Transcriptional Profiling of Host Cell Responses to Virulent Haemophilus parasuis: New Insights into Pathogenesis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Transcriptome Sequencing and Annotation
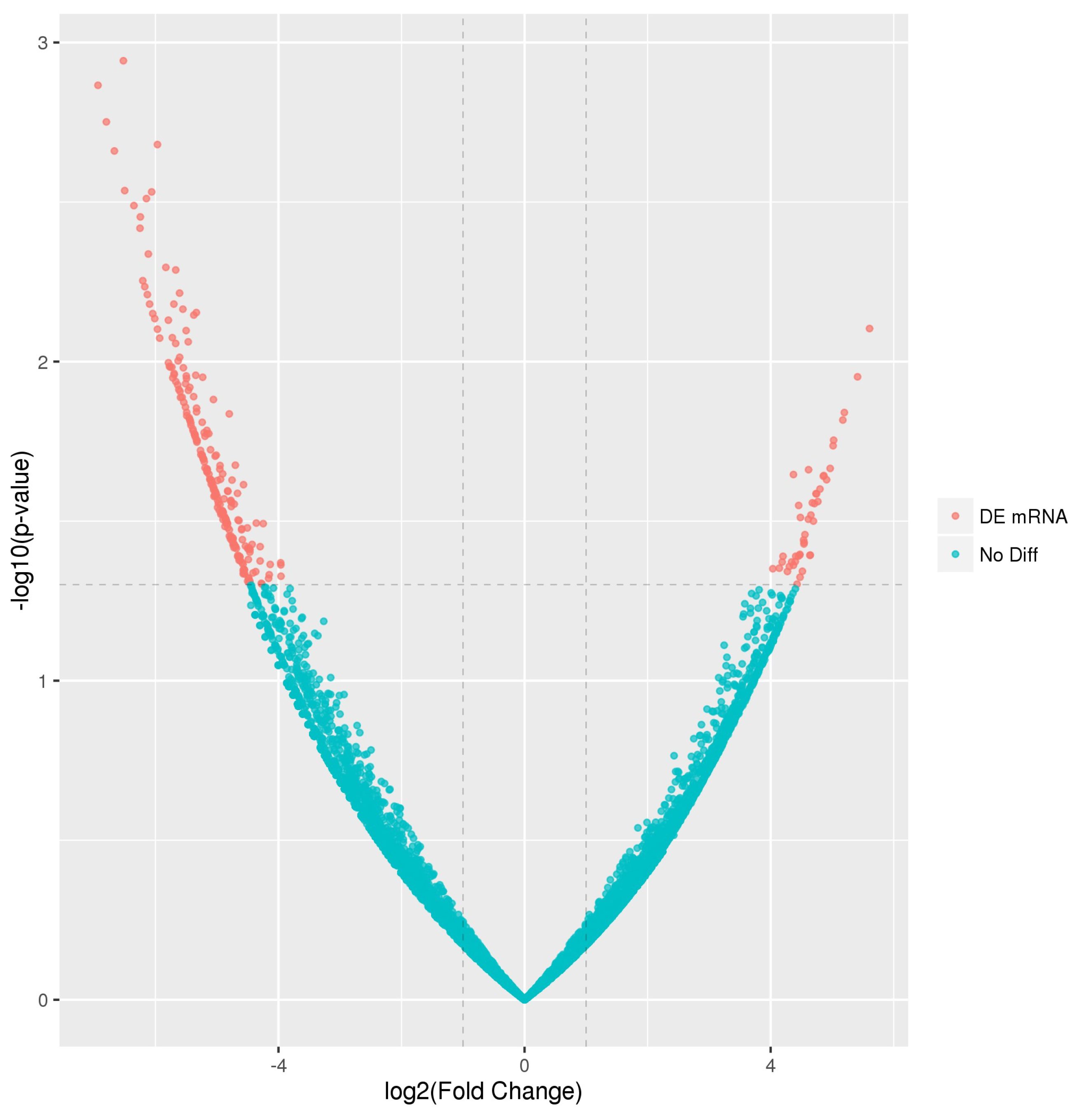
2.2. Identification of Differentially Expressed Genes (DEGs)
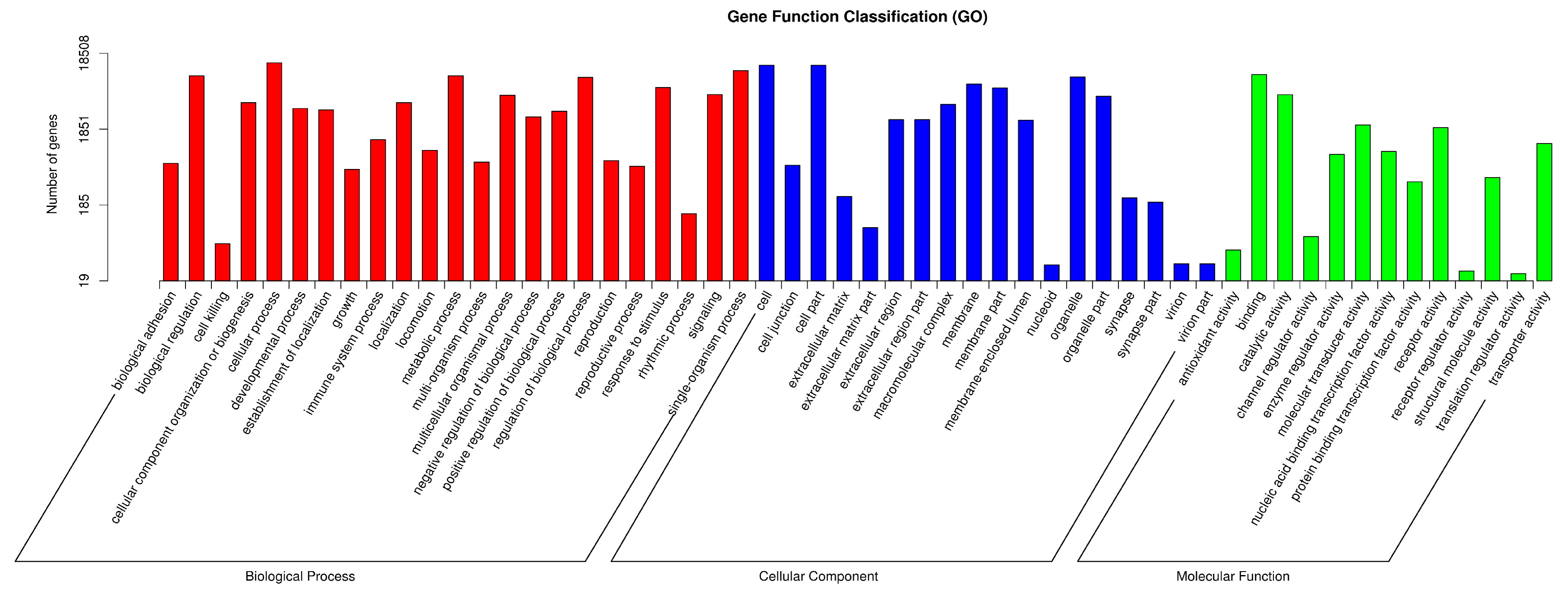
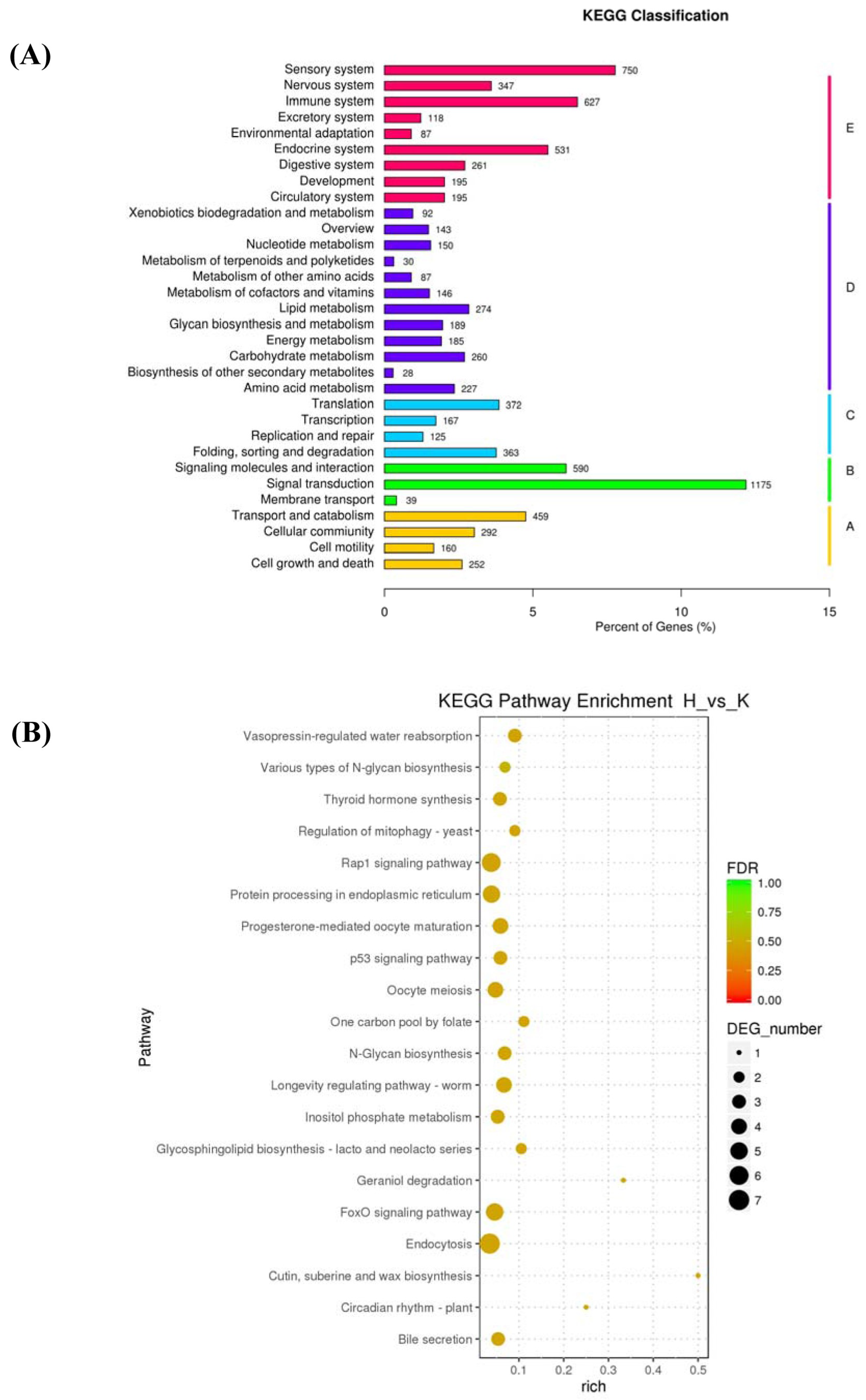
2.3. Functional Annotation of PAVEC Gene Expression Signature Triggered by H. parasuis Infection
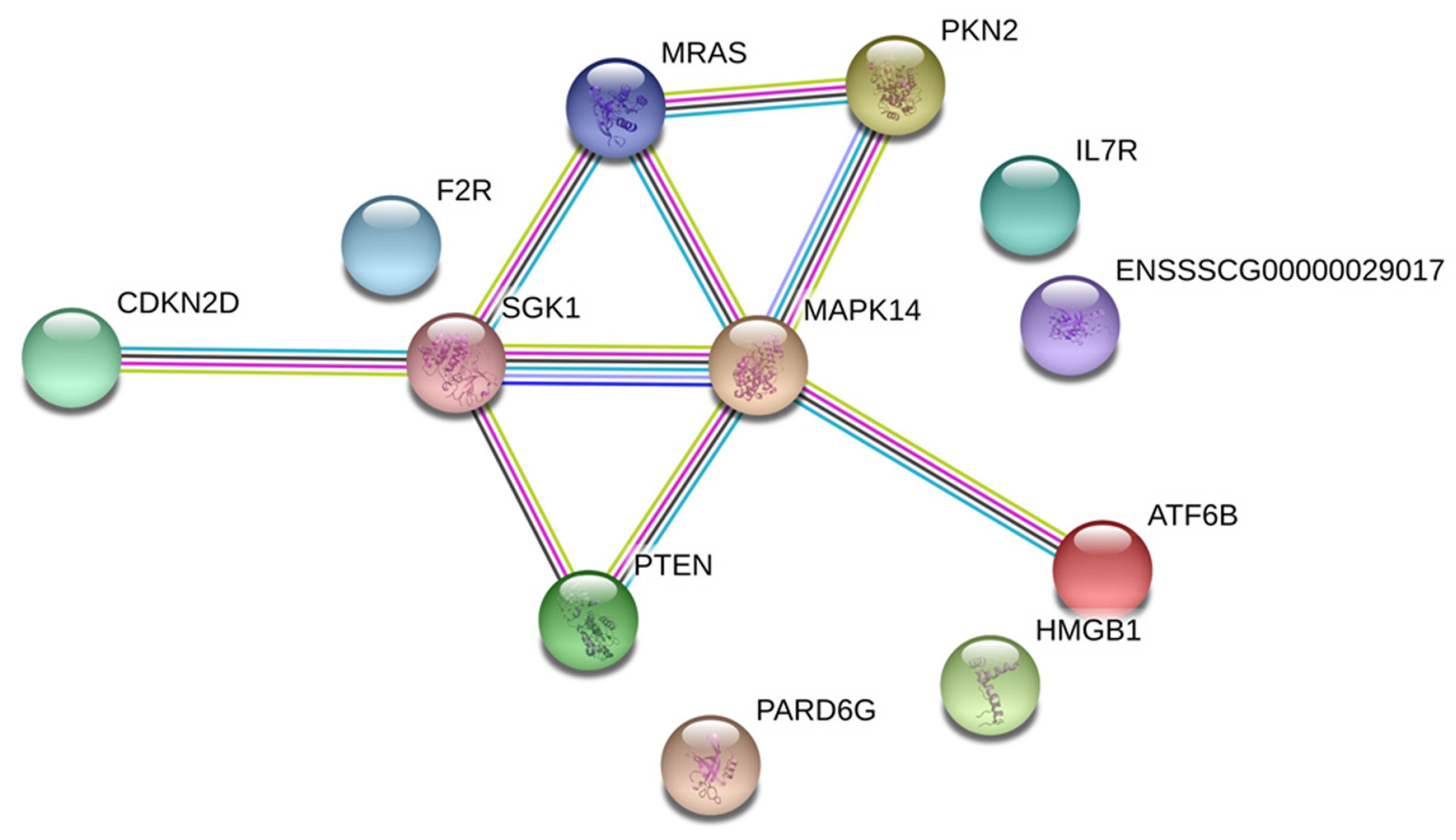
2.4. Analysis of the Association among DEGs of the Main Pathways Using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING)
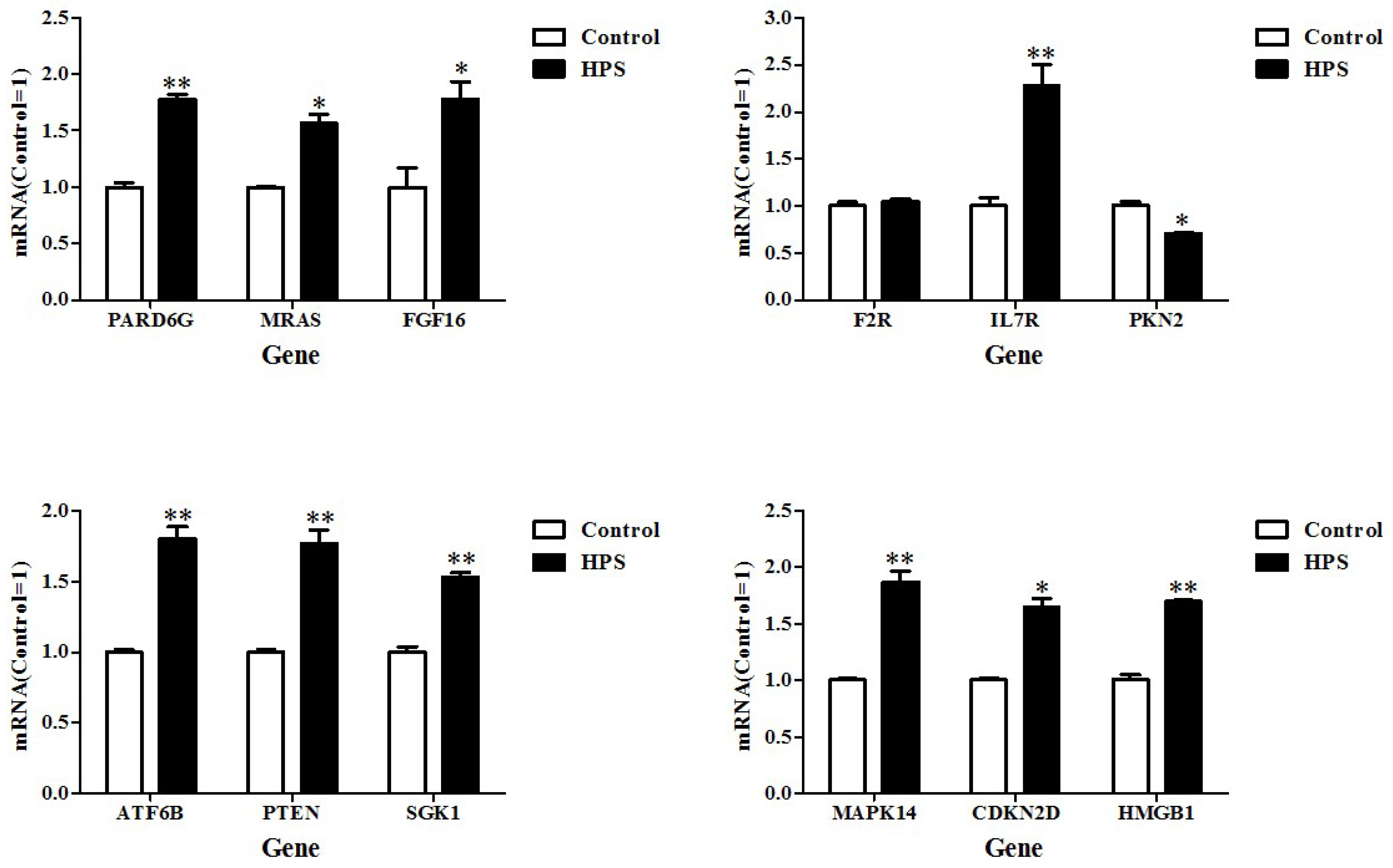
2.5. Real-Time Polymerase Chain Reaction (PCR) Verification of DEGs
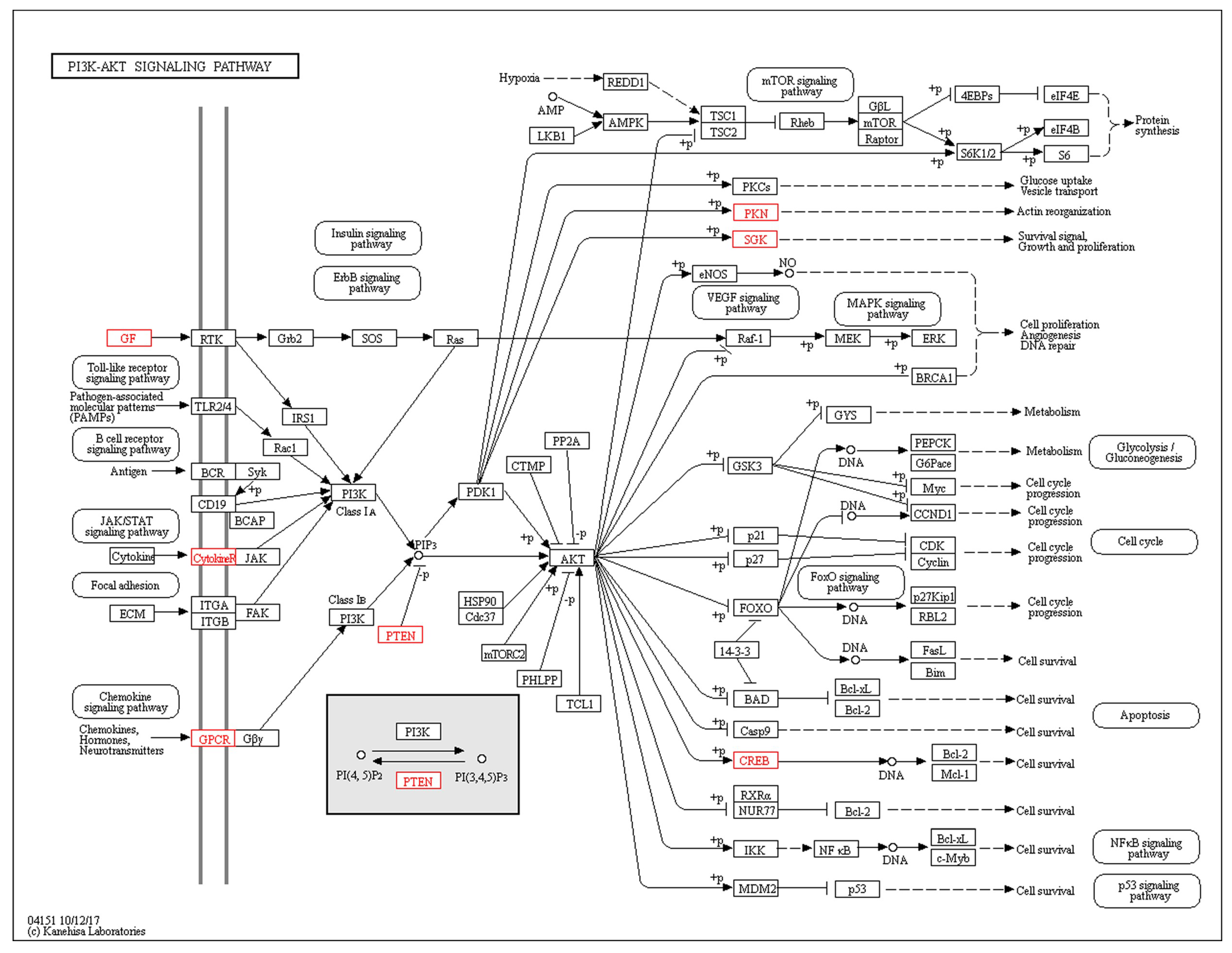
3. Discussion
4. Materials and Methods
4.1. Bacteria and Cells
4.2. Bacterial Infection
4.3. Construction of Library and Illumina Sequencing
4.4. Analysis of RNA-Seq Data
4.5. Validation by Real-Time qRT-PCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Li, G.; Niu, H.; Zhang, Y.; Li, Y.; Xie, F.; Langford, P.R.; Liu, S.; Wang, C. Haemophilus parasuis cytolethal distending toxin induces cell cycle arrest and p53-dependent apoptosis. PLoS ONE 2017, 12, e0177199. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.; Pijoan, C. Haemophilus parasuis: New trends on diagnosis, epidemiology and control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kielstein, P.; Rapp-Gabrielson, V.J. Designation of 15 serovars of Haemophilus parasuis on the basis of immunodiffusion using heat-stable antigen extracts. J. Clin. Microbiol. 1992, 30, 862–865. [Google Scholar] [PubMed]
- Guo, L.; Xu, L.; Wu, T.; Fu, S.; Qiu, Y.; Hu, C.A.; Ren, X.; Liu, R.; Ye, M. Evaluation of recombinant protein superoxide dismutase of Haemophilus parasuis strain SH0165 as vaccine candidate in a mouse model. Can. J. Microbiol. 2017, 63, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, M.; Blackall, P.J. Establishment, validation and use of the Kielstein-Rapp-Gabrielson serotyping scheme for Haemophilus parasuis. Aust. Vet. J. 2000, 78, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Correa-Fiz, F.; Galofre-Mila, N.; Costa-Hurtado, M.; Aragon, V. Identification of a surface epitope specific of virulent strains of Haemophilus parasuis. Vet. Microbiol. 2017, 198, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Feng, F.; Wang, H.; Xu, X.; Chen, H.; Cai, X.; Wang, X. Haemophilus parasuis CpxRA two-component system confers bacterial tolerance to environmental stresses and macrolide resistance. Microbiol. Res. 2018, 206, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gao, X.; Liu, C.; Lv, X.; Jiang, N.; Zheng, S. Deletion of the vacJ gene affects the biology and virulence in Haemophilus parasuis serovar 5. Gene 2017, 603, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wen, X.; He, L.; Yan, X.; Wen, Y.; Cao, S.; Huang, X.; Wu, R.; Wen, Y. The arcA gene contributes to the serum resistance and virulence of Haemophilus parasuis serovar 13 clinical strain EP3. Vet. Microbiol. 2016, 196, 67–71. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wen, X.; Yan, X.; Ding, L.; Cao, S.; Huang, X.; Wu, R.; Wen, Y. Effect of cheY deletion on growth and colonization in a Haemophilus parasuis serovar 13 clinical strain EP3. Gene 2016, 577, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Y.; Wen, Y.; Lau, G.W.; Huang, X.; Wu, R.; Yan, Q.; Huang, Y.; Zhao, Q.; Ma, X.; et al. HtrA Is Important for Stress Resistance and Virulence in Haemophilus parasuis. Infect. Immun. 2016, 84, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yu, Y.; Zeng, Z.; Ren, Y.; Yue, H. Deletion of the rfaE gene in Haemophilus parasuis SC096 strain attenuates serum resistance, adhesion and invasion. Microb. Pathog. 2014, 74, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, X.; Wu, Y.; Li, L.; Cao, R.; Cai, X.; Chen, H. Polysaccharide biosynthesis protein CapD is a novel pathogenicity-associated determinant of Haemophilus parasuis involved in serum-resistance ability. Vet. Microbiol. 2013, 164, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.J.; Hansen, M.; Troemel, E. Autophagy and innate immunity: Insights from invertebrate model organisms. Autophagy 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Blasdel, B.; Ceyssens, P.J.; Lavigne, R. Preparing cDNA libraries from lytic phage-infected cells for whole transcriptome analysis by RNA-Seq. Methods Mol. Biol. 2018, 1681, 185–194. [Google Scholar] [PubMed]
- Marino, R.; Capoferri, R.; Panelli, S.; Minozzi, G.; Strozzi, F.; Trevisi, E.; Snel, G.G.M.; Ajmone-Marsan, P.; Williams, J.L. Johne’s disease in cattle: An in vitro model to study early response to infection of Mycobacterium avium subsp. paratuberculosis using RNA-seq. Mol. Immunol. 2017, 91, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.W.; Humphrys, M.S.; Myers, G.S.A. A laboratory methodology for dual RNA-sequencing of bacteria and their host cells in vitro. Front. Microbiol. 2017, 8, 1830. [Google Scholar] [CrossRef] [PubMed]
- Pröll, M.J.; Neuhoff, C.; Schellander, K.; Uddin, M.J.; Cinar, M.U.; Sahadevan, S.; Qu, X.; Islam, M.A.; Poirier, M.; Müller, M.A.; et al. Transcriptome profile of lung dendritic cells after in vitro porcine reproductive and respiratory syndrome virus (PRRSV) infection. PLoS ONE 2017, 12, e0187735. [Google Scholar] [CrossRef] [PubMed]
- Wee, J.; Hong, S.Y.; Roze, L.V.; Day, D.M.; Chanda, A.; Linz, J.E. The Fungal bZIP Transcription Factor AtfB Controls Virulence-Associated Processes in Aspergillus parasiticus. Toxins 2017, 9, E287. [Google Scholar] [CrossRef] [PubMed]
- Brady, L.K.; Wang, H.; Radens, C.M.; Bi, Y.; Radovich, M.; Maity, A.; Ivan, C.; Ivan, M.; Barash, Y.; Koumenis, C. Transcriptome analysis of hypoxic cancer cells uncovers intron retention in EIF2B5 as a mechanism to inhibit translation. PLoS Biol. 2017, 15, e2002623. [Google Scholar] [CrossRef] [PubMed]
- Nueda, M.J.; Martorell-Marugan, J.; Martí, C.; Tarazona, S.; Conesa, A. Identification and visualisation of differential isoform expression in RNA-seq time series. Bioinformatics 2018, 34, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Badr, E.; ElHefnawi, M.; Heath, L.S. Computational identification of tissue-specific splicing regulatory elements in human genes from RNA-Seq data. PLoS ONE 2016, 11, e0166978. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Teh, C.S.J.; Thong, K.L. Biofilm-related diseases and omics: Global transcriptional profiling of Enterococcus faecium reveals different gene expression patterns in the biofilm and planktonic cells. OMICS 2017, 21, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Martins, A.J.; Shallom, S.; Kamenyeva, O.; Kashyap, A.; Sampaio, E.P.; Kabat, J.; Olivier, K.N.; Zelazny, A.M.; Tsang, J.S.; et al. Transcriptional response of respiratory epithelium to nontuberculous mycobacteria. Am. J. Respir. Cell. Mol. Biol. 2018, 58, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Matson, D.R.; Hardin, H.; Buehler, D.; Lloyd, R.V. AKT activity is elevated in aggressive thyroid neoplasms where it promotes proliferation and invasion. Exp. Mol. Pathol. 2017, 103, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Li, X.; Ye, P.; Wang, G.; Dai, W.; Liu, Y.; Gao, Q.; Shen, G. Oxymatrine induces apoptosis and inhibits invasion in Gallbladder carcinoma via PTEN/PI3K/AKT pathway. Cytotechnology 2018, 70, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yuan, B.; Yuan, W.; Wang, C.; Gao, R.; Wang, J. The expression and clinical significance of high mobility group nucleosome binding domain 5 in human osteosarcoma. Tumour. Biol. 2014, 35, 6539–6547. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; He, L.; Yao, K.; Tan, J.; Zeng, Q.; Dai, Y.; Liu, J.; Tang, Y. Knockdown of HMGN5 increases the chemosensitivity of human urothelial bladder cancer cells to cisplatin by targeting PI3K/Akt signaling. Oncol. Lett. 2017, 14, 6463–6470. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.J.; Chen, X.; Wang, Y.W.; Liu, H.T.; Ma, R.R.; Gao, P. miR-1268b confers chemosensitivity in breast cancer by targeting ERBB2-mediated PI3K-AKT pathway. Oncotarget 2017, 8, 89631–89642. [Google Scholar] [CrossRef] [PubMed]
- Temme, S.; Friebe, D.; Schmidt, T.; Poschmann, G.; Hesse, J.; Steckel, B.; Stühler, K.; Kunz, M.; Dandekar, T.; Ding, Z.; et al. Genetic profiling and surface proteome analysis of human atrial stromal cells and rat ventricular epicardium-derived cells reveals novel insights into their cardiogenic potential. Stem Cell Res. 2017, 25, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.D.; Liu, W.; Fang, W.G.; Kim, K.S.; Chen, Y.H. Vascular endothelial growth factor receptor 1 contributes to Escherichia coli K1 invasion of human brain microvascular endothelial cells through the phosphatidylinositol 3-kinase/Akt signaling pathway. Infect. Immun. 2010, 78, 4809–4816. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Hattori, S.; Kasai, K. Lipopolysaccharide activates Akt in vascular smooth muscle cells resulting in induction of inducible nitric oxide synthase through nuclear factor-κB activation. Eur. J. Pharmacol. 2003, 481, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Tang, B.; Jia, Y.P.; Zhu, P.; Zhuang, Y.; Fang, Y.; Li, Q.; Wang, K.; Zhang, W.J.; Guo, G.; et al. Helicobacter pylori CagA protein negatively regulates autophagy and promotes inflammatory response via c-Met-PI3K/Akt-mTOR signaling pathway. Front. Cell. Infect. Microbiol. 2017, 7, 417. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Sun, F.; Wang, F.; Zhang, J.; Zheng, B.; Zhong, J.; Yue, T.; Zheng, X.; Xu, J.F.; Wang, C.Y. Aloperine protects mice against DSS-induced colitis by PP2A-mediated PI3K/Akt/mTOR signaling suppression. Mediat. Inflamm. 2017, 2017, 5706152. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Qiu, M.; Zhou, H.; Wang, T.; Guo, W. PTEN, insulin resistance and cancer. Curr. Pharm. Des. 2017, 23, 3667–3676. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Guo, X.; Zhang, J.; Wu, D.; Hu, X.; Li, J.; Lan, Q.; Liu, Y.; Dong, W. PTEN gene induces cell invasion and migration via regulating AKT/GSK-3β/β-catenin signaling pathway in human gastric cancer. Dig. Dis. Sci. 2017, 62, 3415–3425. [Google Scholar] [CrossRef] [PubMed]
- Hilbi, H.; Kortholt, A. Role of the small GTPase Rap1 in signal transduction, cell dynamics and bacterial infection. Small GTPases 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, X.; Hardwidge, P.R. Heat-labile enterotoxin-induced activation of NF-κB and MAPK pathways in intestinal epithelial cells impacts enterotoxigenic Escherichia coli (ETEC) adherence. Cell. Microbiol. 2012, 14, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Le, N.T.; Tran, T.D.; Kim, E.H.; Park, S.S.; Luong, T.T.; Chung, K.T.; Pyo, S.; Rhee, D.K. Streptococcus pneumoniae ClpL modulates adherence to A549 human lung cells through Rap1/Rac1 activation. Infect. Immun. 2014, 82, 3802–3810. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Sukhova, G.K.; Wong, H.K.; Xu, A.; Tergaonkar, V.; Vanhoutte, P.M.; Tang, E.H. Rap1 induces cytokine production in pro-inflammatory macrophages through NF-κB signaling and is highly expressed in human atherosclerotic lesions. Cell Cycle 2015, 14, 3580–3592. [Google Scholar] [CrossRef] [PubMed]
- Marques, E.; Klefström, J. Par6 family proteins in cancer. Oncoscience 2015, 2, 894–895. [Google Scholar] [PubMed]
- Erdogan, E.; Lamark, T.; Stallings-Mann, M.; Lee, Jamieson.; Pellecchia, M.; Thompson, E.A.; Johansen, T.; Fields, A.P. Aurothiomalate inhibits transformed growth by targeting the PB1 domain of protein kinase Ciota. J. Biol. Chem. 2006, 281, 28450–28459. [Google Scholar] [CrossRef] [PubMed]
- Regala, R.P.; Thompson, E.A.; Fields, A.P. Atypical protein kinase C iota expression and aurothiomalate sensitivity in human lung cancer cells. Cancer. Res. 2008, 68, 5888–5895. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Shen, Y.; Lu, Z.; Shang, D.; Zhao, Z.; Lu, Y.; Wu, Y.; Zhang, Y.; Tu, Z.; Liu, H. Roles of partitioning-defective protein 6 (Par6) and its complexes in the proliferation, migration and invasion of cancer cells. Clin. Exp. Pharmacol. Physiol. 2017, 44, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Bullock, M. FOXO factors and breast cancer: Outfoxing endocrine resistance. Endocr. Relat. Cancer 2016, 23, R113–R130. [Google Scholar] [CrossRef] [PubMed]
- Eijkelenboom, A.; Burgering, B.M. FOXOs: Signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Hornsveld, M.; Dansen, T.B.; Derksen, P.W.; Burgering, B.M.T. Re-evaluating the role of FOXOs in cancer. Semin. Cancer Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cheng, J.; North, B.J.; Wei, W. Functional analyses of major cancer-related signaling pathways in Alzheimer’s disease etiology. Biochim. Biophys. Acta 2017, 1868, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Wang, H.; Gaur, U.; Little, P.J.; Xu, J.; Zheng, W. FOXO signaling pathways as therapeutic targets in cancer. Int. J. Biol. Sci. 2017, 13, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Yang, F.; Yang, J.; Bei, W.; Cai, X.; Chen, L.; Dong, J.; Zhou, R.; Jin, M.; Jin, Q.; et al. Complete genome sequence of Haemophilus parasuis SH0165. J. Bacteriol. 2009, 191, 1359–1360. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Zhang, M.; Xu, J.; Ou, J.; Wang, Y.; Liu, H.; Liu, J.; Chen, H.; Bei, W. Immunogenicity and protective efficacy of recombinant Haemophilus parasuis SH0165 putative outer membrane proteins. Vaccine 2013, 31, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, A.; Chamorro, S.; Rodríguez-Gago, M.; Alvarez, B.; Molina, M.J.; Rodríguez-Barbosa, J.I.; Sánchez, A.; Ramírez, P.; Muñoz, A.; Domínguez, J.; et al. Isolation and characterization of immortalized porcine aortic endothelial cell lines. Vet. Immunol. Immunopathol. 2002, 89, 91–98. [Google Scholar] [CrossRef]
- Fu, S.; Liu, H.; Xu, L.; Qiu, Y.; Liu, Y.; Wu, Z.; Ye, C.; Hou, Y.; Hu, C.A. Baicalin modulates NF-κB and NLRP3 inflammasome signaling in porcine aortic vascular endothelial cells Infected by Haemophilus parasuis Causing Glässer’s disease. Sci. Rep. 2018, 8, 807. [Google Scholar] [CrossRef] [PubMed]
- Beaudet, J.; Tulman, E.R.; Pflaum, K.; Liao, X.; Kutish, G.F.; Szczepanek, S.M.; Silbart, L.K.; Geary, S.J. Transcriptional profiling of the chicken tracheal response to virulent Mycoplasma gallisepticum strain Rlow. Infect. Immun. 2017, 85, e00343-17. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, H.; Li, X.; Li, Q.; Ma, Z.; Bai, J.; Qiao, Z.; Feng, R. Transcriptional profiling of host cell responses to encephalomyocarditis virus (EMCV). Virol. J. 2017, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Li, J.; Liang, X.; Yang, Z.; Jiang, Z. Transcriptome profiling of influenza A virus-infected lung epithelial (A549) cells with lariciresinol-4-β-d-glucopyranoside treatment. PLoS ONE 2017, 12, e0173058. [Google Scholar] [CrossRef] [PubMed]
- Namani, A.; Matiur Rahaman, M.; Chen, M.; Tang, X. Gene-expression signature regulated by the KEAP1-NRF2-CUL3 axis is associated with a poor prognosis in head and neck squamous cell cancer. BMC Cancer 2018, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Fu, S.; Yang, B.; Liu, Q.; Ahmed, S.; Xu, L.; Xiong, J.; Cao, J.; Qiu, Y. Comparative transcriptional profiling of tildipirosin-resistant and sensitive Haemophilus parasuis. Sci. Rep. 2017, 7, 7517. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Busscher, H.J.; Chakladar, J.; van der Mei, H.C.; Chaffin, W.L. Transcriptional profiling of C. albicans in a two species biofilm with Rothia dentocariosa. Front. Cell. Infect. Microbiol. 2017, 7, 311. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Seleem, M.N.; Hussain, H.I.; Huang, L.; Hao, H.; Yuan, Z. Comparative virulence studies and transcriptome analysis of Staphylococcus aureus strains isolated from animals. Sci. Rep. 2016, 6, 35442. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Guo, J.; Li, R.; Qiu, Y.; Ye, C.; Liu, Y.; Wu, Z.; Guo, L.; Hou, Y.; Hu, C.-A.A. Transcriptional Profiling of Host Cell Responses to Virulent Haemophilus parasuis: New Insights into Pathogenesis. Int. J. Mol. Sci. 2018, 19, 1320. https://doi.org/10.3390/ijms19051320
Fu S, Guo J, Li R, Qiu Y, Ye C, Liu Y, Wu Z, Guo L, Hou Y, Hu C-AA. Transcriptional Profiling of Host Cell Responses to Virulent Haemophilus parasuis: New Insights into Pathogenesis. International Journal of Molecular Sciences. 2018; 19(5):1320. https://doi.org/10.3390/ijms19051320
Chicago/Turabian StyleFu, Shulin, Jing Guo, Ruizhi Li, Yinsheng Qiu, Chun Ye, Yu Liu, Zhongyuan Wu, Ling Guo, Yongqing Hou, and Chien-An Andy Hu. 2018. "Transcriptional Profiling of Host Cell Responses to Virulent Haemophilus parasuis: New Insights into Pathogenesis" International Journal of Molecular Sciences 19, no. 5: 1320. https://doi.org/10.3390/ijms19051320
APA StyleFu, S., Guo, J., Li, R., Qiu, Y., Ye, C., Liu, Y., Wu, Z., Guo, L., Hou, Y., & Hu, C. -A. A. (2018). Transcriptional Profiling of Host Cell Responses to Virulent Haemophilus parasuis: New Insights into Pathogenesis. International Journal of Molecular Sciences, 19(5), 1320. https://doi.org/10.3390/ijms19051320