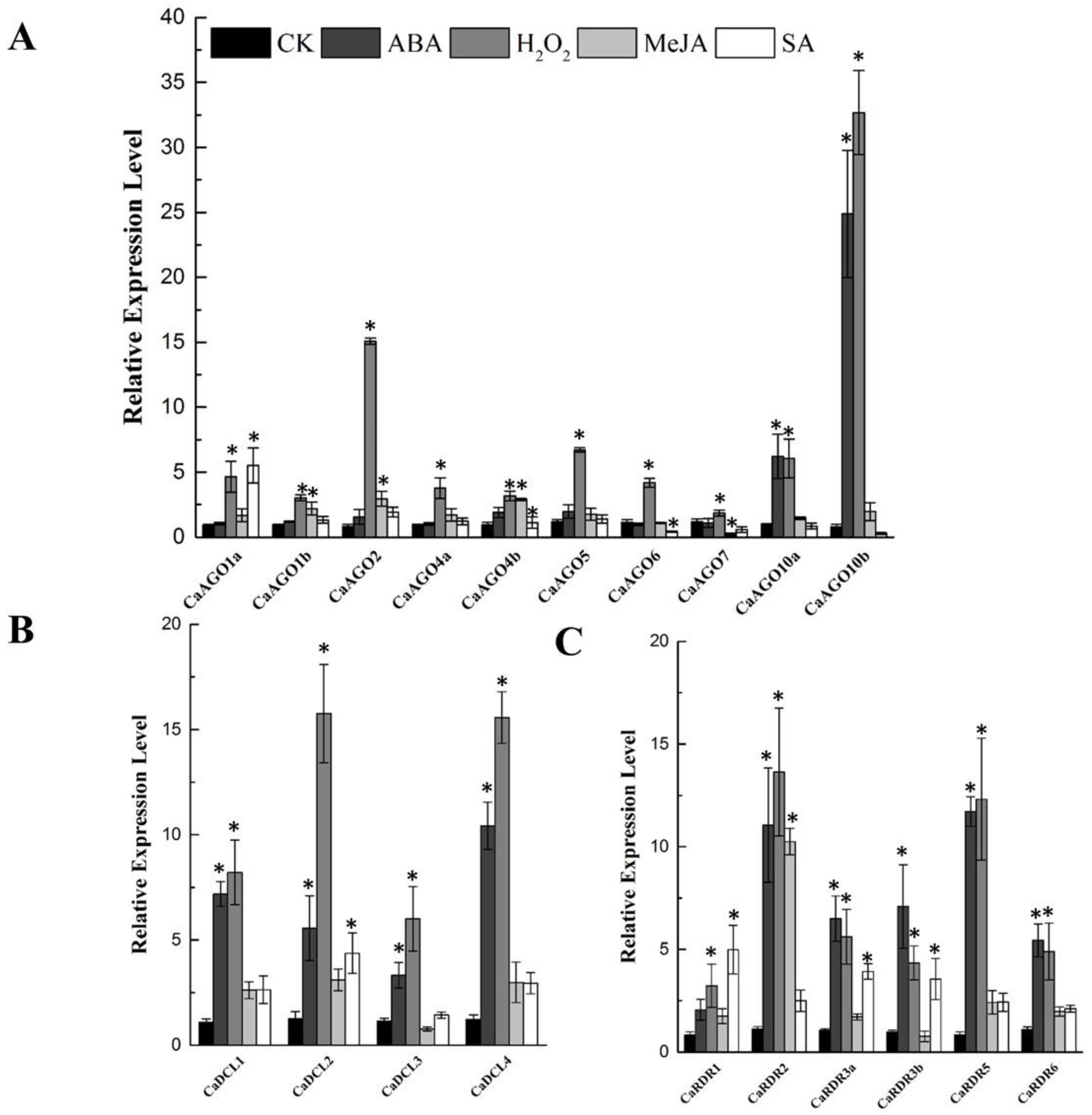
2.6. CaAGOs, CaDCLs and CaRDRs Are Responsive to Phytohormones and H2O2
3. Discussion
3.1. Argonaute (AGO) Proteins in Pepper
3.2. Dicer-Like (DCL) Proteins in Pepper
3.3. RNA-Dependent RNA Polymerase (RDR) Proteins in Pepper
4. Materials and Methods
4.1. Identification of Putative Pepper DCL, AGO, and RDR Genes
4.2. Phylogenetic Analysis
4.3. Plant and Treatment
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AGO | Argonaute Protein |
| ABA | Abscisic Acid |
| CMV | Cucumber Mosaic Virus |
| DCL | Dicer-Like Protein |
| MeJA | Methyl Jasmonate |
| MW | Molecular Weight |
| NaCl | Sodium Chloride |
| PEG | Polyethylene Glycol |
| PVY | Potato Virus Y |
| RDR | RNA-Dependent RNA Polymerase protein |
| RISCs | RNA-induced Silencing Complexes |
| SA | Salicylic Acid |
| ssRNAs | Single-Stranded RNAs |
| TMV | Tobacco Mosaic Virus |
References
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.J.; Carrington, J.C. Specialization and evolution of endogenous small RNA pathways. Nat. Rev. Genet. 2007, 8, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Post-transcriptional small RNA pathways in plants: Mechanisms and regulations. Genes Dev. 2006, 20, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Carmell, M.A.; Hannon, G.J. RNase III enzymes and the initiation of gene silencing. Nat. Struct. Mol. Biol. 2004, 11, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Fusaro, A.F.; Smith, N.A.; Curtin, S.J.; Watson, J.M.; Finnegan, E.J.; Waterhouse, P.M. The evolution and diversification of dicers in plants. FEBS Lett. 2006, 580, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.; Kirkpatrick, J.P.; Eckhardt, S.; Reuter, M.; Rocha, E.A.; Andradenavarro, M.A.; Sehr, P.; Pillai, R.S.; Carlomagno, T. Recognition of 2′-o-methylated 3′-end of piRNA by the PAZ domain of a PIWI protein. Structure 2011, 19, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M.; Krczal, G. Nomenclature and functions of RNA-directed rna polymerases. Trends Plant Sci. 2006, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Yang, G.S.; Chen, W.T.; Mao, Z.C.; Kang, H.X.; Chen, G.H.; Yang, Y.H.; Xie, B.Y. Genome-wide identification of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 2012, 501, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Arora, R.; Lama, T.; Nijhawan, A.; Khurana, J.P.; Tyagi, A.K.; Kapoor, S. Genome-wide identification, organization and phylogenetic analysis of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analysis during reproductive development and stress in rice. BMC Genom. 2008, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Plant Argonautes. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Liang, D.; Wu, J.; Zhan, M.; Yang, F.; Xu, W.; Zhu, S.; Shi, J. Genome-wide identification of the Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in cucumber (Cucumis sativus L.). J. Plant Growth Regul. 2016, 35, 135–150. [Google Scholar] [CrossRef]
- Cao, J.Y.; Xu, Y.P.; Li, W.; Li, S.S.; Rahman, H.; Cai, X.Z. Genome-wide identification of Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in Brassica species and functional analyses of their Arabidopsis homologs in resistance to Sclerotinia sclerotiorum. Front. Plant Sci. 2016, 7, 1614. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, W.; Zhong, Z.; Chen, X.; Wang, A.; Wang, Z. Genome-wide analysis of RNA-interference pathway in Brassica napus, and the expression profile of BnAGOs in response to Sclerotinia sclerotiorum infection. Eur. J. Plant Pathol. 2016, 146, 1–15. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, K.; Wang, J.; Chen, X.; Chen, Z.; Cai, R.; Xiang, Y. Comprehensive analysis of Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in grapevine (vitis vinifera). J. Plant Growth Regul. 2015, 34, 108–121. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (lib.) de bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Erental, A.; Dickman, M.B.; Yarden, O. Sclerotial development in sclerotinia sclerotiorum: Awakening molecular analysis of a “dormant” structure. Fungal Biol. Rev. 2008, 22, 6–16. [Google Scholar] [CrossRef]
- Qin, L.; Mo, N.; Zhang, Y.; Muhammad, T.; Zhao, G.; Zhang, Y.; Liang, Y. CaRDR1, an RNA-dependent RNA polymerase plays a positive role in pepper resistance against TMV. Front. Plant Sci. 2017, 8, 1068. [Google Scholar] [CrossRef] [PubMed]
- Do, H.M.; Lee, S.C.; Jung, H.W.; Sohn, K.H.; Hwang, B.K. Differential expression and in situ localization of a pepper defensin (CADEF1) gene in response to pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Plant Sci. 2004, 166, 1297–1305. [Google Scholar]
- Yu, B.K.; Lee, J.H.; Shin, S.J.; Hwang, E.W.; Kwon, H.B. Molecular characterization of cold stress-related transcription factors, CAEREBP-C1, -C2, -C3, and CaWRKY1a from Capsicum annuum L. J. Plant Biol. 2013, 56, 106–114. [Google Scholar] [CrossRef]
- Lcvan, L.; Eavan, S. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Evolutionary connection between the catalytic subunits of DNA-dependent RNA polymerases and eukaryotic RNA-dependent RNA polymerases and the origin of RNA polymerases. BMC Struct. Biol. 2003, 3, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.S.; Roe, S.M.; Barford, D. Structural insights into mRNA recognition from a PIWI domain-siRNA guide complex. Nature 2005, 434, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Fagard, M.; Boutet, S.; Morel, J.B.; Bellini, C.; Vaucheret, H. AGO1, QDE-2, and RDE-1 are related proteins required for post-transcriptional gene silencing in plants, quelling in fungi, and RNA interference in animals. Proc. Natl. Acad. Sci. USA 2000, 97, 11650–11654. [Google Scholar] [CrossRef] [PubMed]
- Bohmert, K.; Camus, I.; Bellini, C.; Bouchez, D.; Caboche, M.; Benning, C. AGO1 defines a novel locus of Arabidopsis controlling leaf development. EMBO J. 1998, 17, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hu, F.; Wang, R.; Zhou, X.; Sze, S.H.; Liou, L.W.; Barefoot, A.; Dickman, M.; Zhang, X. Arabidopsis Argonaute10 specifically sequesters miR166/165 to regulate shoot apical meristem development. Cell 2011, 145, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Liu, X.; Yan, J.; Wang, W.; Yumul, R.E.; Yun, J.K.; Dinh, T.T.; Liu, J.; Cui, X.; Zheng, B. Argonaute10 and Argonaute1 regulate the termination of floral stem cells through two micrornas in Arabidopsis. PLoS Genet. 2011, 7, e1001358. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Mallory, A.C.; Bartel, D.P. AGO1 homeostasis entails coexpression of miR168 and AGO1 and preferential stabilization of miR168 by AGO1. Mol. Cell 2006, 22, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ji, L.; Le, B.H.; Zhai, J.; Chen, J.; Luscher, E.; Gao, L.; Liu, C.; Cao, X.; Mo, B. Argonaute10 promotes the degradation of miR165/6 through the SDN1 and SDN2 exonucleases in Arabidopsis. PLoS Biol. 2017, 15, e2001272. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.B.; Provost, P.; Ekwall, K.; Hobman, T.C. Ago1 and Dcr1, two core components of the RNA interference pathway, functionally diverge from Rdp1 in regulating cell cycle events in Schizosaccharomyces pombe. Mol. Biol. Cell 2004, 15, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Fernándeznohales, P.; Domenech, M.J.; Martínez de Alba, A.E.; Micol, J.L.; Ponce, M.R.; Madueño, F. AGO1 controls arabidopsis inflorescence architecture possibly by regulating TFL1 expression. Ann. Bot. 2014, 114, 1471–1481. [Google Scholar]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Irish, V.F. The flowering of Arabidopsis flower development. Plant J. Cell Mol. Biol. 2010, 61, 1014–1028. [Google Scholar]
- Byrne, M.E.; Barley, R.; Curtis, M.; Arroyo, J.M.; Dunham, M.; Hudson, A.; Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell functionin Arabidopsis. Nature 2000, 408, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Semiarti, E.; Ueno, Y.; Tsukaya, H.; Iwakawa, H.; Machida, C.; Machida, Y. The Asymmetric leaves2 gene of Arabidopsis thaliana regulates formation of a symmetric lamina, establishment of venation and repression of meristem-related homeobox genes in leaves. Development 2001, 128, 1771–1783. [Google Scholar] [PubMed]
- Li, Y.; Huang, W.; Hua, W.; Cai, R.; Xu, Y.; Hai, H. Characterizations of a hypomorphic Argonaute1 mutant reveal novel ago1 functions in arabidopsis lateral organ development. Plant Mol. Biol. 2006, 61, 63–78. [Google Scholar]
- Yang, Y.; Jun, Z.; Yi-Dan, O.; Jialing, Y. The integrative expression and co-expression analysis of the ago gene family in rice. Gene 2013, 528, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.J.; Lewsey, M.G.; Patel, K.; Westwood, J.; Heimstädt, S.; Carr, J.P.; Baulcombe, D.C. An antiviral defense role of AGO2 in plants. PLoS ONE 2011, 6, e14639. [Google Scholar] [CrossRef] [PubMed]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.S.; Perry, K.L.; Moffett, P. Argonaute2 mediates RNA-silencing antiviral defenses against Potato virus X in Arabidopsis. Plant Physiol. 2011, 156, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, H.B.; Alvarado, V.Y.; Vegaarreguin, J.C.; Ciomperlik, J.; Odokonyero, D.; Brosseau, C.; Jaubert, M.; Zamora, A.; Moffett, P. Identification of an Argonaute for antiviral RNA silencing in Nicotiana Benthamiana. Plant Physiol. 2011, 156, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Singh, J.; Li, D.; Qu, F. Temperature-dependent survival of turnip crinkle virus-infected Arabidopsis plants relies on an RNA silencing-based defense that requires DCL2, AGO2, and HEN1. J. Virol. 2012, 86, 6847–6854. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, C.; Moffett, P. Functional and genetic analysis identify a role for Arabidopsis Argonaute5 in antiviral RNA silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, H.; Gao, S.; Wang, W.C.; Katiyaragarwal, S.; Huang, H.D.; Raikhel, N.; Jin, H. Arabidopsis Argonaute 2 regulates innate immunity via miRNA393∗-mediated silencing of a Golgi-localized SNARE gene, MEMB12. Mol. Cell 2011, 42, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Odokonyero, D.; Mendoza, M.R.; Alvarado, V.Y.; Zhang, J.; Wang, X.; Scholthof, H.B. Transgenic down-regulation of ARGONAUTE2 expression in Nicotiana benthamiana interferes with several layers of antiviral defenses. Virology 2015, 486, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ludman, M.; Burgyán, J.; Fátyol, K. Crispr/Cas9 mediated inactivation of argonaute 2 reveals its differential involvement in antiviral responses. Sci. Rep. 2017, 7, 1010. [Google Scholar] [CrossRef] [PubMed]
- Diermann, N.; Matoušek, J.; Junge, M.; Riesner, D.; Steger, G. Characterization of plant miRNAs and small RNAs derived from Potato Spindle Tuber Viroid (PSTVD) in infected tomato. Biol. Chem. 2010, 391, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Bazzini, A.A.; Manacorda, C.A.; Tohge, T.; Conti, G.; Rodriguez, M.C.; Nunesnesi, A.; Villanueva, S.; Fernie, A.R.; Carrari, F.; Asurmendi, S. Metabolic and miRNA profiling of TMV infected plants reveals biphasic temporal changes. PLoS ONE 2011, 6, e28466. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.B.; Godon, C.; Mourrain, P.; Beclin, C.; Boutet, S.; Feuerbach, F.; Proux, F.; Vaucheret, H. Fertile hypomorphic argonaute (ago1) mutants impaired in post-transcriptional gene silencing and virus resistance. Plant Cell 2002, 14, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xian, Z.; Huang, W.; Li, Z. Evidence for the biological function of miR403 in tomato development. Sci. Horticult. 2015, 197, 619–626. [Google Scholar] [CrossRef]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Bao, D.; Ganbaatar, O.; Cui, X.; Yu, R.; Bao, W.; Falk, B.W.; Wuriyanghan, H. Downregulation of genes coding for core RNAi components and disease resistance proteins via corresponding microRNAs might be correlated with successful SMV infection in soybean. Mol. Plant Pathol. 2018, 19, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Liu, J.; Takatoshi, K.; Jongchan, W.; Tolulope, O.; Markus, H.; Thomas, T.; Nam-Hai, C.; Wang, X.J. Deep sequencing of small RNAs specifically associated with Arabidopsis AGO1 and AGO4 uncovers new AGO functions. Plant J. 2011, 67, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, K.R.; Mirlohi, S.; Khalaji, F.; Fakhari, Z.; Shiran, B.; Fallahi, H.; Rafiei, F.; Budak, H.; Ebrahimie, E. Differential expression of seven conserved microRNAs in response to abiotic stress and their regulatory network in Helianthus annuus. Front. Plant Sci. 2015, 6, 741. [Google Scholar]
- Hirayama, T.; Shinozaki, K. Perception and transduction of abscisic acid signals: Keys to the function of the versatile plant hormone ABA. Trends Plant Sci. 2007, 12, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Zelicourt, A.D.; Colcombet, J.; Hirt, H. The role of MAPK modules and ABA during abiotic stress signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Johansen, L.K.; Gustafson, A.M.; Kasschau, K.D.; Lellis, A.D.; Zilberman, D.; Jacobsen, S.E.; Carrington, J.C. Genetic and functional diversification of small RNA pathways in plants. PLoS Biol. 2004, 2, E104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Allen, E.; Wilken, A.; Carrington, J.C. Dicer-like 4 functions in trans-acting small interfering RNA biogenesis and vegetative phase change in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 12984–12989. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Denli, A.M.; Hannon, G.J. Biochemical specialization within Arabidopsis RNA silencing pathways. Mol. Cell 2005, 19, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Watanabe, Y. Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc. Natl. Acad. Sci. USA 2004, 101, 12753–12758. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Hong, L.; Fitzpatrick, K.E.; Amasino, R.M. Dicer-like 1 and Dicer-like 3 redundantly act to promote flowering via repression of flowering locus c in Arabidopsis thaliana. Genetics 2007, 176, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Liao, Y.W.; Liu, Y.R.; Liang, J.Y.; Wang, W.P.; Zhou, J.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q.; Shi, K. The relationship between the plant-encoded RNA-dependent RNA polymerase 1 and alternative oxidase in tomato basal defense against tobacco mosaic virus. Planta 2015, 241, 641–650. [Google Scholar] [CrossRef]
- Yu, D.; Fan, B.; Macfarlane, S.A.; Chen, Z. Analysis of the involvement of an inducible Arabidopsis RNA-dependent RNA polymerase in antiviral defense. Mol. Plant-Microbe Interact. 2003, 16, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Curaba, J.; Chen, X. Biochemical activities of Arabidopsis RNA-dependent RNA polymerase 6. J. Biol. Chem. 2008, 283, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Vaistij, F.E.; Jones, L. Compromised virus-induced gene silencing in RDR6-deficient plants. Plant Physiol. 2009, 149, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wu, Q.F.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.M.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, M.; Gao, Z.; Zhu, C.; Guo, X. Isolation of a novel RNA-dependent RNA polymerase 6 from Nicotiana glutinosa, NgRDR6, and analysis of its response to biotic and abiotic stresses. Mol. Biol. Rep. 2011, 38, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, K.D.; Fahlgren, N.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Carrington, J.C. Genome-wide profiling and analysis of Arabidopsis siRNAs. PLoS Biol. 2007, 5, e57. [Google Scholar] [CrossRef] [PubMed]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral genome methylation as an epigenetic defense against geminiviruses. J. Virol. 2008, 82, 8997–9007. [Google Scholar] [CrossRef] [PubMed]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.; Scott, J.W.; Edwards, J.D.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-class RNA-dependent RNA polymerases. PLoS Genet. 2013, 9, e1003399. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega 5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Hunter, L.J.R.; Westwood, J.H.; Heath, G.; Macaulay, K.; Smith, A.G.; MacFarlane, S.A.; Palukaitis, P.; Carr, J.P. Regulation of RNA-dependent RNA polymerase 1 and isochorismate synthase gene expression in Arabidopsis. PLoS ONE 2013, 8, e66530. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Yang, S.; Yan, Y.; Xiao, Z.; Cheng, J.; Wu, J.; Qiu, A.; Yan, L.; Mou, S.; Guan, D. CaWRKY6 transcriptionally activates CaWRKY40, regulates ralstonia solanacearum resistance, and confers high-temperature and high-humidity tolerance in pepper. J. Exp. Bot. 2015, 66, 3163–3174. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Chr | Range | CDS (bp) | Protein (aa) | Mw (kD) |
|---|---|---|---|---|---|---|
| CaAGO1a | Capana06g000835 | 6 | 13,219,325–13,229,295 | 3156 | 1051 | 116.40 |
| CaAGO1b | Capana03g001538 | 3 | 28,791,304–28,798,662 | 3339 | 1112 | 122.52 |
| CaAGO2 | Capana02g001299 | 2 | 119,455,171–119,460,059 | 3027 | 1008 | 112.42 |
| CaAGO4a | Capana01g001805 | 1 | 70,676,331–70,686,308 | 2733 | 910 | 102.02 |
| CaAGO4b | Capana06g000702 | 6 | 11,153,712–11,162,578 | 2730 | 909 | 101.33 |
| CaAGO4d | Capana08g001169 | 8 | 125,940,932–125,950,280 | 2646 | 881 | 98.44 |
| CaAGO5 | Capana06g000572 | 6 | 8,647,138–8,654,022 | 3048 | 1015 | 111.73 |
| CaAGO6 | Capana07g001363 | 7 | 175,512,846–175,528,611 | 2718 | 905 | 101.76 |
| CaAGO7 | Capana01g002131 | 1 | 114,240,252–114,244,871 | 3036 | 1011 | 115.15 |
| CaAGO10a | Capana03g004637 | 3 | 261,275,238–261,283,423 | 2967 | 988 | 110.83 |
| CaAGO10b | Capana09g000331 | 9 | 10,489,518–10,495,890 | 2844 | 947 | 107.02 |
| CaAGO15 | Capana03g001292 | 3 | 22,639,388–22,645,377 | 2682 | 893 | 100.94 |
| CaDCL1 | Capana10g000732 | 10 | 43,673,571–43,714,716 | 5736 | 1911 | 214.29 |
| CaDCL2 | Capana12g002509 | 12 | 220,158,256–220,168,547 | 4206 | 1401 | 158.07 |
| CaDCL3 | Capana08g000619 | 8 | 95,835,542–95,908,094 | 4956 | 1651 | 186.11 |
| CaDCL4 | Capana07g000265 | 7 | 11,520,463–11,570,768 | 4854 | 1617 | 182.23 |
| CaRDR1 | Capana11g001709 | 11 | 191,597,130–191,608,269 | 3351 | 1116 | 127.40 |
| CaRDR2 | Capana03g000988 | 3 | 16,378,521–16,387,609 | 3393 | 1130 | 128.28 |
| CaRDR3a | Capana07g000168 | 7 | 8,346,974–8,399,430 | 3024 | 1007 | 114.57 |
| CaRDR3b | Capana08g000377 | 8 | 51,083,159–51,128,885 | 5049 | 1682 | 191.94 |
| CaRDR5 | Capana09g000243 | 9 | 7,491,673–7,505,214 | 3204 | 1067 | 122.95 |
| CaRDR6 | Capana05g000179 | 5 | 2,848,056–2,858,044 | 3591 | 1196 | 135.98 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; Mo, N.; Muhammad, T.; Liang, Y. Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.). Int. J. Mol. Sci. 2018, 19, 1038. https://doi.org/10.3390/ijms19041038
Qin L, Mo N, Muhammad T, Liang Y. Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.). International Journal of Molecular Sciences. 2018; 19(4):1038. https://doi.org/10.3390/ijms19041038
Chicago/Turabian StyleQin, Lei, Ning Mo, Tayeb Muhammad, and Yan Liang. 2018. "Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.)" International Journal of Molecular Sciences 19, no. 4: 1038. https://doi.org/10.3390/ijms19041038
APA StyleQin, L., Mo, N., Muhammad, T., & Liang, Y. (2018). Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.). International Journal of Molecular Sciences, 19(4), 1038. https://doi.org/10.3390/ijms19041038