A Comparison of the Effects of FATTY ACID DESATURASE 7 and HYDROPEROXIDE LYASE on Plant–Aphid Interactions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
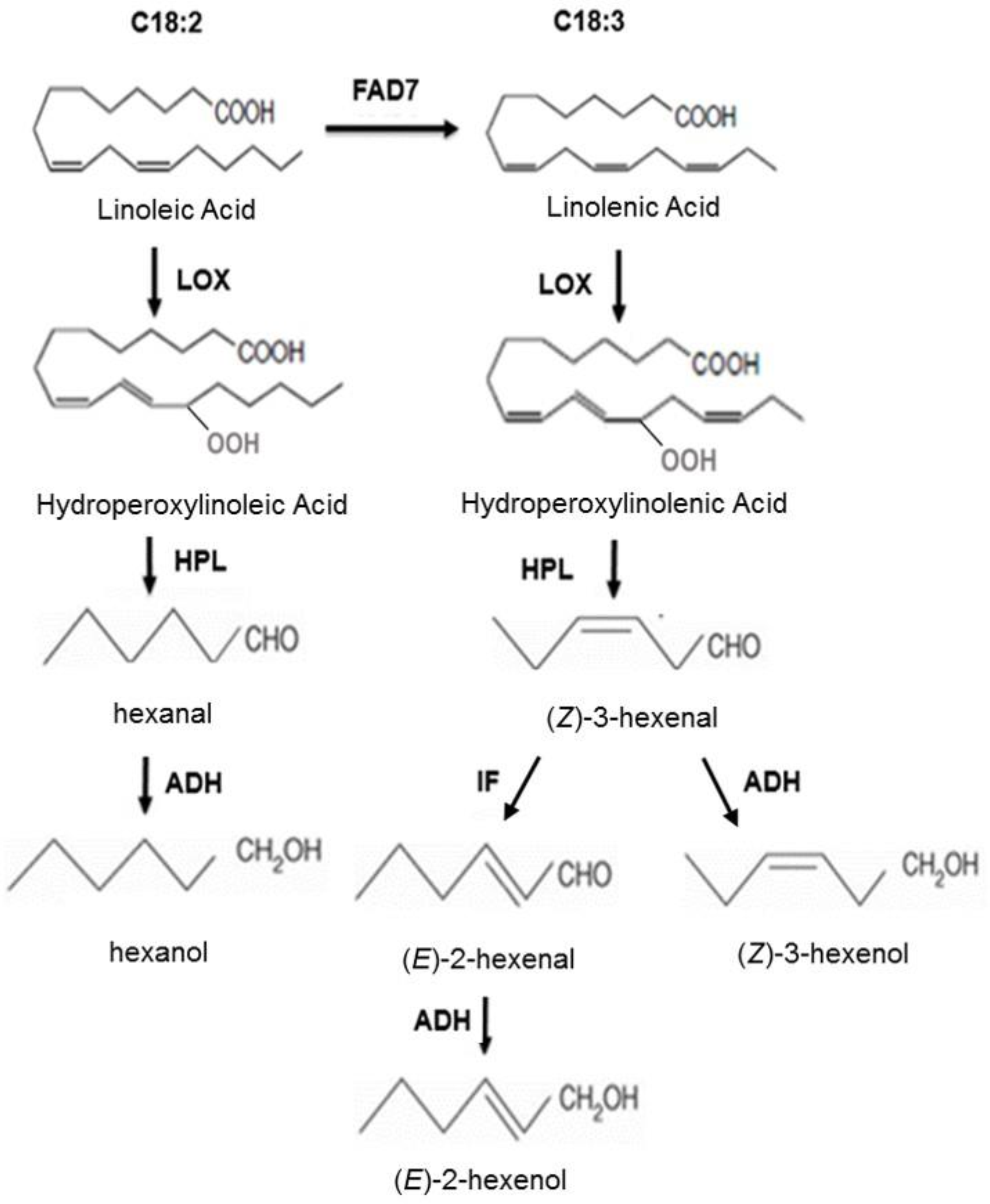
:1. Introduction
2. Results
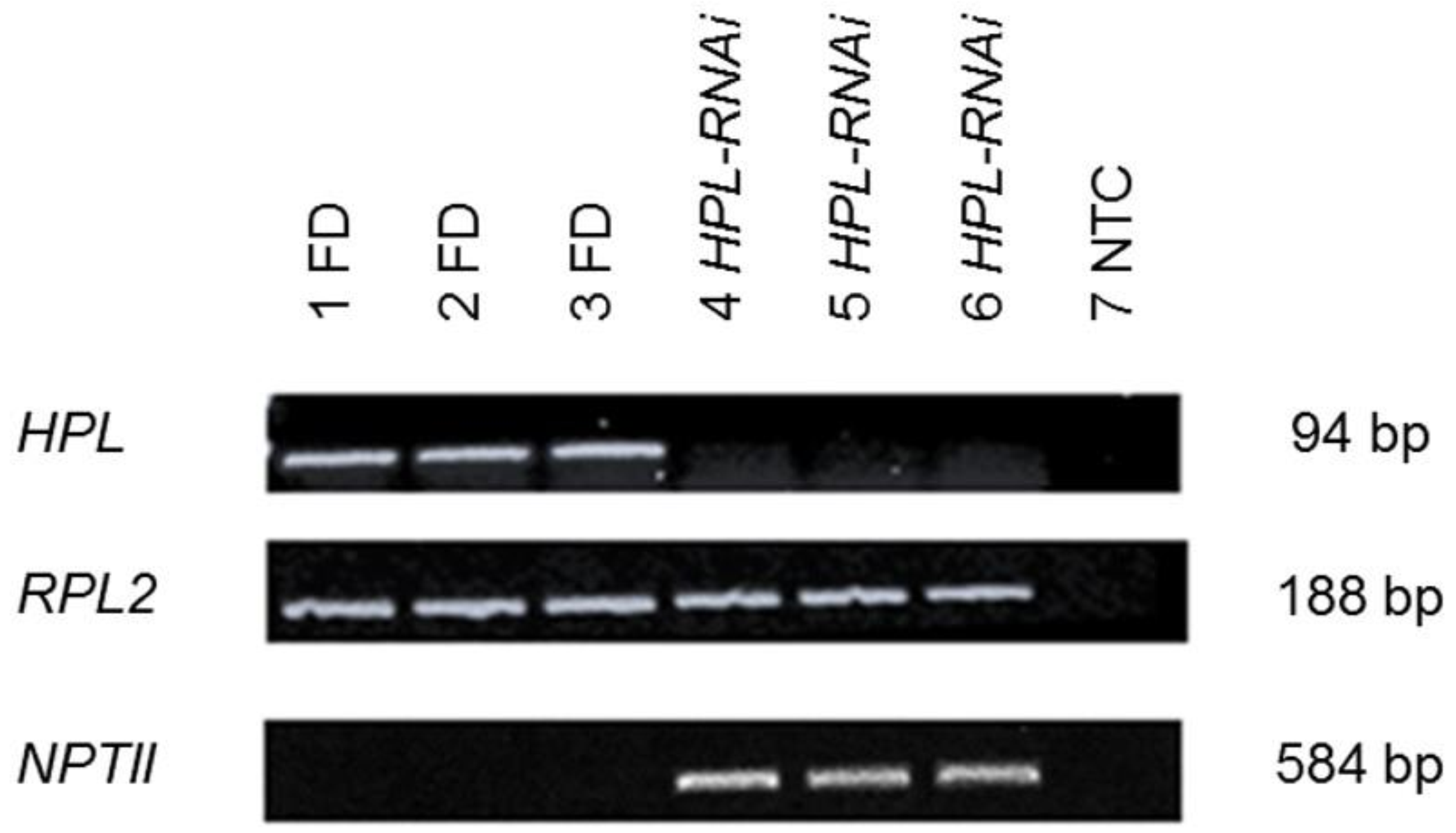
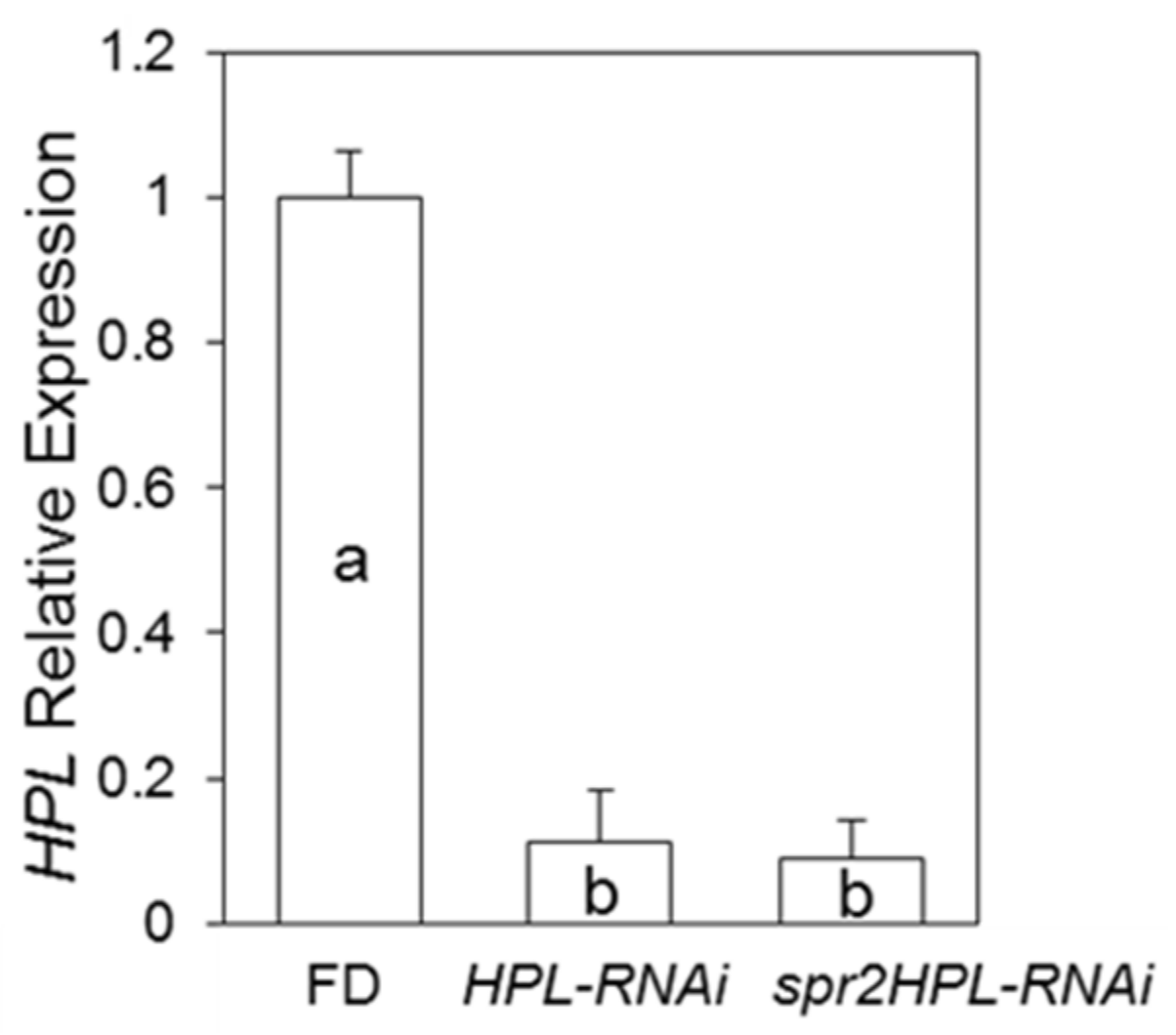
2.1. Confirmation of Silencing of HPL in Tomato
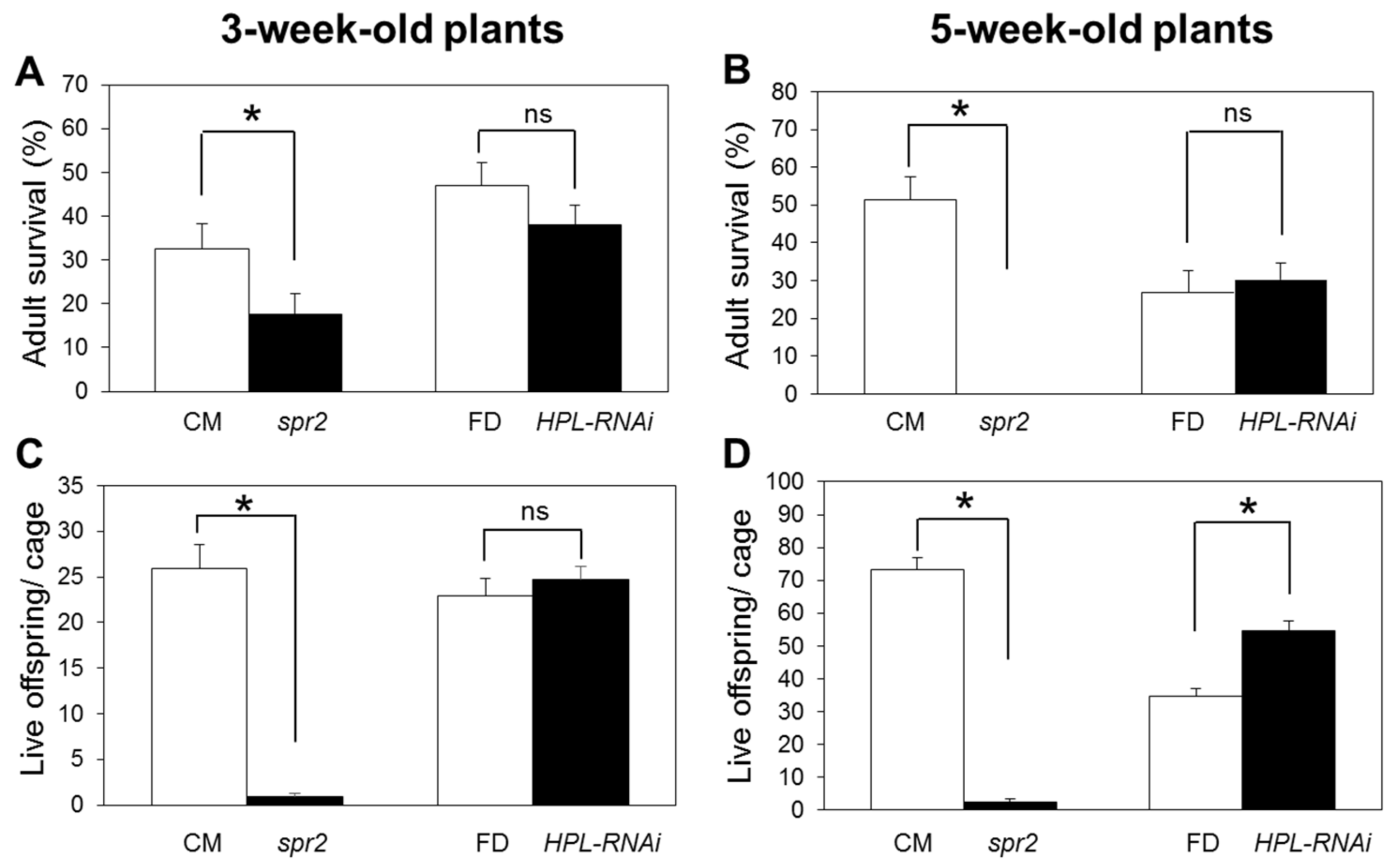
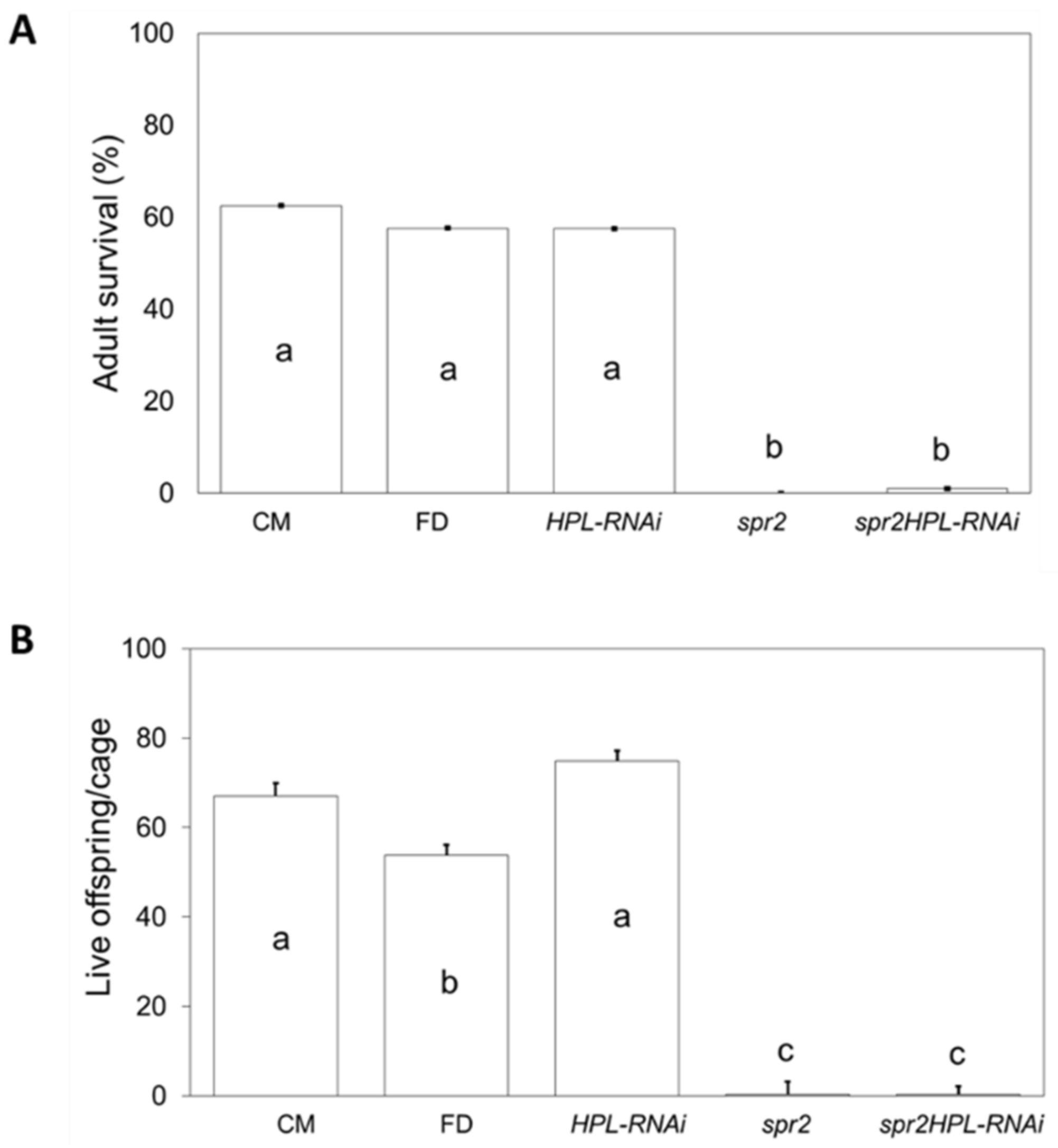
2.2. Influence of HPL and spr2 on Aphid Survival and Fecundity on Tomato
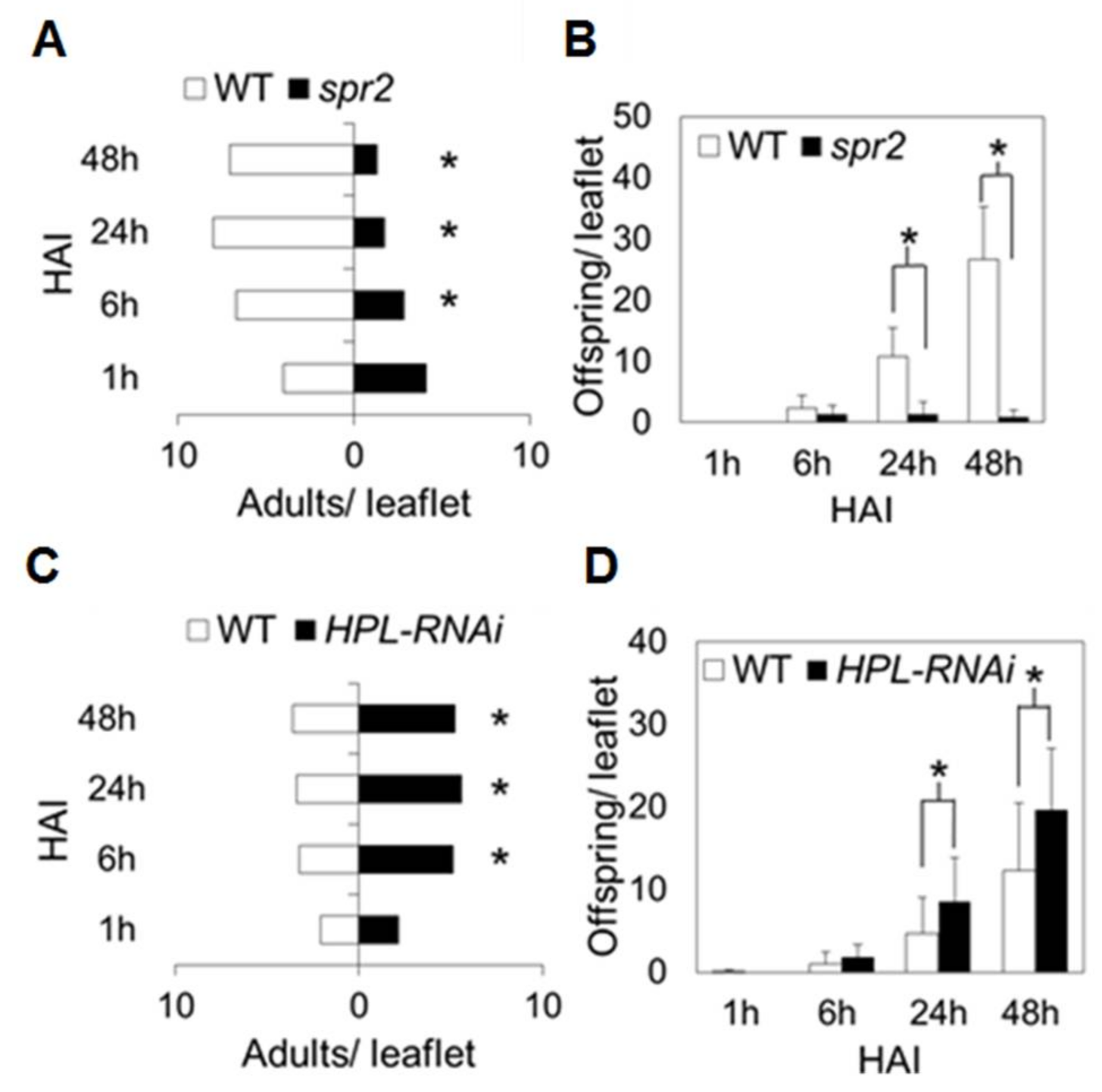
2.3. Influence of HPL and spr2 on Aphid Host Preference on Tomato
2.4. Silencing HPL in the spr2 Mutant
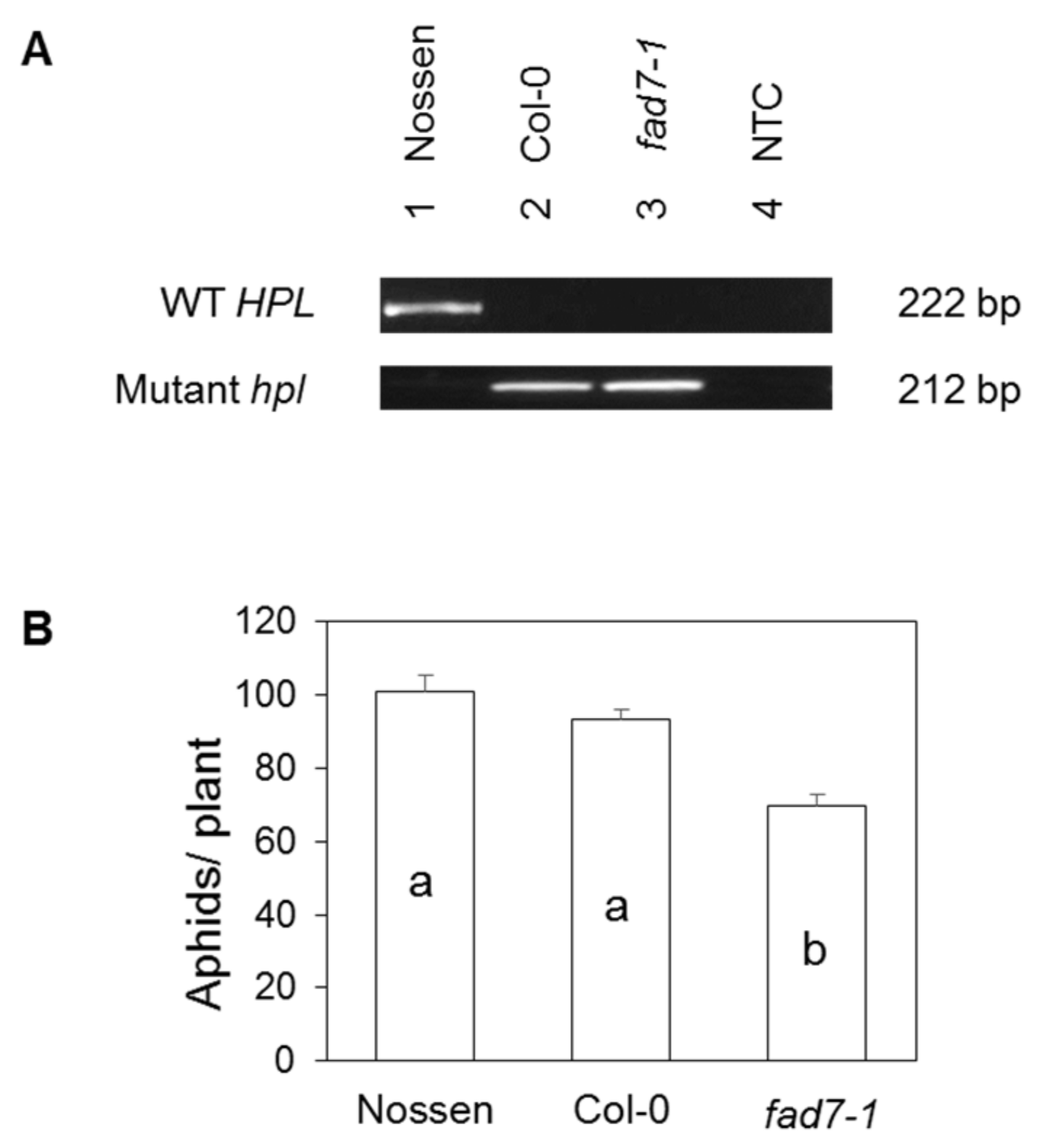
2.5. FAD7 and HPL in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Tomato Culture
4.2. Development of the spr2HPL-RNAi Line
4.3. Arabidopsis Culture and Materials Development
4.4. Identification of the HPL Mutation in the Arabidopsis fad7-1 Mutant
4.5. RNA Isolation and Gene Expression Analysis
4.6. Aphid Bioassays
4.6.1. Insect Materials
4.6.2. Aphid Survival and Fecundity on Tomato
4.6.3. Aphid Host Preference on Tomato
4.6.4. Aphid Survival and Fecundity on Arabidopsis
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Browse, J.; McCourt, P.; Somerville, C. A mutant of Arabidopsis deficient in C18:3 and C16:3 leaf lipids. Plant Physiol. 1986, 81, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, G.; Xu, C.; Lee, G.I.; Bauer, P.; Ling, H.Q.; Ganal, M.W.; Howe, G.A. The tomato suppressor of prosystemin-mediated responses2 gene encodes a fatty acid desaturase required for the biosynthesis of jasmonic acid and the production of a systemic wound signal for defense gene expression. Plant Cell 2003, 15, 1646–1661. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.; Arondel, V.; Iba, K.; Somerville, C. Cloning of a temperature-regulated gene encoding a chloroplast omega-3 desaturase from Arabidopsis thaliana. Plant Physiol. 1994, 106, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Berberich, T.; Harada, M.; Sugawara, K.; Kodama, H.; Iba, K.; Kusano, T. Two maize genes encoding omega-3 fatty acid desaturase and their differential expression to temperature. Plant Mol. Biol. 1998, 36, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Nishiuchi, T.; Seo, S.; Ohashi, Y.; Iba, K. Possible involvement of protein phosphorylation in the wound-responsive expression of Arabidopsis plastid ω-3 fatty acid desaturase gene. Plant Sci. 2000, 155, 153–160. [Google Scholar] [CrossRef]
- Dong, C.-J.; Cao, N.; Zhang, Z.-G.; Shang, Q.-M. Characterization of the fatty acid desaturase genes in cucumber: Structure, phylogeny, and expression patterns. PLoS ONE 2016, 11, e0149917. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Tsuyama, M.; Kobayashi, Y.; Kodama, H.; Iba, K. Trienoic fatty acids and plant tolerance of high temperature. Science 2000, 287, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Im, Y.J.; Kim, M.S.; Yang, K.Y.; Kim, Y.H.; Back, K.; Cho, B.H. Antisense expression of a ω-3 fatty acid desaturase gene in tobacco plants enhances susceptibility against pathogens. Can. J. Bot. 2004, 82, 297–303. [Google Scholar] [CrossRef]
- Yara, A.; Yaeno, T.; Hasegawa, M.; Seto, H.; Montillet, J.-L.; Kusumi, K.; Seo, S.; Iba, K. Disease resistance against Magnaporthe grisea is enhanced in transgenic rice with suppression of ω-3 fatty acid desaturases. Plant Cell Physiol. 2007, 48, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.A.; Arevalo-Soliz, L.M.; Jia, L.; Navarre, D.A.; Chen, Z.; Howe, G.A.; Meng, Q.W.; Smith, J.E.; Goggin, F.L. Loss of function of FATTY ACID DESATURASE 7 in tomato enhances basal aphid resistance in a salicylate-dependent manner. Plant Physiol. 2012, 158, 2028–2041. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Canoles, M.A.; Beaudry, R.M.; Li, C.; Howe, G. Deficiency of linolenic acid in Lefad7 mutant tomato changes the volatile profile and sensory perception of disrupted leaf and fruit tissue. J. Am. Soc. Hortic. Sci. 2006, 131, 284–289. [Google Scholar]
- Sanchez-Hernandez, C.; Lopez, M.G.; Delano-Frier, J.P. Reduced levels of volatile emissions in jasmonate-deficient spr2 tomato mutants favour oviposition by insect herbivores. Plant Cell Environ. 2006, 29, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, T.; Hernández, M.L.; Pennycooke, J.C.; Jiménez, P.; Martínez-Rivas, J.M.; Sanz, C.; Stockinger, E.J.; Sánchez-Serrano, J.J.; Sanmartín, M. Increasing ω-3 desaturase expression in tomato results in altered aroma profile and enhanced resistance to cold stress. Plant Physiol. 2010, 153, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, D.F.; Brown, G.C.; Jackson, D.M.; Hamilton-Kemp, T.R. Effects of some leaf-emitted volatile compounds on aphid population increase. J. Chem. Ecol. 1993, 19, 1875–1887. [Google Scholar] [CrossRef] [PubMed]
- Vancanneyt, G.; Sanz, C.; Farmaki, T.; Paneque, M.; Ortego, F.; Castanera, P.; Sanchez-Serrano, J.J. Hydroperoxide lyase depletion in transgenic potato plants leads to an increase in aphid performance. Proc. Natl. Acad. Sci. USA 2001, 98, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Qi, J.; Zhu, X.; Mao, B.; Zeng, L.; Wang, B.; Li, Q.; Zhou, G.; Xu, X.; Lou, Y.; et al. The rice hydroperoxide lyase OsHPL3 functions in defense responses by modulating the oxylipin pathway. Plant J. 2012, 71, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Chehab, E.W.; Kaspi, R.; Savchenko, T.; Rowe, H.; Negre-Zakharov, F.; Kliebenstein, D.; Dehesh, K. Distinct roles of jasmonates and aldehydes in plant-defense responses. PLoS ONE 2008, 3, e1904. [Google Scholar] [CrossRef] [PubMed]
- Halitschke, R.; Ziegler, J.; Keinanen, M.; Baldwin, I.T. Silencing of hydroperoxide lyase and allene oxide synthase reveals substrate and defense signaling crosstalk in Nicotiana attenuata. Plant J. 2004, 40, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.; Zhang, L.; Zhang, Z.; Chen, Z.; Sun, X. A tea hydroperoxide lyase gene, CsiHPL1, regulates tomato defense response against Prodenia litura (Fabricius) and Alternaria alternate f. sp. Lycopersici by modulating green leaf volatiles (GLVs) release and jasmonic acid (JA) gene expression. Plant Mol. Biol. Report. 2014, 32, 62–69. [Google Scholar] [CrossRef]
- Shen, J.; Tieman, D.; Jones, B.; Taylor, G.; Schmelz, E.; Huffaker, A.; Bies, D.; Chen, K.; Klee, H.J. A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 flavour volatiles in tomato. J. Exp. Bot. 2014, 65, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, F.; Tang, J.; Wang, W.; Zhang, F.; Wang, G.; Chu, J.; Yan, C.; Wang, T.; Chu, C.; et al. Activation of the jasmonic acid pathway by depletion of the hydroperoxide lyase OsHPL3 reveals crosstalk between the HPL and AOS branches of the oxylipin pathway in rice. PLoS ONE 2012. [Google Scholar] [CrossRef] [PubMed]
- Hanley, M.E.; Girling, R.D.; Felix, A.-E.; Olliff, E.D.; Newland, P.L.; Poppy, G.M. Olfactory selection of Plantago lanceolata declines with seedling age. Ann. Bot. 2013, 112, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Huang, M.-Y.; Palacio, K.; Schuler, M.A. Variations in CYP74B2 (hyperoxide lyase) gene expression differentially affect hexenal signaling in the Columbia and Landsberg erecta ecotypes of Arabidopsis. Plant Physiol. 2005, 139, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Madey, E.; Nowack, L.; Thompson, J. Isolation and characterization of lipid in phloem sap of canola. Planta 2002, 214, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.A.; Ryan, C.A. Suppressors of systemin signaling identify genes in the tomato wound response pathway. Genetics 1999, 153, 1411–1421. [Google Scholar] [PubMed]
- Vaughn, K.L.; Avila, C.A.; Padilla-Marcia, C.S.; Goggin, F.L. Development of fad7-1 single mutant Arabidopsis thaliana plants that are resistant to aphids. Discovery 2014, 15, 94–99. [Google Scholar]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef] [PubMed]
- Kasajima, I.; Ide, Y.; Ohkama-Ohtsu, N.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. A protocol for rapid DNA extraction from Arabidopsis thaliana for PCR analysis. Plant Mol. Biol. Report. 2004, 22, 49–52. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, evolutionary, and applied perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Gӧrlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Avila, C.A.; Tieman, D.M.; Klee, H.J.; Goggin, F.L. A Comparison of the Effects of FATTY ACID DESATURASE 7 and HYDROPEROXIDE LYASE on Plant–Aphid Interactions. Int. J. Mol. Sci. 2018, 19, 1077. https://doi.org/10.3390/ijms19041077
Li J, Avila CA, Tieman DM, Klee HJ, Goggin FL. A Comparison of the Effects of FATTY ACID DESATURASE 7 and HYDROPEROXIDE LYASE on Plant–Aphid Interactions. International Journal of Molecular Sciences. 2018; 19(4):1077. https://doi.org/10.3390/ijms19041077
Chicago/Turabian StyleLi, Jiamei, Carlos A. Avila, Denise M. Tieman, Harry J. Klee, and Fiona L. Goggin. 2018. "A Comparison of the Effects of FATTY ACID DESATURASE 7 and HYDROPEROXIDE LYASE on Plant–Aphid Interactions" International Journal of Molecular Sciences 19, no. 4: 1077. https://doi.org/10.3390/ijms19041077
APA StyleLi, J., Avila, C. A., Tieman, D. M., Klee, H. J., & Goggin, F. L. (2018). A Comparison of the Effects of FATTY ACID DESATURASE 7 and HYDROPEROXIDE LYASE on Plant–Aphid Interactions. International Journal of Molecular Sciences, 19(4), 1077. https://doi.org/10.3390/ijms19041077