Fluorescent Proteins for Investigating Biological Events in Acidic Environments

Abstract
:
1. Introduction
2. GFP-Like Protein
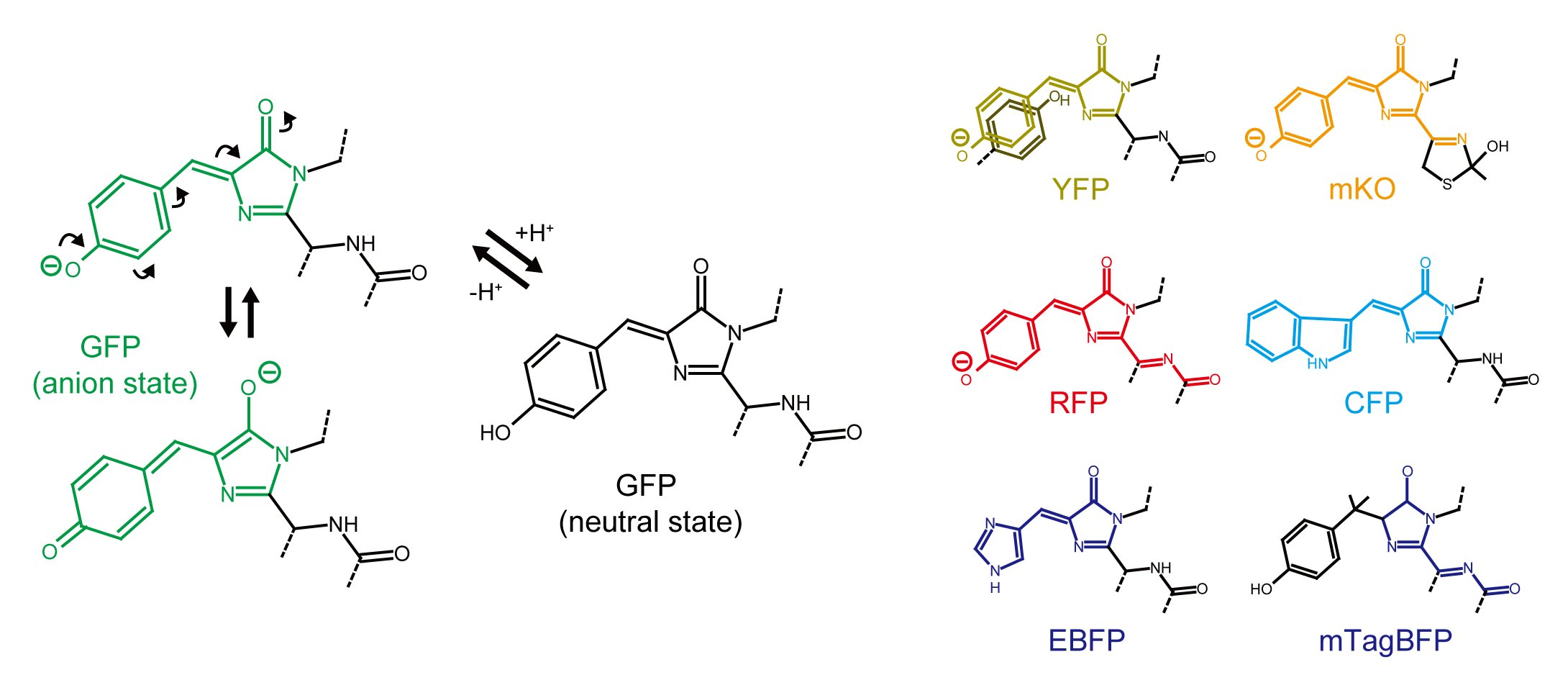
2.1. Green-Yellow-Red Fluorescent Protein
2.2. Blue-Cyan Fluorescent Protein
2.3. Character and Stability of β-Barrel Structure
2.4. Resistance to Degradative Enzymes in Lysosomes
3. FP-Based Sensor for Acidic Organelle Imaging
3.1. FP-Based Sensor and Chemical Dye-Based Sensor
3.2. pH Sensors
3.3. Calcium Ion Sensors
3.4. Zinc Ion Sensors
3.5. Chloride Ion Sensors
4. Conclusions and Future Prospective
Acknowledgments
Conflicts of Interest
References
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed]
- Settembre, C.; Fraldi, A.; Medina, D.L.; Ballabio, A. Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nat. Rev. Mol. Cell Biol. 2013, 14, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Turk, B.; Turk, V. Lysosomes as “suicide bags” in cell death: Myth or reality? J. Biol. Chem. 2009, 284, 21783–21787. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Docampo, R. Acidic calcium stores open for business: Expanding the potential for intracellular Ca2+ signaling. Trends Cell Biol. 2010, 20, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Stauber, T.; Jentsch, T.J. Chloride in Vesicular Trafficking and Function. Annu. Rev. Physiol. 2013, 75, 453–477. [Google Scholar] [CrossRef] [PubMed]
- Zoncu, R.; Bar-Peled, L.; Efeyan, A.; Wang, S.; Sancak, Y.; Sabatini, D.M. mTORC1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar H+-ATPase. Science 2011, 334, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. The Green Fluorescent Proteins. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef] [PubMed]
- Cranfill, P.J.; Sell, B.R.; Baird, M.A.; Allen, J.R.; Lavagnino, Z.; De Gruiter, H.M.; Kremers, G.; Davidson, M.W.; Ustione, A.; Piston, D.W. Quantitative assessment of fluorescent proteins. Nat. Methods 2016, 13, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Takenaka, Y.; Yamaguchi, A.; Nishikawa, S.; Mizuno, H. A novel yellowish-green fluorescent protein from the marine copepod, Chiridius poppei, and its use as a reporter protein in HeLa cells. Gene 2006, 372, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Bomati, E.K.; Haley, J.E.; Noel, J.P.; Deheyn, D.D. Spectral and structural comparison between bright and dim green fluorescent proteins in Amphioxus. Sci. Rep. 2014, 4, 5469. [Google Scholar] [CrossRef] [PubMed]
- Matz, M.V.; Fradkov, A.F.; Labas, Y.A.; Savitsky, A.P.; Zaraisky, A.G.; Markelov, M.L.; Lukyanov, S.A. Fluorescent proteins from nonbioluminescent Anthozoa species. Nat. Biotechnol. 1999, 17, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, H.; Ma, Y.; Nakashima, R.; Sakurai, K.; Matsuda, T.; Nagai, T. Acid-Tolerant Monomeric GFP from Olindias formosa. Cell Chem. Biol. 2018, 25, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E.; Tour, O.; Palmer, A.E.; Steinbach, P.A.; Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. A monomeric red fluorescent protein. Proc. Natl. Acad. Sci. USA 2002, 99, 7877–7882. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.G.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, Y.; Wu, J.; Shaner, N.C.; Campbell, R.E. Engineering of mCherry variants with long Stokes shift, red-shifted fluorescence, and low cytotoxicity. PLoS ONE 2017, 12, e0171257. [Google Scholar] [CrossRef] [PubMed]
- Bindels, D.S.; Haarbosch, L.; Van Weeren, L.; Postma, M.; Wiese, K.E.; Mastop, M.; Aumonier, S.; Gotthard, G.; Royant, A.; Hink, M.A.; et al. mScarlet: A bright monomeric red fluorescent protein for cellular imaging. Nat. Methods 2016, 14, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jackson, W.C.; Steinbach, P.A.; Tsien, R.Y. Evolution of new nonantibody proteins via iterative somatic hypermutation. Proc. Natl. Acad. Sci. USA 2004, 101, 16745–16749. [Google Scholar] [CrossRef] [PubMed]
- Kredel, S.; Oswald, F.; Nienhaus, K.; Deuschle, K.; Röcker, C.; Wolff, M.; Heilker, R.; Nienhaus, G.U.; Wiedenmann, J. mRuby, a bright monomeric red fluorescent protein for labeling of subcellular structures. PLoS ONE 2009, 4, e4391. [Google Scholar] [CrossRef] [PubMed]
- Lam, A.J.; St-Pierre, F.; Gong, Y.; Marshall, J.D.; Cranfill, P.J.; Baird, M.A.; McKeown, M.R.; Wiedenmann, J.; Davidson, M.W.; Schnitzer, M.J.; et al. Improving FRET dynamic range with bright green and red fluorescent proteins. Nat. Methods 2012, 9, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Bajar, B.T.; Wang, E.S.; Lam, A.J.; Kim, B.B.; Jacobs, C.L.; Howe, E.S.; Davidson, M.W.; Lin, M.Z.; Chu, J. Improving brightness and photostability of green and red fluorescent proteins for live cell imaging and FRET reporting. Sci. Rep. 2016, 6, 20889. [Google Scholar] [CrossRef] [PubMed]
- Merzlyak, E.M.; Goedhart, J.; Shcherbo, D.; Bulina, M.E.; Shcheglov, A.S.; Fradkov, A.F.; Gaintzeva, A.; Lukyanov, K.A.; Lukyanov, S.; Gadella, T.W.J.; et al. Bright monomeric red fluorescent protein with an extended fluorescence lifetime. Nat. Methods 2007, 4, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Piatkevich, K.D.; Malashkevich, V.N.; Morozova, K.S.; Nemkovich, N.A.; Almo, S.C.; Verkhusha, V.V. Extended stokes shift in fluorescent proteins: Chromophore-protein interactions in a near-infrared TagRFP675 variant. Sci. Rep. 2013, 3, 1847. [Google Scholar] [CrossRef] [PubMed]
- Shemiakina, I.I.; Ermakova, G.V.; Cranfill, P.J.; Baird, M.A.; Evans, R.A.; Souslova, E.A.; Staroverov, D.B.; Gorokhovatsky, A.Y.; Putintseva, E.V.; Gorodnicheva, T.V.; et al. A monomeric red fluorescent protein with low cytotoxicity. Nat. Commun. 2012, 3, 1204–1207. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Hommer, O.; Griesbeck, O. Efficiently folding and circularly permuted variants of the Sapphire mutant of GFP. BMC Biotechnol. 2003, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piatkevich, K.D.; Hulit, J.; Subach, O.M.; Wu, B.; Abdulla, A.; Segall, J.E.; Verkhusha, V.V. Monomeric red fluorescent proteins with a large Stokes shift. Proc. Natl. Acad. Sci. USA 2010, 107, 5369–5374. [Google Scholar] [CrossRef] [PubMed]
- Piatkevich, K.D.; Malashkevich, V.M.; Almo, S.C.; Verkhusha, V.V. Engineering ESPT pathway based on structural analysis of LSSmKate red fluorescent proteins with large Stokes shift. J. Am. Chem. Soc. 2010, 132, 10762–10770. [Google Scholar] [CrossRef] [PubMed]
- Tomosugi, W.; Matsuda, T.; Tani, T.; Nemoto, T.; Kotera, I.; Saito, K.; Horikawa, K.; Nagai, T. An ultramarine fluorescent protein with increased photostability and pH insensitivity. Nat. Methods 2009, 6, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Subach, O.M.; Cranfill, P.J.; Davidson, M.W.; Verkhusha, V.V. An enhanced monomeric blue fluorescent protein with the high chemical stability of the chromophore. PLoS ONE 2011, 6, e28674. [Google Scholar] [CrossRef] [PubMed]
- Katayama, H.; Yamamoto, A.; Mizushima, N.; Yoshimori, T.; Miyawaki, A. GFP-like Proteins Stably Accumulate in Lysosomes. Cell Struct. Funct. 2008, 33, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goedhart, J.; Von Stetten, D.; Noirclerc-Savoye, M.; Lelimousin, M.; Joosen, L.; Hink, M.A.; Van Weeren, L.; Gadella, T.W.J.; Royant, A. Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nat. Commun. 2012, 3, 751. [Google Scholar] [CrossRef] [PubMed]
- Markwardt, M.L.; Kremers, G.J.; Kraft, C.A.; Ray, K.; Cranfill, P.J.C.; Wilson, K.A.; Day, R.N.; Wachter, R.M.; Davidson, M.W.; Rizzo, M.A. An improved cerulean fluorescent protein with enhanced brightness and reduced reversible photoswitching. PLoS ONE 2011, 6, e17896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, H.; Henderson, J.N.; Remington, S.J.; Campbell, R.E. Directed evolution of a monomeric, bright and photostable version of Clavularia cyan fluorescent protein: Structural characterization and applications in fluorescence imaging. Biochem. J. 2006, 400, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Sawano, A.; Miyawaki, A. Directed evolution of green fluorescent protein by a new versatile PCR strategy for site-directed and semi-random mutagenesis. Nucleic Acids Res. 2000, 28, e78. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Patterson, G.H.; Davidson, M.W. Advances in fluorescent protein technology. J. Cell Sci. 2007, 120, 4247–4260. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.M.; Rudolf, F.; Meyer, A.; Pellaux, R.; Whitehead, E.; Panke, S.; Held, M. Identification and Characterisation of a pH-stable GFP. Sci. Rep. 2016, 6, 28166. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Ibata, K.; Park, E.S.; Kubota, M.; Mikoshiba, K.; Miyawaki, A. A variant of YFP with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 2002, 20, 1585–1588. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Lambert, G.G.; Chammas, A.; Ni, Y.; Cranfill, P.J.; Baird, M.A.; Sell, B.R.; Allen, J.R.; Day, R.N.; Israelsson, M.; et al. A bright monomeric green fluorescent protein derived from Branchiostoma lanceolatum. Nat. Methods 2013, 10, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, H.; Karasawa, S.; Okamura, Y.; Miyawaki, A. Improving membrane voltage measurements using FRET with new fluorescent proteins. Nat. Methods 2008, 5, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Dana, H.; Abdelfattah, A.S.; Patel, R.; Shea, J.; Molina, R.S.; Rawal, B.; Rancic, V.; Chang, Y.F.; Wu, L.; et al. A genetically encoded Ca2+ indicator based on circularly permutated sea anemone red fluorescent protein eqFP578. BMC Biol. 2018, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Lin, M.Z.; McKeown, M.R.; Steinbach, P.A.; Hazelwood, K.L.; Davidson, M.W.; Tsien, R.Y. Improving the photostability of bright monomeric orange and red fluorescent proteins. Nat. Methods 2008, 5, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Meurer, M.; Raghavan, S.; Rebane, A.; Lindquist, J.R.; Santos, S.; Kats, I.; Davidson, M.W.; Mazitschek, R.; Hughes, T.E.; et al. Live-cell multiphoton fluorescence correlation spectroscopy with an improved large Stokes shift fluorescent protein. Mol. Biol. Cell 2015, 26, 2054–2066. [Google Scholar] [CrossRef] [PubMed]
- Stiel, A.C.; Trowitzsch, S.; Weber, G.; Andresen, M.; Eggeling, C.; Hell, S.W.; Jakobs, S.; Wahl, M.C. 1.8 Å bright-state structure of the reversibly switchable fluorescent protein Dronpa guides the generation of fast switching variants. Biochem. J. 2007, 402, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Evdokimov, A.G.; Pokross, M.E.; Egorov, N.S.; Zaraisky, A.G.; Yampolsky, I.V.; Merzlyak, E.M.; Shkoporov, A.N.; Sander, I.; Lukyanov, K.A.; Chudakov, D.M. Structural basis for the fast maturation of Arthropoda green fluorescent protein. EMBO Rep. 2006, 7, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Shinobu, A.; Agmon, N. The hole in the barrel: Water exchange at the GFP chromophore. J. Phys. Chem. B 2015, 119, 3464–3478. [Google Scholar] [CrossRef] [PubMed]
- Pletnev, S.; Gurskaya, N.G.; Pletneva, N.V.; Lukyanov, K.A.; Chudakov, D.M.; Martynov, V.I.; Popov, V.O.; Kovalchuk, M.V.; Wlodawer, A.; Dauter, Z.; et al. Structural basis for phototoxicity of the genetically encoded photosensitizer KillerRed. J. Biol. Chem. 2009, 284, 32028–32039. [Google Scholar] [CrossRef] [PubMed]
- Verkhusha, V.V.; Akovbian, N.A.; Efremenko, E.N.; Varfolomeyev, S.D.; Vrzheshch, P.V. Kinetic analysis of maturation and denaturation of DsRed, a coral- derived red fluorescent protein. Biochemistry 2001, 66, 1342–1351. [Google Scholar] [PubMed]
- Huang, L.; Pike, D.; Sleat, D.E.; Nanda, V.; Lobel, P. Potential pitfalls and solutions for use of fluorescent fusion proteins to study the lysosome. PLoS ONE 2014, 9, e88893. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.A.; Kress, B.T.; Lu, Y.; Chandler-Militello, D.; Benraiss, A.; Nedergaard, M. Fluorescent Ca2+ indicators directly inhibit the Na,K-ATPase and disrupt cellular functions. Sci. Signal 2018, 11, eaal2039. [Google Scholar] [CrossRef] [PubMed]
- Kuroki, S.; Yoshida, T.; Tsutsui, H.; Iwama, M.; Ando, R.; Michikawa, T.; Miyawaki, A.; Ohshima, T.; Itohara, S. Excitatory Neuronal Hubs Configure Multisensory Integration of Slow Waves in Association Cortex. Cell Rep. 2018, 22, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Nick Pace, C.; Scholtz, J.M.; Grimsley, G.R. Forces stabilizing proteins. FEBS Lett. 2014, 588, 2177–2184. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tan, S.H.; Nicolas, V.; Bauvy, C.; Di Yang, N.; Zhang, J.; Xue, Y.; Codogno, P.; Shen, H.M. Activation of lysosomal function in the course of autophagy via mTORC1 suppression and autophagosome-lysosome fusion. Cell Res. 2013, 23, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.A.; Myers, J.T.; Swanson, J.A. pH-dependent regulation of lysosomal calcium in macrophages. J. Cell Sci. 2002, 115, 599–607. [Google Scholar] [PubMed]
- Steinberg, B.E.; Huynh, K.K.; Brodovitch, A.; Jabs, S.; Stauber, T.; Jentsch, T.J.; Grinstein, S. A cation counterflux supports lysosomal acidification. J. Cell Biol. 2010, 189, 1171–1186. [Google Scholar] [CrossRef] [PubMed]
- Benčina, M. Illumination of the Spatial Order of Intracellular pH by Genetically Encoded pH-Sensitive Sensors. Sensors 2013, 13, 16736–16758. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Rosendale, M.; Campbell, R.E.; Perrais, D. pHuji, a pH-sensitive red fluorescent protein for imaging of exo- and endocytosis. J. Cell Biol. 2014, 207, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Kogure, T.; Karasawa, S.; Araki, T.; Saito, K.; Kinjo, M.; Miyawaki, A. A fluorescent variant of a protein from the stony coral Montipora facilitates dual-color single-laser fluorescence cross-correlation spectroscopy. Nat. Biotechnol. 2006, 24, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Katayama, H.; Kogure, T.; Mizushima, N.; Yoshimori, T.; Miyawaki, A. A sensitive and quantitative technique for detecting autophagic events based on lysosomal delivery. Chem. Biol. 2011, 18, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Tantama, M.; Hung, Y.P.; Yellen, G. Imaging intracellular pH in live cells with a genetically encoded red fluorescent protein sensor. J. Am. Chem. Soc. 2011, 133, 10034–10037. [Google Scholar] [CrossRef] [PubMed]
- Serresi, M.; Bizzarri, R.; Cardarelli, F.; Beltram, F. Real-time measurement of endosomal acidification by a novel genetically encoded biosensor. Anal. Bioanal. Chem. 2009, 393, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Miesenböck, G.; De Angelis, D.A.; Rothman, J.E. Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature 1998, 394, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Disbrow, G.L.; Hanover, J.A.; Schlegel, R. Endoplasmic Reticulum-Localized Human Papillomavirus Type 16 E5 Protein Alters Endosomal pH but Not trans -Golgi pH Endoplasmic Reticulum-Localized Human Papillomavirus Type 16 E5 Protein Alters Endosomal pH but Not trans -Golgi pH. J. Virol. 2005, 79, 5839–5846. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, T.; Zhao, Y.; Nguyen, T.H.; Campbell, R.E.; Johnson, J.D. Fluorescent biosensors illuminate calcium levels within defined beta-cell endosome subpopulations. Cell Calcium 2015, 57, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Egashira, Y.; Takase, M.; Watanabe, S.; Ishida, J.; Fukamizu, A.; Kaneko, R.; Yanagawa, Y.; Takamori, S. Unique pH dynamics in GABAergic synaptic vesicles illuminates the mechanism and kinetics of GABA loading. Proc. Natl. Acad. Sci. USA 2016, 113, 10702–10707. [Google Scholar] [CrossRef] [PubMed]
- Dickson, E.J.; Duman, J.G.; Moody, M.W.; Chen, L.; Hille, B. Orai-STIM-mediated Ca2+ release from secretory granules revealed by a targeted Ca2+ and pH probe. Proc. Natl. Acad. Sci. USA 2012, 109, E3539–E3548. [Google Scholar] [CrossRef] [PubMed]
- Poëa-Guyon, S.; Pasquier, H.; Mérola, F.; Morel, N.; Erard, M. The enhanced cyan fluorescent protein: A sensitive pH sensor for fluorescence lifetime imaging. Anal. Bioanal. Chem. 2013, 405, 3983–3987. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.J.; Pinton, P.; Varadi, A.; Tacchetti, C.; Ainscow, E.K.; Pozzan, T.; Rizzuto, R.; Rutter, G.A. Dense core secretory vesicles revealed as a dynamic Ca2+ store in neuroendocrine cells with a vesicle-associated membrane protein aequorin chimaera. J. Cell Biol. 2001, 155, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Arosio, D.; Ricci, F.; Marchetti, L.; Gualdani, R.; Albertazzi, L.; Beltram, F. Simultaneous intracellular chloride and pH measurements using a GFP-based sensor. Nat. Methods 2010, 7, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Chen, S.; Shi, W.; Li, L.; Ma, H. Lysosomal pH rise during heat shock monitored by a lysosome-targeting near-infrared ratiometric fluorescent probe. Angew. Chem. Int. Ed. 2014, 53, 10916–10920. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zeng, Y.; Zhuang, X.; Sun, L.; Yao, X.; Pimpl, P.; Jiang, L. Organelle pH in the arabidopsis endomembrane system. Mol. Plant 2013, 6, 1419–1437. [Google Scholar] [CrossRef] [PubMed]
- Bagar, T.; Altenbach, K.; Read, N.D.; Benčina, M. Live-cell imaging and measurement of intracellular pH in filamentous fungi using a genetically encoded ratiometric probe. Eukaryot. Cell 2009, 8, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Gerasimenko, J.V.; Tepikin, A.V.; Petersen, O.H.; Gerasimenko, O.V. Calcium uptake via endocytosis with rapid release from acidifying endosomes. Curr. Biol. 1998, 8, 1335–1338. [Google Scholar] [CrossRef]
- Sherwood, M.W.; Prior, I.A.; Voronina, S.G.; Barrow, S.L.; Woodsmith, J.D.; Gerasimenko, O.V.; Petersen, O.H.; Tepikin, A.V. Activation of trypsinogen in large endocytic vacuoles of pancreatic acinar cells. Proc. Natl. Acad. Sci. USA 2007, 104, 5674–5679. [Google Scholar] [CrossRef] [PubMed]
- Rossier, J.; Tricoire, L.; Tsuzuki, K.; Courjean, O.; Gibelin, N.; Lambolez, B. Calcium dependence of aequorin bioluminescence dissected by random mutagenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 9500–9505. [Google Scholar] [CrossRef]
- Mahapatra, N.R.; Mahata, M.; Hazra, P.P.; Mcdonough, P.M.; Connor, D.T.O.; Mahata, S.K. A Dynamic Pool of Calcium in Catecholamine Storage Vesicles. J. Biol. Chem. 2004, 279, 51107–51121. [Google Scholar] [CrossRef] [PubMed]
- Gee, K.R.; Brown, K.A.; Chen, W.U.; Gray, D.; Johnson, I. Chemical and physiological characterization of fluo-4 Ca2+-indicator dyes. Cell Calcium 2000, 27, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Raveh, A.; Valitsky, M.; Shani, L.; Coorssen, J.R.; Blank, P.S.; Zimmerberg, J.; Rahamimoff, R. Cell Calcium Observations of calcium dynamics in cortical secretory vesicles. Cell Calcium 2012, 52, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Evans, E.; Morgan, A.J.; He, X.; Smith, D.A.; Elliot-Smith, E.; Sillence, D.J.; Churchill, G.C.; Schuchman, E.H.; Galione, A.; Platt, F.M. Niemann-Pick disease type C1 is a sphingosine storage disease that causes deregulation of lysosomal calcium. Nat. Med. 2008, 14, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, N.D.; Thiagarajah, J.R.; Verkman, A.S. Chloride Concentration in Endosomes Measured Using a Ratioable Fluorescent Cl− Indicator. J. Biol. Chem. 2002, 277, 5506–5513. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, N.D.; Verkman, A.S. Determinants of [Cl−] in recycling and late endosomes and Golgi complex measured using fluorescent ligands. J. Cell Biol. 2003, 160, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, S.; Fan, N.; Xiao, H.; Zhang, W.; Zhang, W.; Wang, H. Quantitative Fluorescence Ratio Imaging of Intralysosomal Chloride Ions with Single Excitation/Dual Maximum Emission. Chemistry 2014, 20, 11760–11767. [Google Scholar] [CrossRef] [PubMed]
- Vinkenborg, J.L.; Nicolson, T.J.; Bellomo, E.A.; Koay, M.S.; Rutter, G.A.; Merkx, M. Genetically encoded FRET sensors to monitor intracellular Zn2+ homeostasis. Nat. Methods 2009, 6, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Li, G.; Zhu, D.; Liu, Q.; Jiang, H. Rational Design of a Ratiometric and Targetable Fluorescent Probe for Imaging Lysosomal Zinc Ions. Inorg. Chem. 2012, 51, 10842–10849. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Fan, J.; Zhang, S.; Cao, J.; Song, K.; Ge, D.; Dong, H.; Wang, J.; Peng, X. Ratiometric fluorescence imaging of lysosomal Zn2+ release under oxidative stress in stem cells. Biomater. Sci. 2014, 89–97. [Google Scholar] [CrossRef]
- Lee, H.; Cho, C.; Seo, H.; Singha, S.; Jun, Y.W.; Lee, K.; Jung, Y.; Kim, K.; Park, S.; Bae, S.C.; et al. A two-photon fluorescent probe for lysosomal zinc ions. Chem. Commun. 2016, 52, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Llopis, J.; Heim, R.; McCaffery, J.M.; Adams, J.A.; Ikura, M.; Tsien, R.Y. Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 1997, 388, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Kanemaru, K.; Ishii, K.; Ohkura, M.; Okubo, Y.; Iino, M. Imaging intraorganellar Ca2+ at subcellular resolution using CEPIA. Nat. Commun. 2014, 5, 4153. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.J.; Baldwin, H.A.; Werley, C.A.; Boccardo, S.; Whitaker, L.R.; Yan, X.; Holt, G.T.; Schreiter, E.R.; Looger, L.L.; Cohen, A.E.; et al. A low affinity GCaMP3 variant (GCaMPer) for imaging the endoplasmic reticulum calcium store. PLoS ONE 2015, 10, e0139273. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Araki, S.; Wu, J.; Teramoto, T.; Chang, Y.; Nakano, M.; Abdelfattah, A.S.; Fujiwara, M.; Ishihara, T.; Nagai, T.; Campbell, R.E. An Expanded Palette of Genetically Encoded Ca2+ indicators. Science 2011, 557, 1888–1891. [Google Scholar] [CrossRef] [PubMed]
- Evers, T.H.; Appelhof, M.A.M.; de Graaf-Heuvelmans, P.T.H.M.; Meijer, E.W.; Merkx, M. Ratiometric Detection of Zn(II) Using Chelating Fluorescent Protein Chimeras. J. Mol. Biol. 2007, 374, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Betolngar, D.B.; Erard, M.; Pasquier, H.; Bousmah, Y.; Diop-Sy, A.; Guiot, E.; Vincent, P.; Mérola, F. pH sensitivity of FRET reporters based on cyan and yellow fluorescent proteins. Anal. Bioanal. Chem. 2015, 407, 4183–4193. [Google Scholar] [CrossRef] [PubMed]
- Schö, G. Calcium Signals from the Vacuole. Plants 2013, 2, 589–614. [Google Scholar] [CrossRef]
- Bischof, H.; Rehberg, M.; Stryeck, S.; Artinger, K.; Eroglu, E.; Waldeck-Weiermair, M.; Gottschalk, B.; Rost, R.; Deak, A.T.; Niedrist, T.; et al. Novel genetically encoded fluorescent probes enable real-time detection of potassium in vitro and in vivo. Nat. Commun. 2017, 8, 1422. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wu, S.; Rancic, V.; Qian, Y.; Miyashita, S. Genetically encoded ratiometric indicators for potassium ion. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lindenburg, L.H.; Vinkenborg, J.L.; Oortwijn, J.; Aper, S.J.A.; Merkx, M. MagFRET: The First Genetically Encoded Fluorescent Mg2+ Sensor. PLoS ONE 2013, 8, e82009. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, K.; Matsuda, T.; Shindo, Y.; Imamura, H.; Tamura, S.; Imai, R.; Kawakami, S.; Nagashima, R.; Soga, T.; Noji, H.; et al. A transient Rise in Free Mg2+ Ions Released from ATP-Mg Hydrolysis Contributes to Mitotic Chromosome Condensation. Curr. Biol. 2018, 28, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Hires, S.A.; Zhu, Y.; Tsien, R.Y. Optical measurement of synaptic glutamate spillover and reuptake by linker optimized glutamate-sensitive fluorescent reporters. Proc. Natl. Acad. Sci. USA 2008, 105, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Marvin, J.S.; Borghuis, B.G.; Tian, L.; Cichon, J.; Harnett, M.T.; Akerboom, J.; Gordus, A.; Renninger, S.L.; Chen, T.W.; Bargmann, C.I.; et al. An optimized fluorescent probe for visualizing glutamate neurotransmission. Nat. Methods 2013, 10, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Helassa, N.; Dürst, C.D.; Coates, C.; Kerruth, S.; Arif, U.; Schulze, C.; Wiegert, J.S.; Geeves, M.; Oertner, T.G.; Török, K. Ultrafast glutamate sensors resolve high-frequency release at Schaffer collateral synapses. Proc. Natl. Acad. Sci. USA 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bogner, M.; Ludewig, U. Visualization of arginine influx into plant cells using a specific FRET-sensor. J. Fluoresc. 2007, 17, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Jones, E.; Inoki, K. Lysosomal regulation of mTORC1 by amino acids in mammalian cells. Biomolecules 2017, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nagai, T. Recent progress in expanding the chemiluminescent toolbox for bioimaging. Curr. Opin. Biotechnol. 2017, 48, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Valiadi, M.; Iglesias-Rodriguez, D. Understanding Bioluminescence in Dinoflagellates—How Far Have We Come? Microorganisms 2013, 1, 3–25. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Color | Protein | λex/em a (nm) | ε b (103 M−1 cm−1) | QY c | Bright-ness d | Oligomeric State e (In Vitro/OSER) | pKa f | Resistance to Lysosomal Enzymes | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Blue | Sirius | 355/424 | 15.0 | 0.24 | 3.6 | mono/N.D. | <3.0 | Yes g | [27] |
| mTagBFP2 | 399/454 | 50.6 (76.0) | 0.64 (0.48) | 32.4 (36.5) | mono/oligo | 2.7 (2.4) | eqFP578-derivatives. Maybe no. | [28] | |
| Cyan | mTurquoise2 | 434/474 | 30.0 (31.0) | 0.93 (0.92) | 27.9 (28.5) | mono/mono | 3.1 (3.6) | avGFP-derivatives. Maybe no. [29] h | [30] |
| mCerulean3 | 434/475 | 40.0 (29.0) | 0.80 (0.80) | 32.0 (23.2) | mono/mono | 3.2 (3.4) | [31] | ||
| mTFP1 | 462/492 | 64.0 (53.0) | 0.85 (0.85) | 54.4 (45.1) | mono/mono | 4.3 (4.3) | N.D. | [32] | |
| UV-excitable Green | mT-Sapphire | 399/511 | 44.0 (34.0) | 0.60 (0.59) | 26.4 (20.1) | mono/mono | 4.9 (4.8) | avGFP-derivatives. Basically no. | [24] |
| Green | ECGFP | 463/506 | 23.9 | 0.14 | 3.3 | mono/N.D. | <4.0 | [33] | |
| mEmerald | 487/509 | 57.5 (62.0) | 0.68 (0.75) | 39.1 (46.5) | mono/mono | 6.0 (4.6) | [34] | ||
| pH-tdGFP | 488/515 | N.D. | N.D. | N.D. | tandem-dimer | 4.8 | [35] | ||
| Yellow-Green | mVenus | 515/527 | 105.0 (127.0) | 0.64 (0.67) | 67.2 (85.1) | mono/mono | 6.0 (5.4) | [36] | |
| Gamillus | 504/519 | 83.0 | 0.90 | 74.7 | mono/mono | 3.4 | Yes | [12] | |
| mNeonGreen | 506/517 | 116.0 (113.0) | 0.80 (0.80) | 92.8 (90.4) | mono/mono | 5.7 (5.4) | Maybe no [12] | [37] | |
| Orange | mKOk | 551/563 | 105.0 | 0.61 | 64.1 | mono/N.D. | 4.2 | N.D. | [38] |
| Red | mRFP1 | 584/607 | 50.0 (55.0) | 0.25 (0.35) | 12.5 (19.3) | mono/mono | 4.5 (3.8) | DsRed-derivatives. Basically yes. [39] | [13] |
| mCherry | 587/610 | 72.0 (85.0) | 0.22 (0.30) | 15.8 (25.5) | mono/mono | <4.5 (3.8) | [14] | ||
| mCherry2 | 589/610 | 79.4 | 0.22 | 17.5 | mono/N.D. | 3.3 | [15] | ||
| mStrawberry | 574/596 | 90.0 | 0.29 | 26.1 | mono/mono | <4.5 | [14] | ||
| mScarlet | 569/594 | 100.0 | 0.70 | 70.0 | mono/mono | 5.3 | [16] | ||
| mRuby | 558/605 | 112.0 (109.0) | 0.35 (0.38) | 39.2 (41.4) | mono/mono | 4.4 (4.4) | eqFP611-derivatives. Basically yes. [39] | [18] | |
| mRuby2 | 559/600 | 113.0 (107.0) | 0.38 (0.37) | 42.9 (39.6) | mono/mono | 5.3 (4.4) | [19] | ||
| mRuby3 | 558/592 | 128.0 | 0.45 | 57.6 | mono/N.D. | 4.8 | [20] | ||
| TagRFP | 555/584 | 100.0 (130.0) | 0.48 (0.33) | 48.0 (42.9) | mono/oligo | 3.1 (3.0) | eqFP578-derivatives. Basically no. [39] | [21] | |
| TagRFP-T | 555/584 | 81.0 (106.0) | 0.41 (0.32) | 33.2 (33.9) | mono/oligo | 4.6 (4.3) | [40] | ||
| FusionRed | 580/608 | 83.0 (85.0) | 0.19 (0.30) | 15.8 (25.5) | mono/mono | 4.6 (4.2) | [23] | ||
| Blue-excitable Red | LSSmKate1 | 463/624 | 31.2 | 0.08 | 2.5 | mono/N.D. | 3.2 | [25] | |
| LSSmKate2 | 460/605 | 26.0 | 0.17 | 4.4 | mono/N.D. | 2.7 | [25] | ||
| hmKeima4.15 | 436/612 | 28.0 | 0.29 | 8.1 | mono/N.D. | 5.2 | mKeima-derivative. Maybe yes. | [41] | |
| Far Red | mPlum | 590/649 | 41.0 (80.0) | 0.10 (0.13) | 4.1 (10.4) | mono/mono | <4.5 (4.6) | DsRed-derivatives. Maybe yes. | [17] |
| mRaspberry | 598/625 | 86.0 | 0.15 | 12.9 | mono/mono | <4.5 | [17] | ||
| TagRFP657 | 611/657 | 34.0 | 0.10 | 3.4 | mono/N.D. | 3.4 | eqFP578-derivatives. Maybe no. | [22] |
| Ion | Organelle | Cell | Sensor | Class | pKa or Kd (pH or pKa) | Resting pH, or [Ca2+, Zn2+ or Cl−] (pH) | Ref. |
|---|---|---|---|---|---|---|---|
| pH | Endosome | Foreskin keratino-cyte | Cellubrevin-r-pHluorin | Gen a, Single FP-base dual-excitation | 6.9 [61] | Early endosome: 5.9 | [62] |
| HeLa | Tat-E1GFP | Gen, Single FP-base dual-excitation | 6.4–6.7 | Early endosome: 6.8 Endosome: 5.8–6.3 | [60] | ||
| MIN6 beta-cell | TiVAMP-mKeima | Gen, Single FP-base dual-excitation | 5.8 | Early endosome: 6.3 Late endosome: 5.8 | [63] | ||
| Synaptic vesicle (SV) | Hippocam-pal neuron | Synaptophysin-mOrange2 | Gen, Single FP-base, intensiometric | 6.5 | GABAergic SV: ~6.4 Glutamatergic SV: ~5.8 | [64] | |
| Secretory granule | PC12 | Citrine (YFP) used in D1-SG (Ca2+ sensor) | Gen, intensiometric | 5.8 | 5.8 | [65] | |
| PC12 | CgA-ECFP | Gen, Single FP-base, FLIM | N.D. | 5.5 | [66] | ||
| MIN6 | VAMP2-pH.fluorin(e) | Gen, Single FP-base, Intensiometric | N.D. | 6.3 | [67] | ||
| PC12, WSS-1 | NPY-ClopHensor | Gen, Ratiometric (Non-FRET-base) | 6.8 | 5.2 (PC12) 5.6 (WSS1) | [68] | ||
| Lysosome | HeLa, MCF-7 | Lyso-pH | Chem b, Ratiometric | 5.0 | 4.6 | [69] | |
| Vacuole | A. thaliana | Aleurain-PRpHluorin | Gen, Single FP-base dual-excitation | 6.6 | 5.2 | [70] | |
| A. niger | RaVC (improved version of pHluorin for pH imaging in filamentous fungi) | Gen, Single FP-base dual-excitation | 6.7 (in vitro) 6.9 (in cell) | 6.2–6.5 | [71] | ||
| Ca2+ | Endosome | MIN6 beta-cell | TiVAMP-GEM-GECO1 | Gen, Single FP-base dual-emission | 0.27 (pH 7.5), 0.46 (7.0), 0.68 (6.5), 3.1 (6.0), 17.9 (5.4), 54.2 μM (5.0) | Early endosome: 0.5 (6.3) Late endosome: 2.5 μM (5.8) | [63] |
| 3T3 Swiss fibroblast | Oregon green 488 BAPTA-5N | Chem, Intensiometric | 20 μM | 3.0 μM (5.7) (30 min incubation) | [72] | ||
| Pancreatic acinar cell | Oregon green 488 BAPTA-5N | Chem, Intensiometric | 36.5 (pH 7.2), 55.3 (5.9), 116 μM (5.3) | 37 μM (5.9) | [73] | ||
| Secretory granule | PC12 | D1-SG | Gen, FRET-base | 120–190 μM (pH 7.4–5.5) | 69 μM (5.8) | [65] | |
| MIN6 | VAMP2-mut.aequorin | Gen, Intensiometric | 1–10 μM | ~50 μM (6.3) | [67] | ||
| PC12 | Chromogranin-aequorin | Gen, Intensiometric | 3.8 μM [74] | 1.4 μM (~5.5) | [75] | ||
| Sea urchin egg | Fluo-4 | Chem, Intensiometric | 345 nM (pH 7.2) [76] | ~10–100 μM | [77] | ||
| Lysosome | Mouse macrophage | Fura-2 dextran | Chem, Ratiometric | ~200 μM | 600 μM (4.5) | [53] | |
| Mouse macrophage | Oregon green BAPTA-1 dextran | Chem, Intensiometric | ~500 μM | 400 μM (4.5) | [53] | ||
| Human fibroblast | Rhod dextran | Chem, Intensiometric | 551 ± 107 μM | 550 μM (4.5) | [78] | ||
| Cl− | Endosome | J774 cell, CHO cell | BAC-TMR-dextran | Chem, Ratiometric | ~25–50 mM (pH 7.4) | 17→53 mM (6.95→5.30) (J773) 28→73 mM (6.92→5.60) (CHO) | [79] |
| J774 cell, CHO cell | BAC-dextran-Tf-TMR | Chem, Ratiometric | ~25–50 mM (pH 7.4) | 18→40 mM (6.91→6.05) (J773) 24→46 mM (6.95→6.18) (CHO) | [80] | ||
| Secretory granule | PC12, WSS-1 | NPY-ClopHensor | Gen, Ratiometric (Non-FRET-base) | 13.1 mM (pKa 6.81) | 110 ± 48 mM (5.2) (PC12) 122 mM (5.6) (WSS1) | [68] | |
| Lysosome | Venticular myocyte | 6-methoxyquino-linium–dansyl | Chem, Ratiometric | ~15 mM (pH 4.5) | 9.46 mM | [81] | |
| Zn2+ | Secretory granule | INS-1 (832/13) | VAMP2-eZinCh | Gen, Ratiometric (FRET) | 8 (pH 7.1), 260 μM (6.0) | 1–100 μM | [82] |
| Lysosome | NIH 3T3 | DQZn4 | Chem, Ratiometric | 16 nM (pH 5.2) | N.D. | [83] | |
| NSCs, MCF-7, HeLa | LysoZn-1 | Dhem, Ratiometric | ~150 μM (pH 7.2) | N.D. | [84] | ||
| NIH 3T3 | probe 1 (no name) | Chem, Intensiometric | 8.55 μM (pH 5.0) | N.D. | [85] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinoda, H.; Shannon, M.; Nagai, T. Fluorescent Proteins for Investigating Biological Events in Acidic Environments. Int. J. Mol. Sci. 2018, 19, 1548. https://doi.org/10.3390/ijms19061548
Shinoda H, Shannon M, Nagai T. Fluorescent Proteins for Investigating Biological Events in Acidic Environments. International Journal of Molecular Sciences. 2018; 19(6):1548. https://doi.org/10.3390/ijms19061548
Chicago/Turabian StyleShinoda, Hajime, Michael Shannon, and Takeharu Nagai. 2018. "Fluorescent Proteins for Investigating Biological Events in Acidic Environments" International Journal of Molecular Sciences 19, no. 6: 1548. https://doi.org/10.3390/ijms19061548
APA StyleShinoda, H., Shannon, M., & Nagai, T. (2018). Fluorescent Proteins for Investigating Biological Events in Acidic Environments. International Journal of Molecular Sciences, 19(6), 1548. https://doi.org/10.3390/ijms19061548