Decoding the Divergent Subcellular Location of Two Highly Similar Paralogous LEA Proteins

 and
and
Abstract
:1. Introduction
2. Results
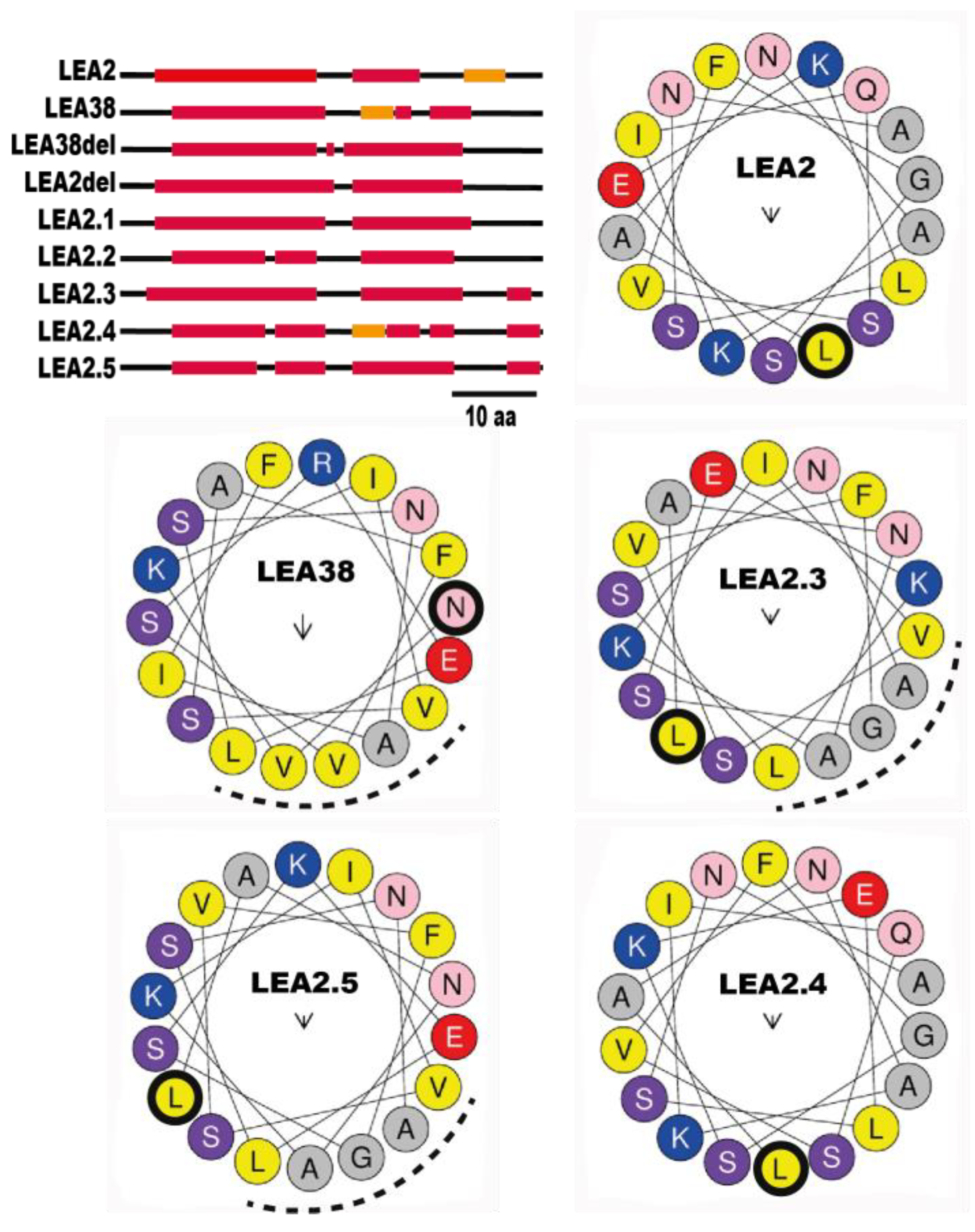
2.1. Proteins of the LEA_3 Family Are Expected to Be Mitochondrial
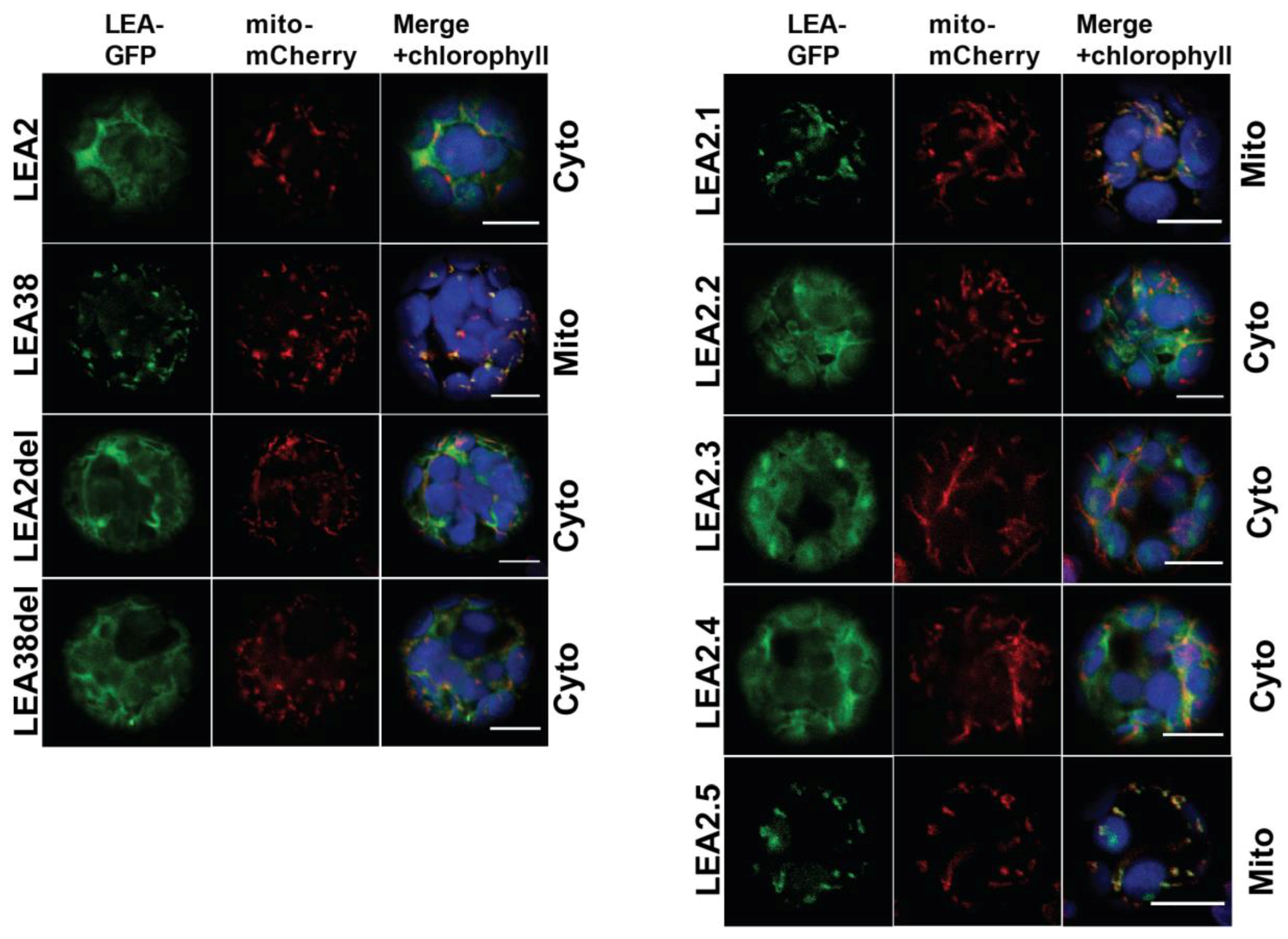
2.2. Determinants of LEA38 Mitochondrial Targeting
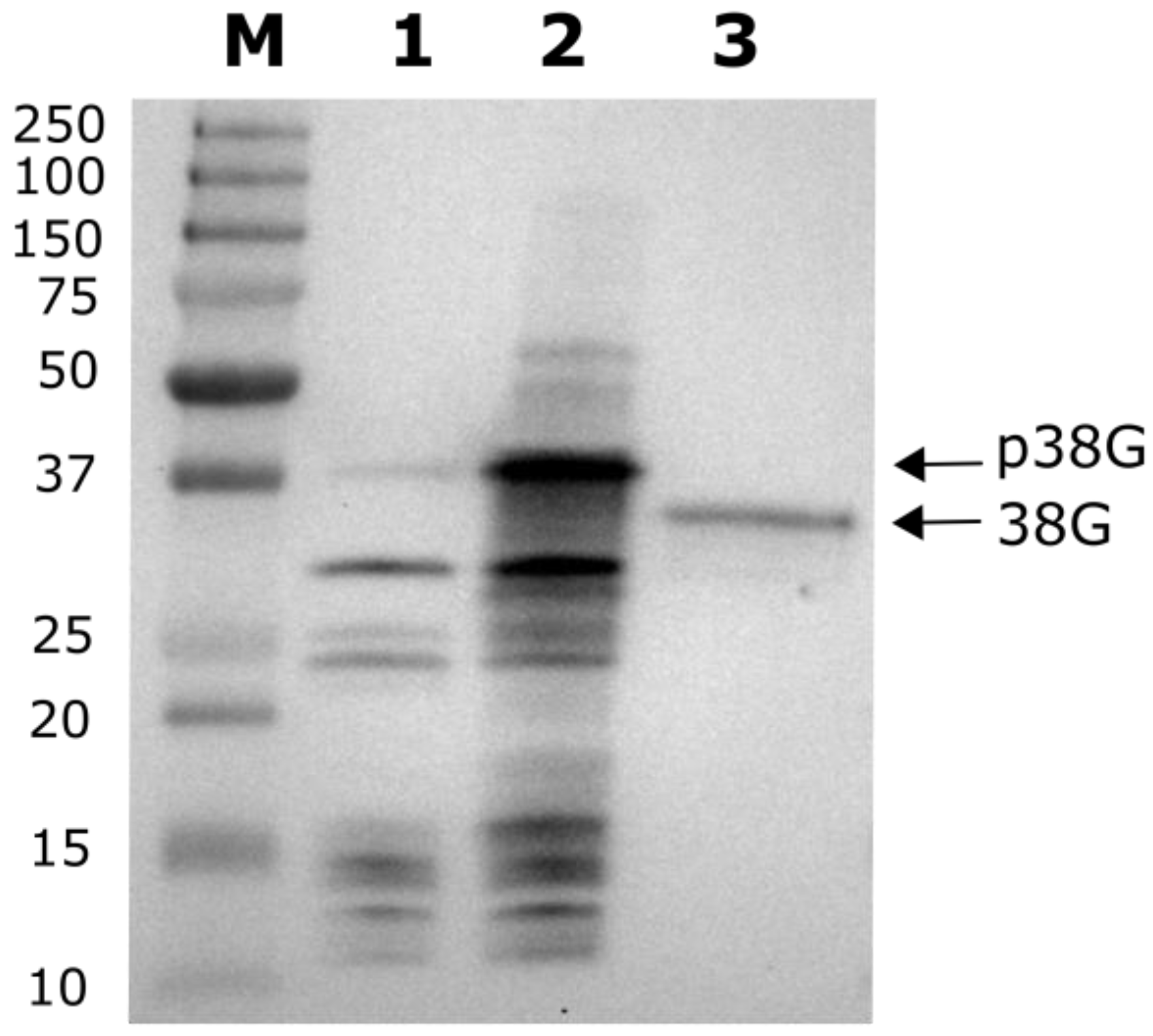
2.3. MTS Cleavage Site Determination for LEA38
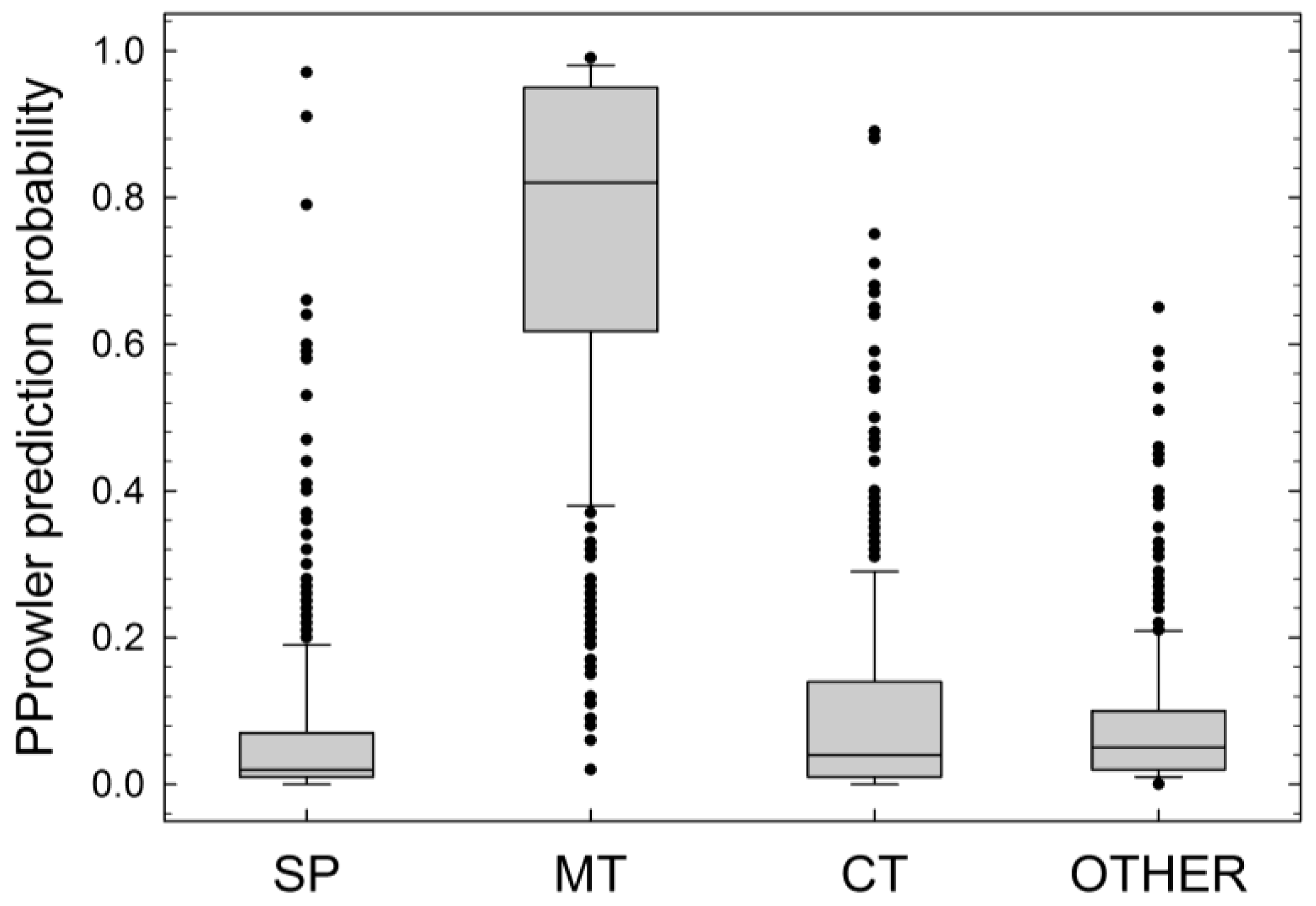
2.4. Comparison of Experimental Data and In Silico Predictions
3. Discussion
4. Materials and Methods
4.1. Plant Culture, Protoplast Isolation, and Transformation
4.2. Expression of Mutated Proteins
4.3. Microscopy
4.4. In Vitro Production of Recombinant Proteins
4.5. Crude Mitochondria Isolation, and N-Terminus Sequencing
4.6. Protein Analysis by Western Blot
4.7. Bioinformatics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MTS | Mitochondrial targeting sequence |
| GFP | Green fluorescent protein |
| LEA | Late Embryogenesis Abundant |
References
- Rao, R.S.P.; Salvato, F.; Thal, B.; Eubel, H.; Thelen, J.J.; Møller, I.M. The proteome of higher plant mitochondria. Mitochondrion 2017, 33, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Murcha, M.W.; Kmiec, B.; Kubiszewski-Jakubiak, S.; Teixeira, P.F.; Glaser, E.; Whelan, J. Protein import into plant mitochondria: Signals, machinery, processing, and regulation. J. Exp. Bot. 2014, 65, 6301–6335. [Google Scholar] [CrossRef] [PubMed]
- Bolender, N.; Sickmann, A.; Wagner, R.; Meisinger, C.; Pfanner, N. Multiple pathways for sorting mitochondrial precursor proteins. EMBO Rep. 2008, 9, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing mitochondrial proteins: Machineries and mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef] [PubMed]
- Pfanner, N.; Geissler, A. Versatility of the mitochondrial protein import machinery. Nat. Rev. Mol. Cell Biol. 2001, 2, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Mokranjac, D.; Neupert, W. Energetics of protein translocation into mitochondria. Biochim. Biophys. Acta Bioenerg. 2008, 1777, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, P.F.; Glaser, E. Processing peptidases in mitochondria and chloroplasts. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.J.; Hulett, J.M.; Likić, V.A.; Lithgow, T.; Gooley, P.R. Convergent evolution of receptors for protein import into mitochondria. Curr. Biol. 2006, 16, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Duncan, O.; Murcha, M.W.; Whelan, J. Unique components of the plant mitochondrial protein import apparatus. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, P. Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol. 2008, 179, 257–285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Glaser, E. Interaction of plant mitochondrial and chloroplast signal peptides with the Hsp70 molecular chaperone. Trends Plant Sci. 2002, 7, 14–21. [Google Scholar] [CrossRef]
- Xu, L.; Carrie, C.; Law, S.R.; Murcha, M.W.; Whelan, J. Acquisition, conservation, and loss of dual-targeted proteins in land plants. Plant Physiol. 2013, 161, 644–662. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Taylor, N.L.; Whelan, J.; Millar, A.H. Refining the definition of plant mitochondrial presequences through analysis of sorting signals, N-terminal modifications, and cleavage motifs. Plant Physiol. 2009, 150, 1272–1285. [Google Scholar] [CrossRef] [PubMed]
- Von Heijne, G.; Steppuhn, J.; Herrmann, R.G. Domain structure of mitochondrial and chloroplastic targeting peptides. Eur. J. Biochem. 1989, 180, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Peeters, N.; Small, I. Dual targeting to mitochondria and chloroplasts. Biochim. Biophys. Acta Mol. Cell Res. 2001, 1541, 54–63. [Google Scholar] [CrossRef]
- Roise, D.; Horvath, S.J.; Tomich, J.M.; Richards, J.H.; Schatz, G. A chemically synthesized pre-sequence of an imported mitochondrial protein can form an amphiphilic helix and perturb natural and artificial phospholipid bilayers. EMBO J. 1986, 5, 1327–1334. [Google Scholar] [PubMed]
- Saitoh, T.; Igura, M.; Obita, T.; Ose, T.; Kojima, R.; Maenaka, K.; Endo, T.; Kohda, D. Tom20 recognizes mitochondrial presequences through dynamic equilibrium among multiple bound states. EMBO J. 2007, 26, 4777–4787. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, S.; Kuhn, C.; Berglund, A.K.; Roth, C.; Glaser, E. The role of the N-terminal domain of chloroplast targeting peptides in organellar protein import and miss-sorting. FEBS Lett. 2006, 580, 3966–3972. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Sjöling, S.; Wallin, E.; Wrede, P.; Glaser, E.; Von Heijne, G. Feature-extraction from endopeptidase cleavage sites in mitochondrial targeting peptides. Proteins Struct. Funct. Bioinform. 1998, 30, 49–60. [Google Scholar] [CrossRef]
- Tanudji, M.; Sjöling, S.; Glaser, E.; Whelan, J. Signals required for the import and processing of the alternative oxidase into mitochondria. J. Biol. Chem. 1999, 274, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Kitada, S.; Yamasaki, E.; Kojima, K.; Ito, A. Determination of the cleavage site of the presequence by mitochondrial processing peptidase on the substrate binding scaffold and the multiple subsites inside a molecular cavity. J. Biol. Chem. 2003, 278, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Sjöling, S.; Tanudji, M.; Somogyi, L.; Andreu, D.; Göran Eriksson, L.E.; Gräslund, A.; Whelan, J.; Glaser, E. Mutagenesis and computer modelling approach to study determinants for recognition of signal peptides by the mitochondrial processing peptidase. Plant J. 2001, 27, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Staiger, C.; Hinneburg, A.; Klösgen, R.B. Diversity in degrees of freedom of mitochondrial transit peptides. Mol. Biol. Evol. 2009, 26, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Heazlewood, J.L.; Kristensen, B.K.; Braun, H.P.; Møller, I.M. The plant mitochondrial proteome. Trends Plant Sci. 2005, 10, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.H.; Macherel, D. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunnacliffe, A.; Hincha, D.K.; Leprince, O.; Macherel, D. LEA proteins: Versatility of form and function. In Sleeping Beauties—Dormancy and Resistance in Harsh Environments; Lubzens, E., Cerda, J., Clark, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Tolleter, D.; Jaquinod, M.; Mangavel, C.; Passirani, C.; Saulnier, P.; Manon, S.; Teyssier, E.; Payet, N.; Avelange-Macherel, M.H.; Macherel, D. Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 2007, 19, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Tolleter, D.; Hincha, D.K.; Macherel, D. A mitochondrial late embryogenesis abundant protein stabilizes model membranes in the dry state. Biochim. Biophys. Acta 2010, 1798, 1926–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avelange-Macherel, M.-H.; Payet, N.; Lalanne, D.; Neveu, M.; Tolleter, D.; Burstin, J.; Macherel, D. Variability within a pea core collection of LEAM and HSP22, two mitochondrial seed proteins involved in stress tolerance. Plant. Cell Environ. 2015, 38, 1299–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, L.M.; Gan, S.; Quirino, B.; Amasino, R.M. A comparison of the expression patterns of several senescence-associated genes in response to stress and hormone treatment. Plant Mol. Biol. 1998, 37, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Mowla, S.B.; Cuypers, A.; Driscoll, S.P.; Kiddle, G.; Thomson, J.; Foyer, C.H.; Theodoulou, F.L. Yeast complementation reveals a role for an Arabidopsis thaliana late embryogenesis abundant (LEA)-like protein in oxidative stress tolerance. Plant J. 2006, 48, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Salleh, F.M.; Evans, K.; Goodall, B.; Machin, H.; Mowla, S.B.; Mur, L.A.J.; Runions, J.; Theodoulou, F.L.; Foyer, C.H.; Rogers, H.J. A novel function for a redox-related LEA protein (SAG21/AtLEA5) in root development and biotic stress responses. Plant Cell Environ. 2012, 35, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Fournier-Level, A.; Korte, A.; Cooper, M.D.; Nordborg, M.; Schmitt, J.; Wilczek, A.M. A map of local adaptation in Arabidopsis thaliana. Science 2011, 334, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Shodai, T.; Muto, T.; Mihara, K.; Torii, H.; Nishikawa, S.; Endo, T.; Kohda, D. Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell 2000, 100, 551–560. [Google Scholar] [CrossRef]
- Duby, G.; Oufattole, M.; Boutry, M. Hydrophobic residues within the predicted N-terminal amphiphilic α-helix of a plant mitochondrial targeting presequence play a major role in in vivo import. Plant J. 2001, 27, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Liao, P.C.; Chuang, K.T.; Kao, M.C. Mitochondrial targeting of human NADH dehydrogenase (ubiquinone) flavoprotein 2 (NDUFV2) and its association with early-onset hypertrophic cardiomyopathy and encephalopathy. J. Biomed. Sci. 2011, 18, 29. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Nelson, C.J.; Li, L.; Taylor, N.L.; Ströher, E.; Petereit, J.; Millar, A.H. INTERMEDIATE CLEAVAGE PEPTIDASE55 modifies enzyme amino termini and alters protein stability in Arabidopsis mitochondria. Plant Physiol. 2015, 168, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Trelles, F.; Tarrío, R.; Ayala, F.J. Origins and evolution of spliceosomal introns. Annu. Rev. Genet. 2006, 40, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.D.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Zhang, J. Protein subcellular relocalization in the evolution of yeast singleton and duplicate genes. Genome Biol. Evol. 2009, 1, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, Y.; Lavrov, D.V.; Gu, X. Comparative study of human mitochondrial proteome reveals extensive protein subcellular relocalization after gene duplications. BMC Evol. Biol. 2009, 9, 275. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-L.; Pan, A.Q.; Adams, K.L. Protein subcellular relocalization of duplicated genes in Arabidopsis. Genome Biol. Evol. 2014, 6, 2501–2515. [Google Scholar] [CrossRef] [PubMed]
- Candat, A.; Poupart, P.; Andrieu, J.-P.; Chevrollier, A.; Reynier, P.; Rogniaux, H.; Avelange-Macherel, M.-H.; Macherel, D. Experimental determination of organelle targeting-peptide cleavage sites using transient expression of green fluorescent protein translational fusions. Anal. Biochem. 2013, 434, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Miller, W. A time-efficient, linear-space similarity algorithm. Adv. Appl. Math. 1991, 12, 337–357. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cowley, A.; Uludag, M.; Gur, T.; McWilliam, H.; Squizzato, S.; Park, Y.M.; Buso, N.; Lopez, R. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res. 2015, 43, W580–W584. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER web server: 2015 Update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef] [PubMed]
- Nakai, K.; Horton, P. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 1999, 24, 34–36. [Google Scholar] [CrossRef]
- Claros, M.G.; Vincens, P. Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur. J. Biochem. 1996, 241, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Small, I.; Peeters, N.; Legeai, F.; Lurin, C. Predotar: A tool for rapidly screening proteomes for N-terminal targeting sequences. Proteomics 2004, 4, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Bodén, M.; Hawkins, J. Prediction of subcellular localization using sequence-biased recurrent networks. Bioinformatics 2005, 21, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, Y.; Tsuji, J.; Fu, S.C.; Tomii, K.; Horton, P.; Imai, K. MitoFates: Improved prediction of mitochondrial targeting sequences and their cleavage sites. Mol. Cell. Proteom. 2015, 14, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Halligan, B.D.; Ruotti, V.; Jin, W.; Laffoon, S.; Twigger, S.N.; Dratz, E.A. ProMoST (Protein Modification Screening Tool): A web-based tool for mapping protein modifications on two-dimensional gels. Nucleic Acids Res. 2004, 32, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef] [PubMed]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific alpha-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation | Description |
|---|---|
| LEA2del | LEA2 without residues 33 to 37 (KTALD) |
| LEA38del | LEA38 without residues 33 to 43 (AQGSVSSGGRS) |
| LEA2.1 | LEA38 (residues 1 to 43) + LEA2 (residues 38 to 91) |
| LEA2.2 | LEA2 with (KTALD) replaced by (AQGSVSSGGRS) |
| LEA2.3 | Same as LEA2.2 with Q11V mutation |
| LEA2.4 | Same as LEA2.2 with inversion at positions 17 and 18 (EK into KE) |
| LEA2.5 | Same as LEA2.2 with Q11V mutation and inversion at position 17 and 18 |
| IPsort | MitoFates | MITOPROT II v1.101 | PProwler 1.2 | Predotar | TargetP 1.1 | |
|---|---|---|---|---|---|---|
| LEA2 | M | M (0.839) | M (0.958) | M (0.84) | M (0.43) | Ct (0.364) |
| - | 28 | 35 | - | - | 48 | |
| LEA38 | Ct | M (0.544) | M (0.986) | M (0.90) | M (0.31) | Ct (0.493) |
| - | 28 | 43 | - | - | 46 | |
| LEA2-GFP | M | M (0.889) | M (0.949) | M (0.85) | M (0.43) | M (0.357) |
| - | 28 | 35 | - | - | 28 | |
| LEA38-GFP | Ct | M (0.672) | M (0.995) | M (0.85) | M (0.31) | Ct (0.429) |
| - | 28 | 43 | - | - | 46 | |
| LEA38del-GFP | Ct | M (0.760) | M (0.984) | M (0.92) | M (0.32) | SP (0.499) |
| - | 28 | 28 | - | - | 13 | |
| LEA2.1-GFP | Ct | M (0.493) | M (0.984) | M (0.88) | M (0.31) | Ct (0.539) |
| - | 28 | 43 | - | - | 54 | |
| LEA2.5-GFP | M | M (0.500) | M (0.924) | M (0.73) | M (0.35) | Ct (0.657) |
| - | 28 | 43 | - | - | 54 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avelange-Macherel, M.-H.; Candat, A.; Neveu, M.; Tolleter, D.; Macherel, D. Decoding the Divergent Subcellular Location of Two Highly Similar Paralogous LEA Proteins. Int. J. Mol. Sci. 2018, 19, 1620. https://doi.org/10.3390/ijms19061620
Avelange-Macherel M-H, Candat A, Neveu M, Tolleter D, Macherel D. Decoding the Divergent Subcellular Location of Two Highly Similar Paralogous LEA Proteins. International Journal of Molecular Sciences. 2018; 19(6):1620. https://doi.org/10.3390/ijms19061620
Chicago/Turabian StyleAvelange-Macherel, Marie-Hélène, Adrien Candat, Martine Neveu, Dimitri Tolleter, and David Macherel. 2018. "Decoding the Divergent Subcellular Location of Two Highly Similar Paralogous LEA Proteins" International Journal of Molecular Sciences 19, no. 6: 1620. https://doi.org/10.3390/ijms19061620
APA StyleAvelange-Macherel, M. -H., Candat, A., Neveu, M., Tolleter, D., & Macherel, D. (2018). Decoding the Divergent Subcellular Location of Two Highly Similar Paralogous LEA Proteins. International Journal of Molecular Sciences, 19(6), 1620. https://doi.org/10.3390/ijms19061620