5-Azacitidine Induces Cell Death in a Tissue Culture of Brachypodium distachyon

 ,
, 
 ,
,  and
and 
Abstract
:
1. Introduction
2. Results
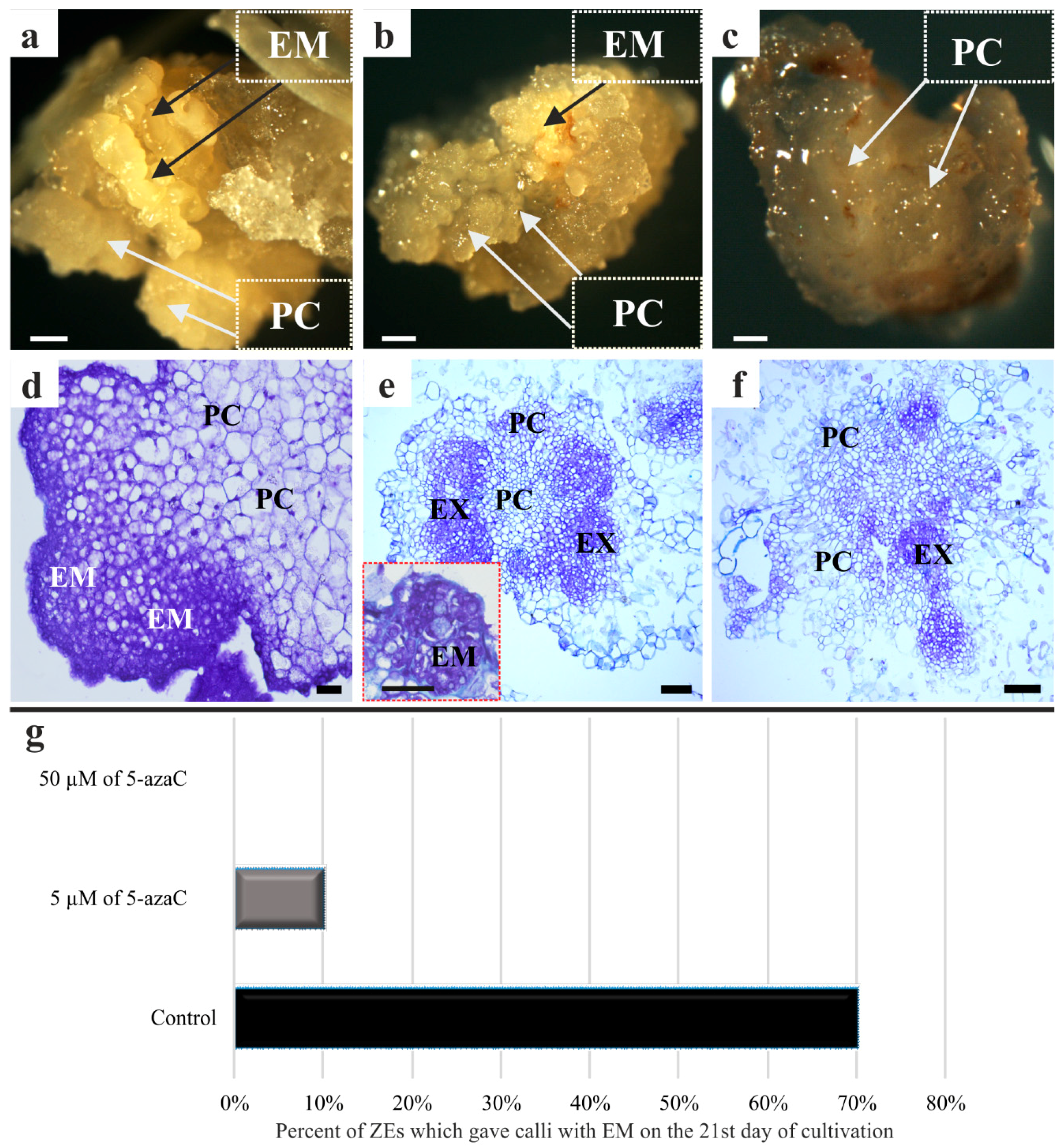
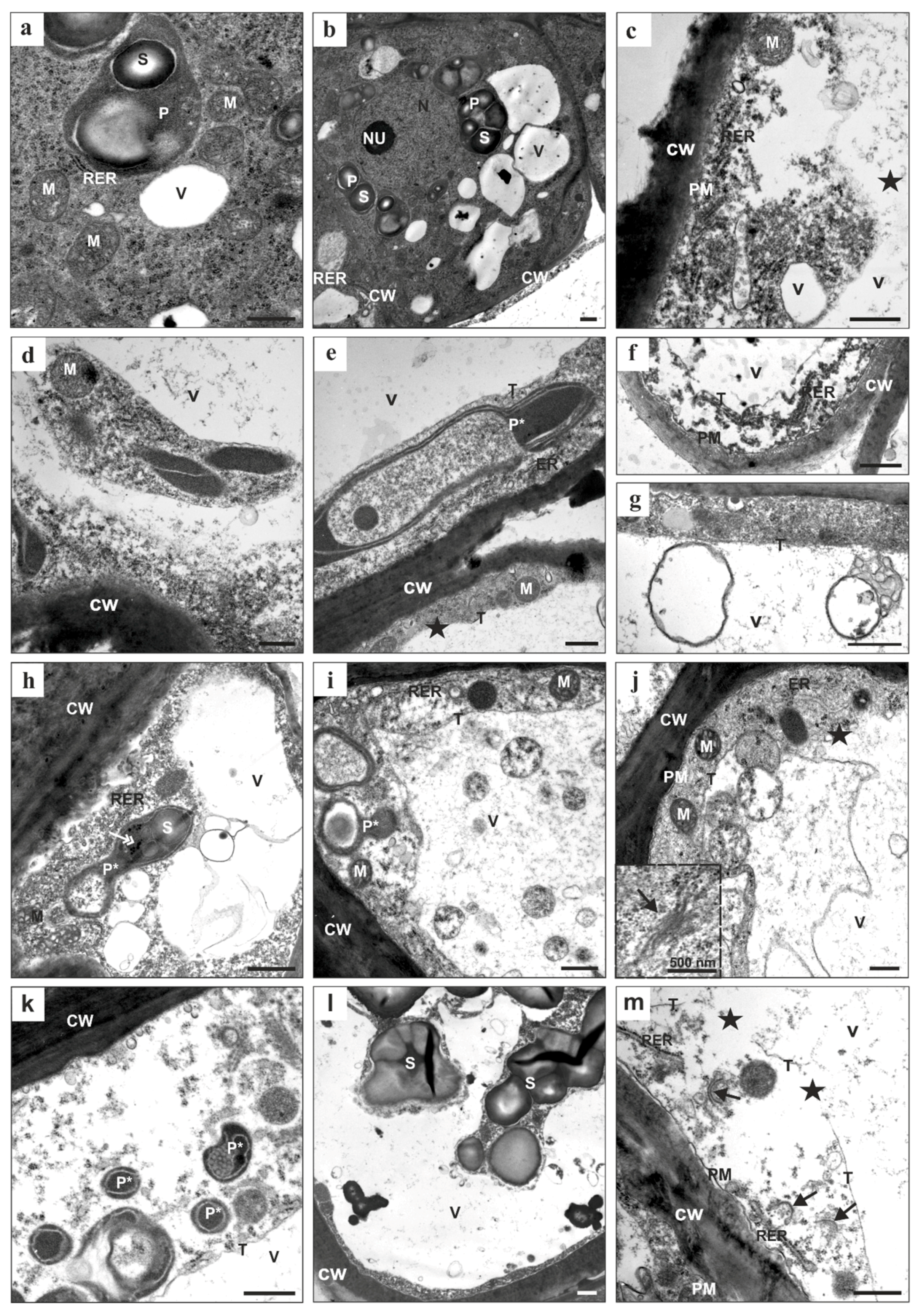
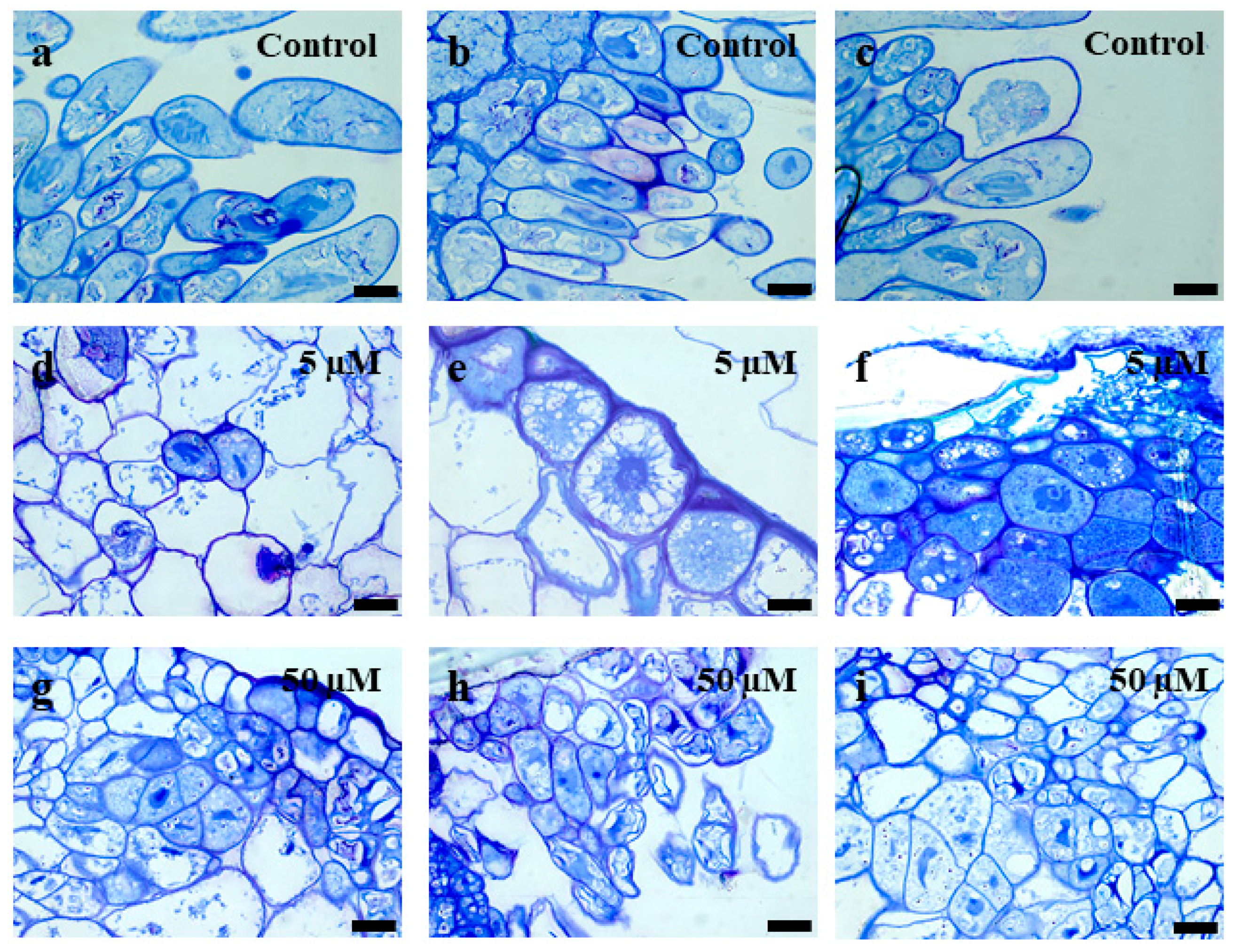
2.1. Morphological, Histological, Cytological, and Ultrastructural Observations in the Brachypodium Callus
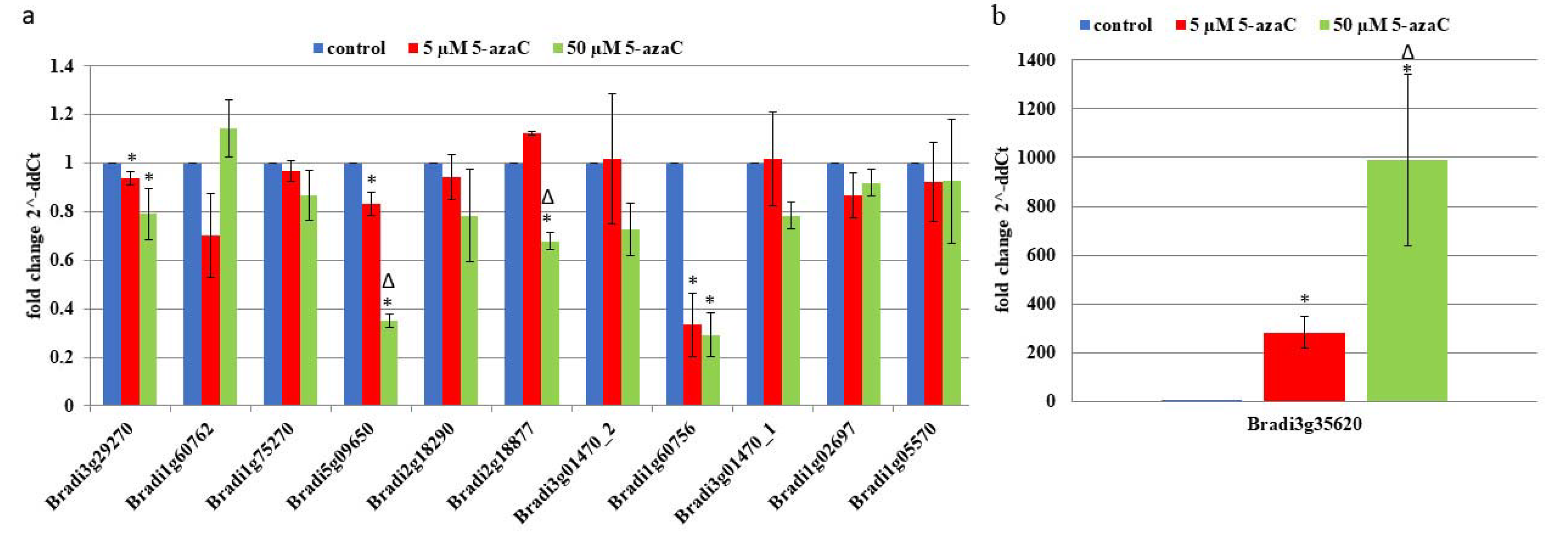
2.2. RT-PCR
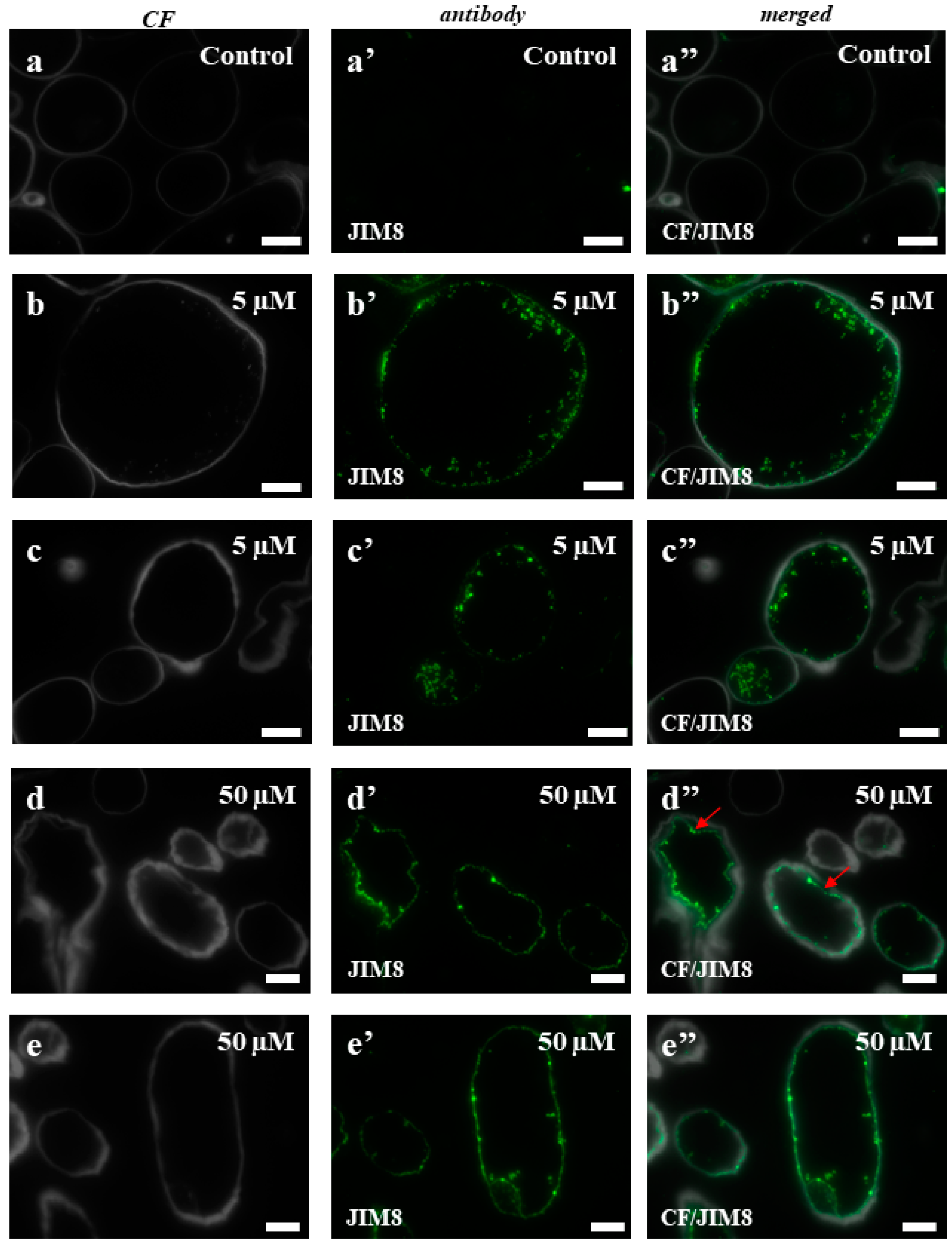
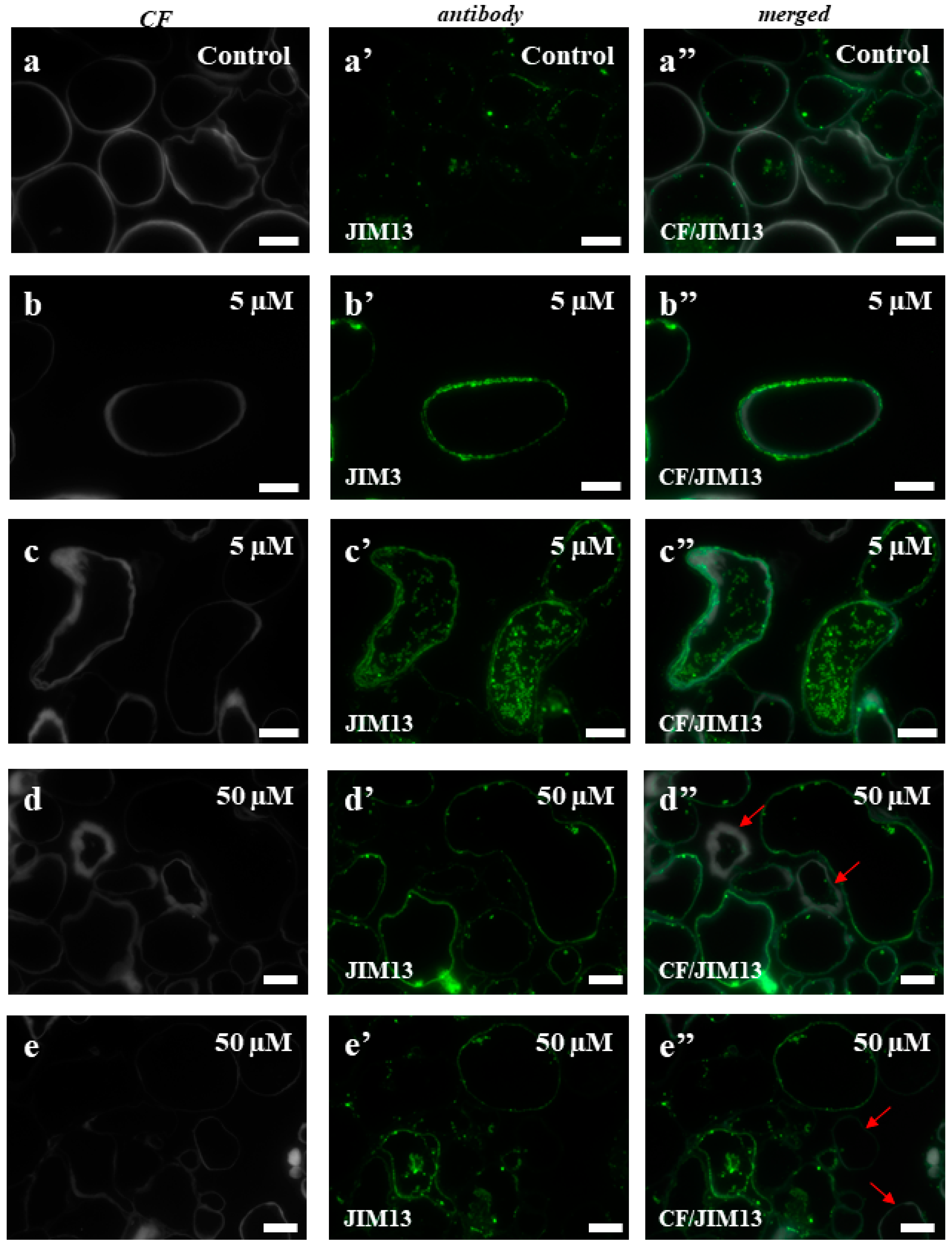
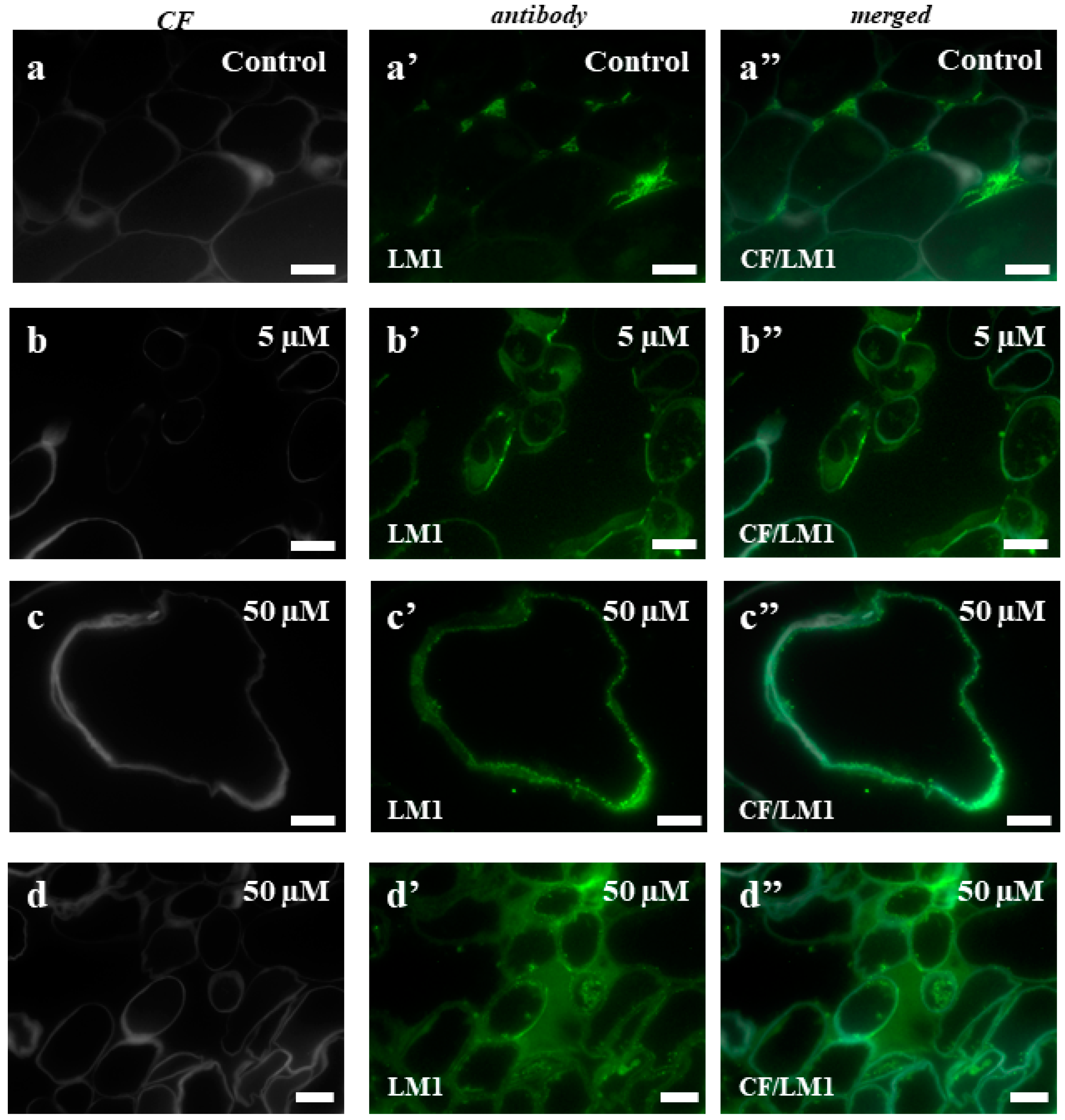
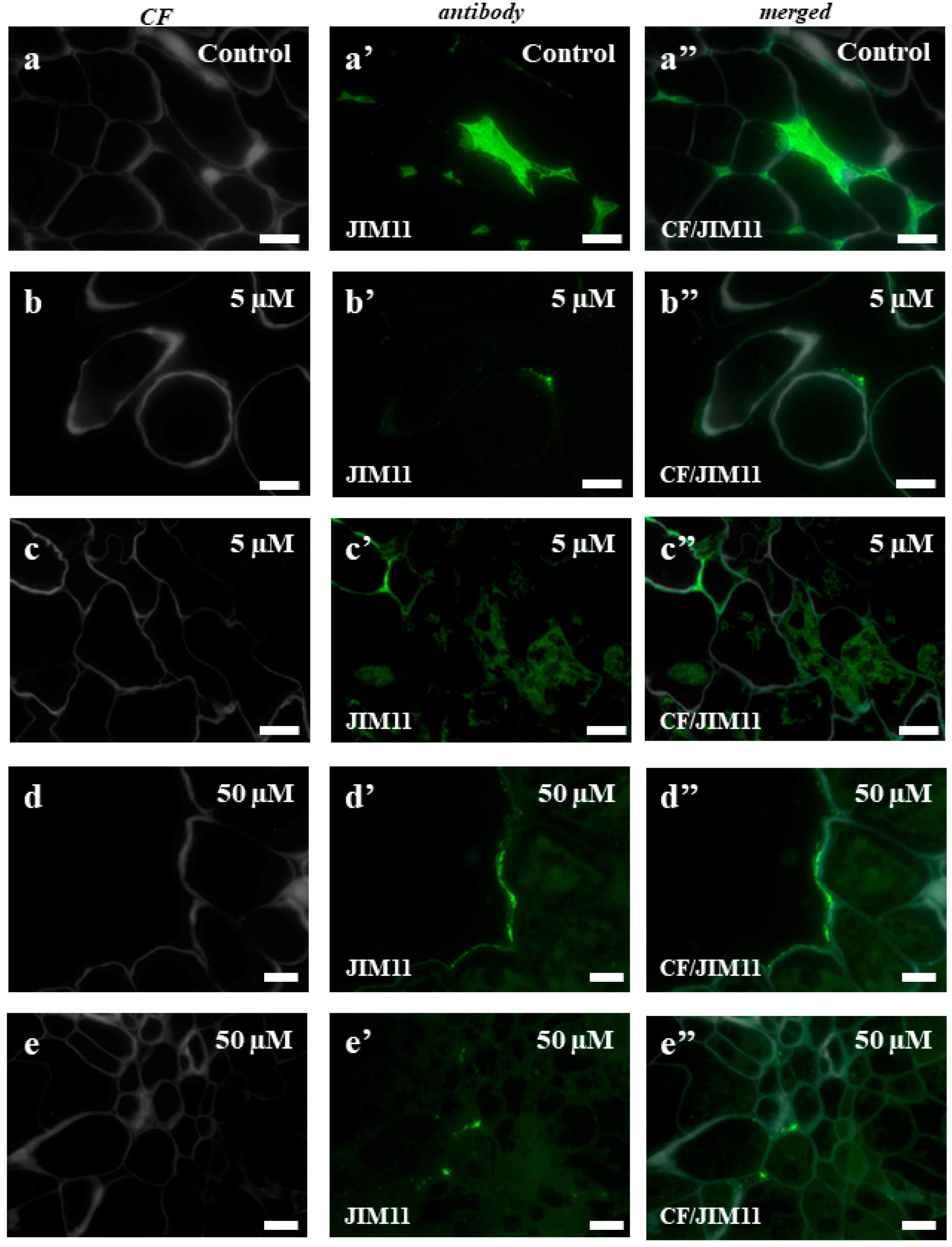
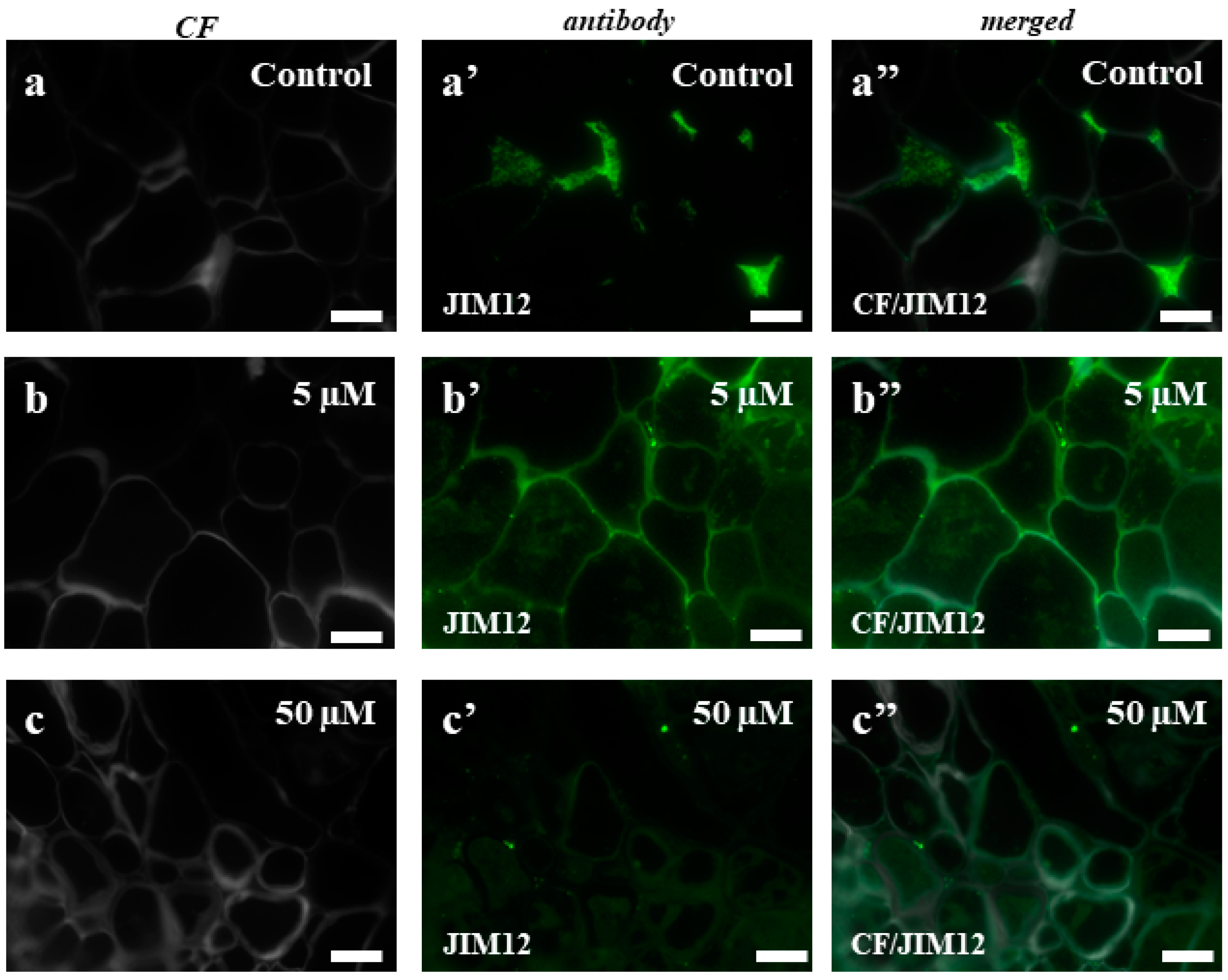
2.3. AGPs and Extensins under Different 5-azaC Treatments
3. Discussion
4. Materials and Methods
4.1. Plant Material Growth, In Vitro Culture Conditions, and 5-azaC Treatment
4.2. Histological Procedures
4.3. TEM
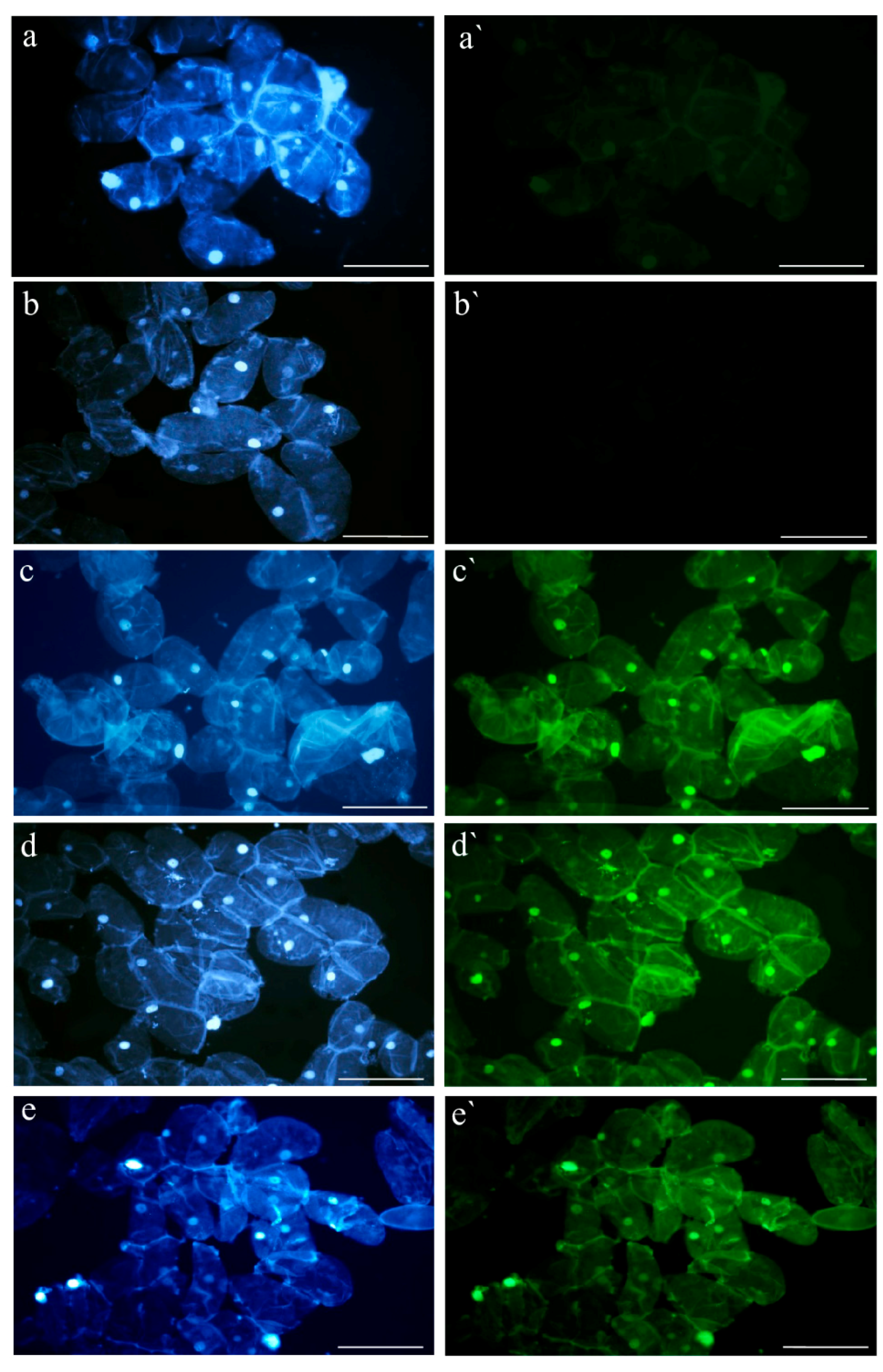
4.4. TUNEL Assay
4.5. Gene Expression Analysis
4.6. Immunocytochemistry
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

References
- Rancour, D.M.; Marita, J.M.; Hatfield, R.D. Cell wall composition throughout development for the model grass Brachypodium distachyon. Front. Plant Sci. 2012, 3, 266. [Google Scholar] [CrossRef] [PubMed]
- Betekhtin, A.; Rojek, M.; Milewska-Hendel, A.; Gawecki, R.; Karcz, J.; Kurczynska, E.; Hasterok, R. Spatial distribution of selected chemical cell wall components in the embryogenic callus of Brachypodium distachyon. PLoS ONE 2016, 11, e0167426. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.I.; Gayomba, S.R.; Yan, J.; Vatamaniuk, O.K. Brachypodium distachyon as a model system for studies of copper transport in cereal crops. Front. Plant Sci. 2014, 5, 236. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.J.; Koehler, A.D.; Rocha, D.I.; Vieira, L.M.; Pinheiro, M.V.; de Matos, E.M.; da Cruz, A.C.; da Silva, T.C.; Tanaka, F.A.; Nogueira, F.T.; et al. Morpho-histological, histochemical, and molecular evidences related to cellular reprogramming during somatic embryogenesis of the model grass Brachypodium distachyon. Protoplasma 2017, 254, 2017–2034. [Google Scholar] [CrossRef] [PubMed]
- Kurczynska, E.U.; Potocka, I.; Dobrowolska, I.; Kulinska-Lukaszek, K.; Sala, K.; Wrobel, J. Cellular markers for somatic embryogenesis. In Embryogenesis; Sato, K.-I., Ed.; InTech: Rijeka, Croatia, 2012; Chapter 14; pp. 307–332. Available online: https://www.intechopen.com/download/pdf/35572 (accessed on 13 June 2018).
- Vanyushin, B.F.; Ashapkin, V.V. DNA methylation in higher plants: Past, present and future. Biochim. Biophys. Acta 2011, 1809, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Matzke, M.A.; Primig, M.; Trnovsky, J.; Matzke, A.J. Reversible methylation and inactivation of marker genes in sequentially transformed tobacco plants. EMBO J. 1989, 8, 643–649. [Google Scholar] [PubMed]
- Saze, H.; Tsugane, K.; Kanno, T.; Nishimura, T. DNA methylation in plants: Relationship to small RNAs and histone modifications, and functions in transposon inactivation. Plant Cell Physiol. 2012, 53, 766–784. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Fukushima, S.; Takatsuji, H. RNA-directed DNA methylation induces transcriptional activation in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 1660–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Kobayashi, H.; Togashi, T.; Mori, Y.; Kikuchi, K.; Kuriyama, K.; Tokuji, Y. Formation of embryogenic cell clumps from carrot epidermal cells is suppressed by 5-azacytidine, a DNA methylation inhibitor. Plant Phys. J. 2005, 162, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Quinga, L.A.; Fraga, H.; Nascimento Vieira, L.; Guerra, M.P. Epigenetics of long-term somatic embryogenesis in Theobroma cacao L.: DNA methylation and recovery of embryogenic potential. PCTOC 2017, 131, 295–305. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Genger, R.K.; Kovac, K.; Peacock, W.J.; Dennis, E.S. DNA methylation and the promotion of flowering by vernalization. Proc. Natl. Acad. Sci. USA 1998, 95, 5824–5829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanyushin, B.F.; Shorning, B.Y.; Seredina, A.V.; Aleksandrushkina, N.I. The effects of phytohormones and 5-azacytidine on apoptosis in etiolated wheat seedlings. Russ. J. Plant Physiol. 2002, 49, 501–506. [Google Scholar] [CrossRef]
- Griffin, P.T.; Niederhuth, C.E.; Schmitz, R.J. A Comparative analysis of 5-azacytidine- and zebularine-induced DNA demethylation. G3 (Bethesda) 2016, 6, 2773–2780. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.B.; Wang, L.; Song, Y.C. Identification of programmed cell death in situ in individual plant cells in vivo using a chromosome preparation technique. J. Exp. Bot. 2002, 53, 651–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.P.; Kim, C.; Landgraf, F.; Apel, K. EXECUTER1- and EXECUTER2-dependent transfer of stress-related signals from the plastid to the nucleus of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 10270–10275. [Google Scholar] [CrossRef] [PubMed]
- Salinas, A.E.; Wong, M.G. Glutathione S-transferases a review. Curr. Med. Chem. 1999, 6, 279–309. [Google Scholar] [PubMed]
- Sharma, R.; Sahoo, A.; Devendran, R.; Jain, M. Over-expression of a rice tau class glutathione s-transferase gene improves tolerance to salinity and oxidative stresses in Arabidopsis. PLoS ONE 2014, 9, e92900. [Google Scholar] [CrossRef] [PubMed]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkins, B.A.; Vasil, I.K. Cellular and Molecular Biology of Plant Seed Development; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA, 1997; 648p. [Google Scholar]
- Nothnagel, E.A.; Bacic, A.; Clarke, A.E. Cell and Developmental Biology of Arabinogalactan-Proteins; Kluwer Academic: New York, NY, USA; London, UK, 2000; 301p. [Google Scholar]
- Nothnagel, E.A. Proteoglycans and related components in plant cells. Int. Rev. Cytol. 1997, 174, 195–291. [Google Scholar] [PubMed]
- Showalter, A.M. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. 2001, 58, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Showalter, A.M. Yariv reagent treatment induces programmed cell death in Arabidopsis cell cultures and implicates arabinogalactan protein involvement. Plant J. 1999, 19, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Qian, J.; Luo, W.; Zhan, Y.; Xin, Y.; Yang, C. Stability of transgenes in long-term micropropagation of plants of transgenic birch (Betula platyphylla). Biotechnol. Lett. 2010, 32, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Yan-hao, X.; Wang, J. DNA-methylation changes induced by salt stress in wheat Triticum aestivum. Afr. J. Biotechnol. 2009, 8, 6201–6207. [Google Scholar]
- Chwialkowska, K.; Nowakowska, U.; Mroziewicz, A.; Szarejko, I.; Kwasniewski, M. Water-deficiency conditions differently modulate the methylome of roots and leaves in barley (Hordeum vulgare L.). J. Exp. Bot. 2016, 67, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Seymour, D.K.; Becker, C. The causes and consequences of DNA methylome variation in plants. Curr. Opin. Plant Biol. 2017, 36, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Solis, M.T.; El-Tantawy, A.A.; Cano, V.; Risueno, M.C.; Testillano, P.S. 5-azacytidine promotes microspore embryogenesis initiation by decreasing global DNA methylation, but prevents subsequent embryo development in rapeseed and barley. Front. Plant Sci. 2015, 6, 472. [Google Scholar] [CrossRef] [PubMed]
- Fraga, H.P.; Vieira, L.N.; Caprestano, C.A.; Steinmacher, D.A.; Micke, G.A.; Spudeit, D.A.; Pescador, R.; Guerra, M.P. 5-Azacytidine combined with 2,4-D improves somatic embryogenesis of Acca sellowiana (O. Berg) Burret by means of changes in global DNA methylation levels. Plant Cell Rep. 2012, 31, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Hailong, S.; Jianmin, C. Effect of 5-azacytidine on zygotic cotyledon explants and somatic embryogenesis of Fraxinus mandshurica. Northeast For. Univ. J. 2011, 8, 25–27. [Google Scholar]
- Shkute, N.; Stivrina, N. 5-Azacytidine decreases fragmentation of nuclear DNA and pigment formation in first leaf cells of barley seedlings. Cell Biol. Int. 2005, 29, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Vanyushin, B.F.; Bakeeva, L.E.; Zamyatnina, V.A.; Aleksandrushkina, N.I. Apoptosis in plants: Specific features of plant apoptotic cells and effect of various factors and agents. Int. Rev. Cytol. 2004, 233, 135–179. [Google Scholar] [PubMed]
- Kiziltepe, T.; Hideshima, T.; Catley, L.; Raje, N.; Yasui, H.; Shiraishi, N.; Okawa, Y.; Ikeda, H.; Vallet, S.; Pozzi, S.; et al. 5-Azacytidine, a DNA methyltransferase inhibitor, induces ATR-mediated DNA double-strand break responses, apoptosis, and synergistic cytotoxicity with doxorubicin and bortezomib against multiple myeloma cells. Mol. Cancer Ther. 2007, 6, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, J. The Effects of 5-Azacytidine on Transformed Hairy Root Cultures of Artemisia Annua. Ph.D. Thesis, Worcester Polytechnic Institute, Worcester, MA, USA, 2006. [Google Scholar]
- Teyssier, C.; Maury, S.; Beaufour, M.; Grondin, C.; Delaunay, A.; Le Mette, C.; Ader, K.; Cadene, M.; Label, P.; Lelu-Walter, M.A. In search of markers for somatic embryo maturation in hybrid larch (Larix × eurolepis): Global DNA methylation and proteomic analyzes. Physiol. Plant. 2014, 150, 271–291. [Google Scholar] [CrossRef] [PubMed]
- Juttermann, R.; Li, E.; Jaenisch, R. Toxicity of 5-aza-2′-deoxycytidine to mammalian cells is mediated primarily by covalent trapping of DNA methyltransferase rather than DNA demethylation. Proc. Natl. Acad. Sci. USA 1994, 91, 11797–11801. [Google Scholar] [CrossRef] [PubMed]
- Burn, J.E.; Bagnall, D.J.; Metzger, J.D.; Dennis, E.S.; Peacock, W.J. DNA methylation, vernalization, and the initiation of flowering. Proc. Natl. Acad. Sci. USA 1993, 90, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Emani, C.; Sunilkumar, G.; Rathore, K.S. Transgene silencing and reactivation in sorghum. Plant Sci. 2002, 162, 181–192. [Google Scholar] [CrossRef]
- Van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklin-Tong, V.E.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological classification of plant cell deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladish, D.K.; Xu, J.; Niki, T. Apoptosis-like programmed cell death occurs in procambium and ground meristem of pea (Pisum sativum) root tips exposed to sudden flooding. Ann. Bot. 2006, 97, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Potten, C.; Wilson, J. Apoptosis: The Life and Death of Cells; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Bakeeva, L.E.; Dzyubinskaya, E.V.; Samuilov, V.D. Programmed cell death in plants: Ultrastructural changes in pea guard cells. Biochemistry 2005, 70, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Gray, J. Programmed Cell Death in Plants; Blackwell Publishing/CRC Press: Oxford, UK; London, UK, 2004. [Google Scholar]
- White, E. Life, death, and the pursuit of apoptosis. Genes Dev. 1996, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Austin, J.R., 2nd; Frost, E.; Vidi, P.A.; Kessler, F.; Staehelin, L.A. Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 2006, 18, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Munne-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef]
- Sam, O.; Ramirez, C.; Coronado, M.J.; Testillano, P.S.; Risueno, M.C. Changes in tomato leaves induced by NaCl stress: Leaf organization and cell ultrastructure. Biol. Plant. 2003, 47, 361–366. [Google Scholar] [CrossRef]
- Britvec, M.; Reichenauer, T.; Soja, G.; Ljubesic, N.; Eid, M.; Pecina, M. Ultrastructure changes in grapevine chloroplasts caused by increased tropospheric ozone concentrations. Biologia 2001, 56, 417–424. [Google Scholar]
- Gadjev, I.; Stone, J.M.; Gechev, T.S. Programmed cell death in plants: New insights into redox regulation and the role of hydrogen peroxide. Int. Rev. Cell Mol. Biol. 2008, 270, 87–144. [Google Scholar] [PubMed]
- Vaux, D.L.; Strasser, A. The molecular biology of apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolska, I.; Kwasniewska, J.; Barlow, P.W.; Kurczynska, E. The fate of surface cell layers of Daucus carota (L.) embryos raised in suspension culture. Plant Biosyst. 2015, 150, 622–630. [Google Scholar] [CrossRef]
- Ezaki, B.; Suzuki, M.; Motoda, H.; Kawamura, M.; Nakashima, S.; Matsumoto, H. Mechanism of gene expression of Arabidopsis glutathione S-transferase, AtGST1, and AtGST11 in response to aluminum stress. Plant Physiol. 2004, 134, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.Y.; Kim, Y.; Kim, C.S.; Guh, J.O.; Kim, K.C.; Cho, B.H. Characterization of glutation S-transferase gene AtGST1 in Arabidopsis thaliana. Plant Cell Rep. 1998, 17, 700–704. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Gelhaye, E.; Rouhier, N.; Navrot, N.; Jacquot, J.P. The plant thioredoxin system. Cell. Mol. Life Sci. 2005, 62, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Wenderoth, I.; Scheibe, R.; von Schaewen, A. Identification of the cysteine residues involved in redox modification of plant plastidic glucose-6-phosphate dehydrogenase. J. Biol. Chem 1997, 272, 26985–26990. [Google Scholar] [CrossRef] [PubMed]
- Minina, E.A.; Filonova, L.H.; Fukada, K.; Savenkov, E.I.; Gogvadze, V.; Clapham, D.; Sanchez-Vera, V.; Suarez, M.F.; Zhivotovsky, B.; Daniel, G.; et al. Autophagy and metacaspase determine the mode of cell death in plants. J. Cell Biol. 2013, 203, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Meskauskiene, R.; Zhang, S.; Lee, K.P.; Lakshmanan Ashok, M.; Blajecka, K.; Herrfurth, C.; Feussner, I.; Apel, K. Chloroplasts of Arabidopsis are the source and a primary target of a plant-specific programmed cell death signaling pathway. Plant Cell 2012, 24, 3026–3039. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kim, Y.; Guo, Y.; Stevenson, B.; Zhu, J.K. The Arabidopsis SOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion. Plant Cell 2003, 15, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Takac, T.; Burbach, C.; Menzel, D.; Samaj, J. Developmental localization and the role of hydroxyproline rich glycoproteins during somatic embryogenesis of banana (Musa spp. AAA). BMC Plant Biol. 2011, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Egertsdotter, U.; von Arnold, S. Importance of arabinogalactan proteins for the development of somatic embryos of Picea abies. Physiol. Plant. 1995, 93, 334–345. [Google Scholar] [CrossRef]
- Chapman, A.; Blervacq, A.S.; Vasseur, J.; Hilbert, J.L. Arabinogalactan-proteins in Cichorium somatic embryogenesis: Effect of beta-glucosyl Yariv reagent and epitope localisation during embryo development. Planta 2000, 211, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Furlan, C.; Salinas-Grenet, H.; Sandoval, O.; Recabarren, C.; Arrano-Salinas, P.; Soto-Alvear, S.; Orellana, A.; Blanco-Herrera, F. The root hair specific SYP123 regulates the localization of cell wall components and contributes to rizhobacterial priming of induced systemic resistance. Front. Plant Sci. 2016, 7, 1081. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, S.; Almeida, J.; Junqueira, V.; Costa, M.L.; Pereira, L.G. Arabinogalactan proteins as molecular markers in Arabidopsis thaliana sexual reproduction. J. Exp. Bot. 2007, 58, 4027–4035. [Google Scholar] [CrossRef] [PubMed]
- Schindler, T.; Bergfeld, R.; Schopfer, P. Arabinogalactan proteins in maize coleoptiles: Developmental relationship to cell death during xylem differentiation but not to extension growth. Plant J. 1995, 7, 25–36. [Google Scholar] [CrossRef] [PubMed]
- García-Angulo, P.; Willats, W.G.T.; Encina, A.E.; Alonso-Simon, A.; Alvarez, J.M.; Acebes, J.L. Immunocytochemical characterization of the cell walls of bean cell suspensions during habituation and dehabituation to dichlobenil. Physiol. Plant. 2006, 127, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, T.; Ariizumi, T.; Kawai-Yamada, M.; Uchimiya, H.; Toriyama, K. Abolition of the tapetum suicide program ruins microsporogenesis. Plant Cell Physiol. 2006, 47, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.C.; Terneus, K.; Hall, Q.; Tan, L.; Wang, Y.; Wegenhart, B.L.; Chen, L.; Lamport, D.T.; Chen, Y.; Kieliszewski, M.J. Self-assembly of the plant cell wall requires an extensin scaffold. Proc. Natl. Acad. Sci. USA 2008, 105, 2226–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steedman, H.F. Polyester wax; a new ribboning embedding medium for histology. Nature 1957, 179, 1345. [Google Scholar] [CrossRef] [PubMed]
- Wolny, E.; Braszewska-Zalewska, A.; Hasterok, R. Spatial distribution of epigenetic modifications in Brachypodium distachyon embryos during seed maturation and germination. PLoS ONE 2014, 9, e101246. [Google Scholar] [CrossRef] [PubMed]
- Kwasniewska, J.; Kus, A.; Swoboda, M.; Braszewska-Zalewska, A. DNA replication after mutagenic treatment in Hordeum vulgare. Mutat. Res. 2016, 812, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Mornkham, T.; Wangsomnuk, P.P.; Fu, Y.B.; Wangsomnuk, P.; Jogloy, S.; Patanothai, A. Extractions of high quality RNA from the seeds of Jerusalem artichoke and other plant species with high levels of starch and lipid. Plants 2013, 2, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Pennell, R.I.; Janniche, L.; Kjellbom, P.; Scofield, G.N.; Peart, J.M.; Roberts, K. Developmental regulation of a plasma membrane arabinogalactan protein epitope in oilseed rape flowers. Plant Cell 1991, 3, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Linstead, P.J.; Peart, J.; Cooper, C.; Roberts, K. Developmentally regulated epitopes of cell surface arabinogalactan proteins and their relation to root tissue pattern formation. Plant J. 1991, 1, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smallwood, M.; Martin, H.; Knox, J.P. An epitope of rice threonine-rich and hydroxyproline-rich glycoprotein is common to cell-wall and hydrophobic plasma-membrane glycoproteins. Planta 1995, 196, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, M.; Beven, A.; Donovan, N.; Neill, S.J.; Peart, J.; Roberts, K.; Knox, J.P. Localization of cell wall proteins in relation to the developmental anatomy of the carrot root apex. Plant J. 1994, 5, 237–246. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Description of the Genes | Primer Sequence (5′–3′) |
|---|---|---|
| AK437296 | ubiquitine | pF–GAGGGTGGACTCCTTTTGGA |
| pR–TCCACACTCCACTTGGTGCT | ||
| Bradi3g01470.1 | aminoacylase activity, catalase activity | pF–TTGTGAAGAGGTTCGCGGATGC |
| pR–TCCCACACGACTTATCACACTGG | ||
| Bradi3g35620.1 | putative glutathione S-transferase | pF–TTTCCATTGCTGAGCACAAGAGC |
| pR–GGGACTTGACCAAATGGATTGCG | ||
| Bradi5g09650.1 | thioredoxin peroxidase activity | pF–GAACCCTTCAGGCCCTGCAATATG |
| pR–AACCTGCTGGGCAAACCTCATC | ||
| Bradi2g18877.1 | hexokinase | pF–AATGACACGGTAGGCGAGGATG |
| pR–GACTTTCATATCGAGACCCTGACG | ||
| Bradi2g18290.1 | single-stranded DNA specific 5′–3′ exodeoxyribonuclease activity | pF–AGGCACCTTGTGAAGCAGAAGC |
| pR–TCTGATGCGACAGCATACACCTTG | ||
| Bradi1g75270.1 | pheophorbide oxygenase (PAO, ACD1) | pF–ACCGTCCTTTCAAAGCGTGAGATG |
| pR–CGCTCCTTTGCAAGACGAACAC | ||
| Bradi1g60762.1 | metacaspase involved in regulation of apoptosis | pF–ACTGCATCCTCATCCTCACAGAG |
| pR–AGCCAGCAGATTCTCCTTCGTC | ||
| Bradi1g60756.1 | metacaspase involved in regulation of apoptosis | pF–ACTGCATCCTCACCCTTACACC |
| pR–AGAAGTGGAACACCAGGGAGTC | ||
| Bradi1g05570.1 | BAX inhibitor related | pF–ACGCCATCGTCCTGATGTTGTTC |
| pR–TGAGGAAGGCCGAGAAGATGAGC | ||
| Bradi1g02697.1 | XP-G/RAD2 DNA repair endonuclease family | pF–AGGGTTTGACGAAGCTGCTG |
| pR–TCCTTTCCTTCCTACCACAACCAG | ||
| Bradi3g01470.2 | aminoacylase activity, catalase activity | pF–AGGTGATGGACCCAGATGAG |
| pR–GAAGTTGTCCACGTTTCGGT | ||
| Bradi3g29270.1 | protein executer 1 | pF–GTTGGTGGCAACAGGAAACT |
| pR–GAATTCGGCTGAAGTGGGTA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betekhtin, A.; Milewska-Hendel, A.; Chajec, L.; Rojek, M.; Nowak, K.; Kwasniewska, J.; Wolny, E.; Kurczynska, E.; Hasterok, R. 5-Azacitidine Induces Cell Death in a Tissue Culture of Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 1806. https://doi.org/10.3390/ijms19061806
Betekhtin A, Milewska-Hendel A, Chajec L, Rojek M, Nowak K, Kwasniewska J, Wolny E, Kurczynska E, Hasterok R. 5-Azacitidine Induces Cell Death in a Tissue Culture of Brachypodium distachyon. International Journal of Molecular Sciences. 2018; 19(6):1806. https://doi.org/10.3390/ijms19061806
Chicago/Turabian StyleBetekhtin, Alexander, Anna Milewska-Hendel, Lukasz Chajec, Magdalena Rojek, Katarzyna Nowak, Jolanta Kwasniewska, Elzbieta Wolny, Ewa Kurczynska, and Robert Hasterok. 2018. "5-Azacitidine Induces Cell Death in a Tissue Culture of Brachypodium distachyon" International Journal of Molecular Sciences 19, no. 6: 1806. https://doi.org/10.3390/ijms19061806
APA StyleBetekhtin, A., Milewska-Hendel, A., Chajec, L., Rojek, M., Nowak, K., Kwasniewska, J., Wolny, E., Kurczynska, E., & Hasterok, R. (2018). 5-Azacitidine Induces Cell Death in a Tissue Culture of Brachypodium distachyon. International Journal of Molecular Sciences, 19(6), 1806. https://doi.org/10.3390/ijms19061806