Molecular Characterization of Gastric Epithelial Cells Using Flow Cytometry


Abstract
:1. Introduction
2. Results
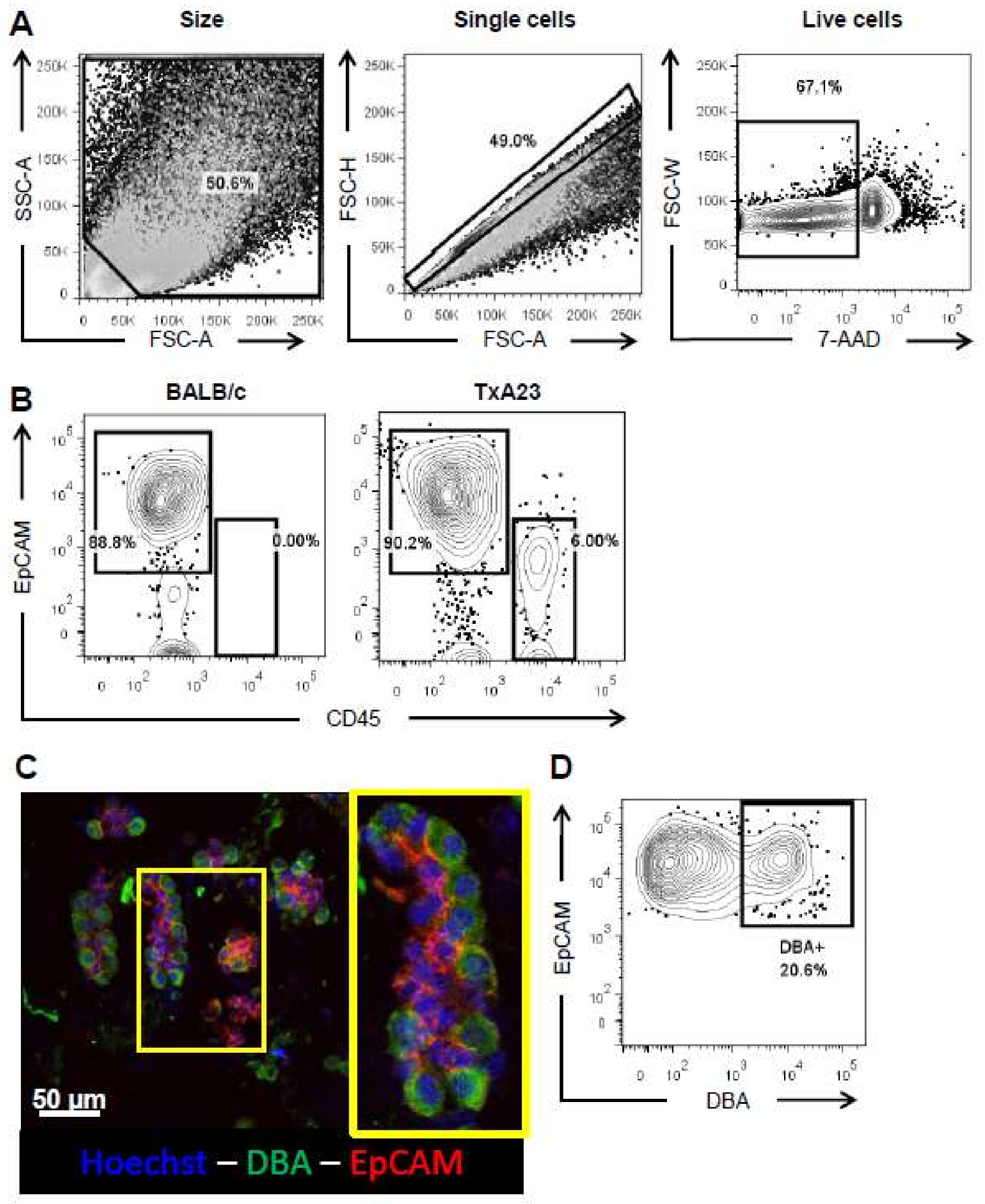
2.1. Enzymatic Digestion Results in a Single Cell Suspension Comprised of Major Gastric Epithelial Cell Lineages
2.2. Optimizing Flow Cytometric Analysis of Gastric Epithelial Cells
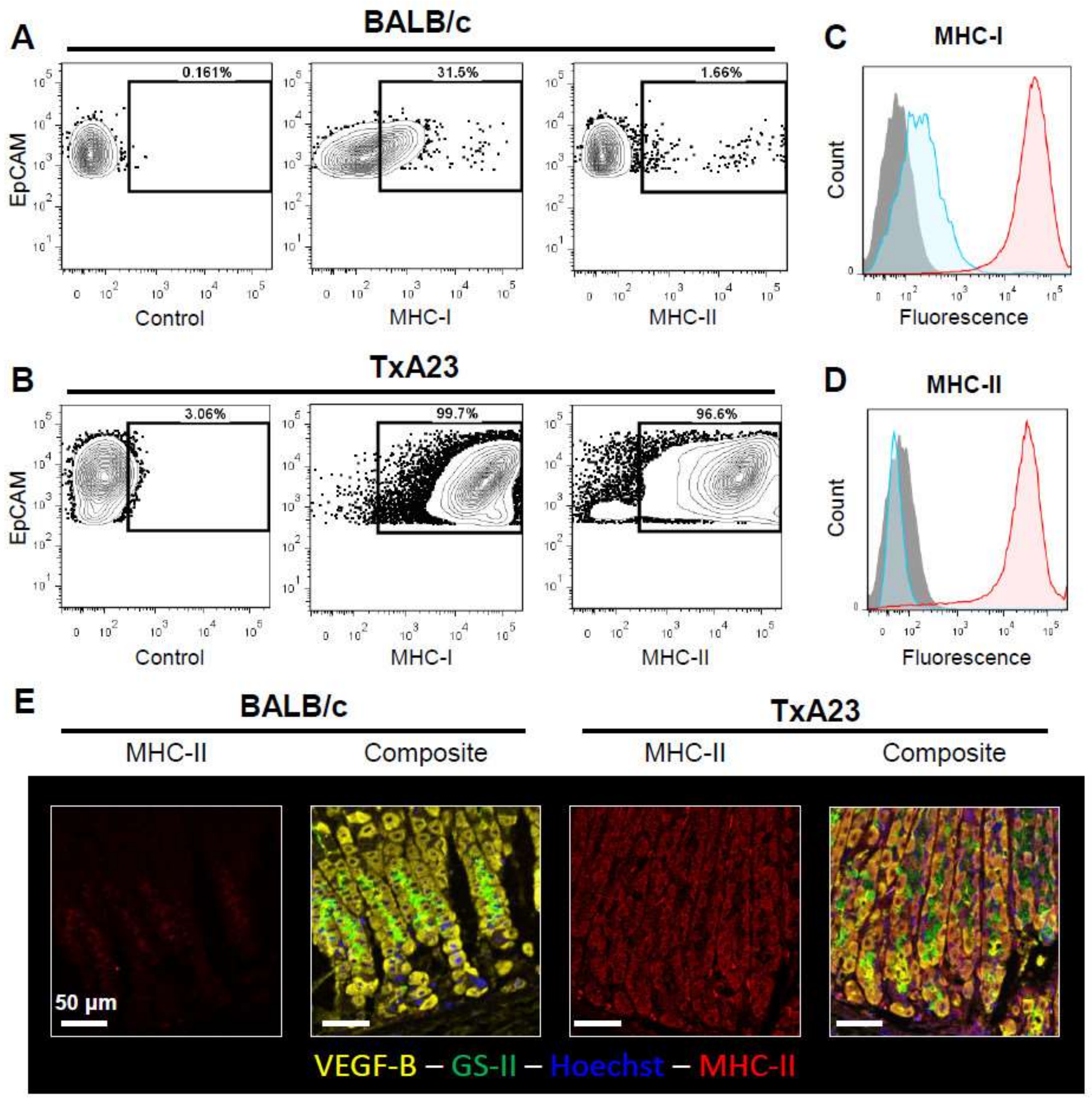
2.3. Gastric Epithelial Cells Upregulate MHC-I and MHC-II in Response to Chronic Inflammation
3. Discussion
4. Methods
4.1. Mice
4.2. Immunofluorescence/Immunohistochemistry
4.3. Generation of Gastric Epithelial Single Cell Suspensions
4.4. Quantitative Real Time PCR
4.5. Flow Cytometry
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ohata, H.; Kitauchi, S.; Yoshimura, N.; Mugitani, K.; Iwane, M.; Nakamura, H.; Yoshikawa, A.; Yanaoka, K.; Arii, K.; Tamai, H.; et al. Progression of chronic atrophic gastritis associated with helicobacter pylori infection increases risk of gastric cancer. Int. J. Cancer 2004, 109, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Landgren, A.M.; Landgren, O.; Gridley, G.; Dores, G.M.; Linet, M.S.; Morton, L.M. Autoimmune disease and subsequent risk of developing alimentary tract cancers among 4.5 million us male veterans. Cancer 2011, 117, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Correa, P. A human model of gastric carcinogenesis. Cancer Res. 1988, 48, 3554–3560. [Google Scholar] [PubMed]
- Zavros, Y.; van Antwerp, M.; Merchant, J.L. Use of flow cytometry to quantify mouse gastric epithelial cell populations. Dig. Dis. Sci. 2000, 45, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Wittwer, C. Flow cytometry: Principles and clinical applications in hematology. Clin. Chem. 2000, 46, 1221–1229. [Google Scholar] [PubMed]
- Lyons, A.B.; Parish, C.R. Determination of lymphocyte division by flow cytometry. J. Immunol. Methods 1994, 171, 131–137. [Google Scholar] [CrossRef]
- Jung, T.; Schauer, U.; Heusser, C.; Neumann, C.; Rieger, C. Detection of intracellular cytokines by flow cytometry. J. Immunol. Methods 1993, 159, 197–207. [Google Scholar] [CrossRef]
- Riccardi, C.; Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef] [PubMed]
- Krutzik, P.O.; Crane, J.M.; Clutter, M.R.; Nolan, G.P. High-content single-cell drug screening with phosphospecific flow cytometry. Nat. Chem. Biol. 2008, 4, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.D.; Jin, R.U.; Osaki, L.; Romero-Gallo, J.; Noto, J.; Peek, R.M., Jr.; Mills, J.C. Identification of alanyl aminopeptidase (CD13) as a surface marker for isolation of mature gastric zymogenic chief cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G955–G964. [Google Scholar] [CrossRef] [PubMed]
- Bertaux-Skeirik, N.; Feng, R.; Schumacher, M.A.; Li, J.; Mahe, M.M.; Engevik, A.C.; Javier, J.E.; Peek, R.M., Jr.; Ottemann, K.; Orian-Rousseau, V.; et al. CD44 plays a functional role in helicobacter pylori-induced epithelial cell proliferation. PLoS Pathog. 2015, 11, e1004663. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, K.L.; Bane, G.C.; Jazayeri, A.; Samuelson, L.C. Enhanced calcium signaling and acid secretion in parietal cells isolated from gastrin-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G145–G153. [Google Scholar] [CrossRef] [PubMed]
- McHugh, R.S.; Shevach, E.M.; Margulies, D.H.; Natarajan, K. A t cell receptor transgenic model of severe, spontaneous organ-specific autoimmunity. Eur. J. Immunol. 2001, 31, 2094–2103. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Khurana, S.S.; Bellone, C.J.; Capoccia, B.J.; Sagartz, J.E.; Kesman, R.A., Jr.; Mills, J.C.; DiPaolo, R.J. Autoimmune gastritis mediated by CD4+ T cells promotes the development of gastric cancer. Cancer Res. 2013, 73, 2117–2126. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Karam, S.M.; Gordon, J.I. Diphtheria toxin-mediated ablation of parietal cells in the stomach of transgenic mice. J. Biol. Chem. 1996, 271, 3671–3676. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.C.; Syder, A.J.; Hong, C.V.; Guruge, J.L.; Raaii, F.; Gordon, J.I. A molecular profile of the mouse gastric parietal cell with and without exposure to helicobacter pylori. Proc. Natl. Acad. Sci. USA 2001, 98, 13687–13692. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Crowe, S.E.; Behar, S.; Gunasena, H.; Ye, G.; Haeberle, H.; van Houten, N.; Gourley, W.K.; Ernst, P.B.; Reyes, V.E. The effect of class II major histocompatibility complex expression on adherence of helicobacter pylori and induction of apoptosis in gastric epithelial cells: A mechanism for T helper cell type 1-mediated damage. J. Exp. Med. 1998, 187, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Gunasena, H.; Cheng, Z.; Espejo, R.; Crowe, S.E.; Ernst, P.B.; Reyes, V.E. Helicobacter pylori urease binds to class ii mhc on gastric epithelial cells and induces their apoptosis. J. Immunol. 2000, 165, 1918–1924. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.N.; Al-Menhali, A.A.; Keeley, T.M.; Ren, J.; El-Zaatari, M.; Chen, X.; Merchant, J.L.; Ross, T.S.; Chew, C.S.; Samuelson, L.C. Hip1r is expressed in gastric parietal cells and is required for tubulovesicle formation and cell survival in mice. J. Clin. Investig. 2008, 118, 2459–2470. [Google Scholar] [CrossRef] [PubMed]
- El-Zaatari, M.; Kao, J.Y.; Tessier, A.; Bai, L.; Hayes, M.M.; Fontaine, C.; Eaton, K.A.; Merchant, J.L. Gli1 deletion prevents helicobacter-induced gastric metaplasia and expansion of myeloid cell subsets. PLoS ONE 2013, 8, e58935. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Dipaolo, R.J. A new mouse model of inflammation and gastric cancer. Oncoimmunology 2013, 2, e25911. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Makhlouf, N.T.; Anthony, B.A.; Teague, R.M.; DiPaolo, R.J. In vitro induced regulatory t cells are unique from endogenous regulatory t cells and effective at suppressing late stages of ongoing autoimmunity. PLoS ONE 2014, 9, e104698. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, V.G.; Doherty, J.M.; Chen, C.C.; Stappenbeck, T.S.; Konieczny, S.F.; Mills, J.C. The maturation of mucus-secreting gastric epithelial progenitors into digestive-enzyme secreting zymogenic cells requires mist1. Development 2007, 134, 211–222. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference | Disruption | Enzymes | Region | FACs Gating Description |
|---|---|---|---|---|
| Zavros et al.; 2000 [5] | Medimachine | Collagenase | Whole Stomach | No |
| Moore et al.; 2015 [11] | Medimachine | Dispase II | Corpus | No |
| Hinkle et al.; 2003 [13] | None | Pronase | Corpus | No |
| Bockerstett et al.; 2018 | None | Collagenase + Dispase II | Corpus | Yes |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bockerstett, K.A.; Wong, C.F.; Koehm, S.; Ford, E.L.; DiPaolo, R.J. Molecular Characterization of Gastric Epithelial Cells Using Flow Cytometry. Int. J. Mol. Sci. 2018, 19, 1096. https://doi.org/10.3390/ijms19041096
Bockerstett KA, Wong CF, Koehm S, Ford EL, DiPaolo RJ. Molecular Characterization of Gastric Epithelial Cells Using Flow Cytometry. International Journal of Molecular Sciences. 2018; 19(4):1096. https://doi.org/10.3390/ijms19041096
Chicago/Turabian StyleBockerstett, Kevin A., Chun Fung Wong, Sherri Koehm, Eric L. Ford, and Richard J. DiPaolo. 2018. "Molecular Characterization of Gastric Epithelial Cells Using Flow Cytometry" International Journal of Molecular Sciences 19, no. 4: 1096. https://doi.org/10.3390/ijms19041096
APA StyleBockerstett, K. A., Wong, C. F., Koehm, S., Ford, E. L., & DiPaolo, R. J. (2018). Molecular Characterization of Gastric Epithelial Cells Using Flow Cytometry. International Journal of Molecular Sciences, 19(4), 1096. https://doi.org/10.3390/ijms19041096