Boldine Improves Kidney Damage in the Goldblatt 2K1C Model Avoiding the Increase in TGF-β



Abstract
:1. Introduction
2. Results
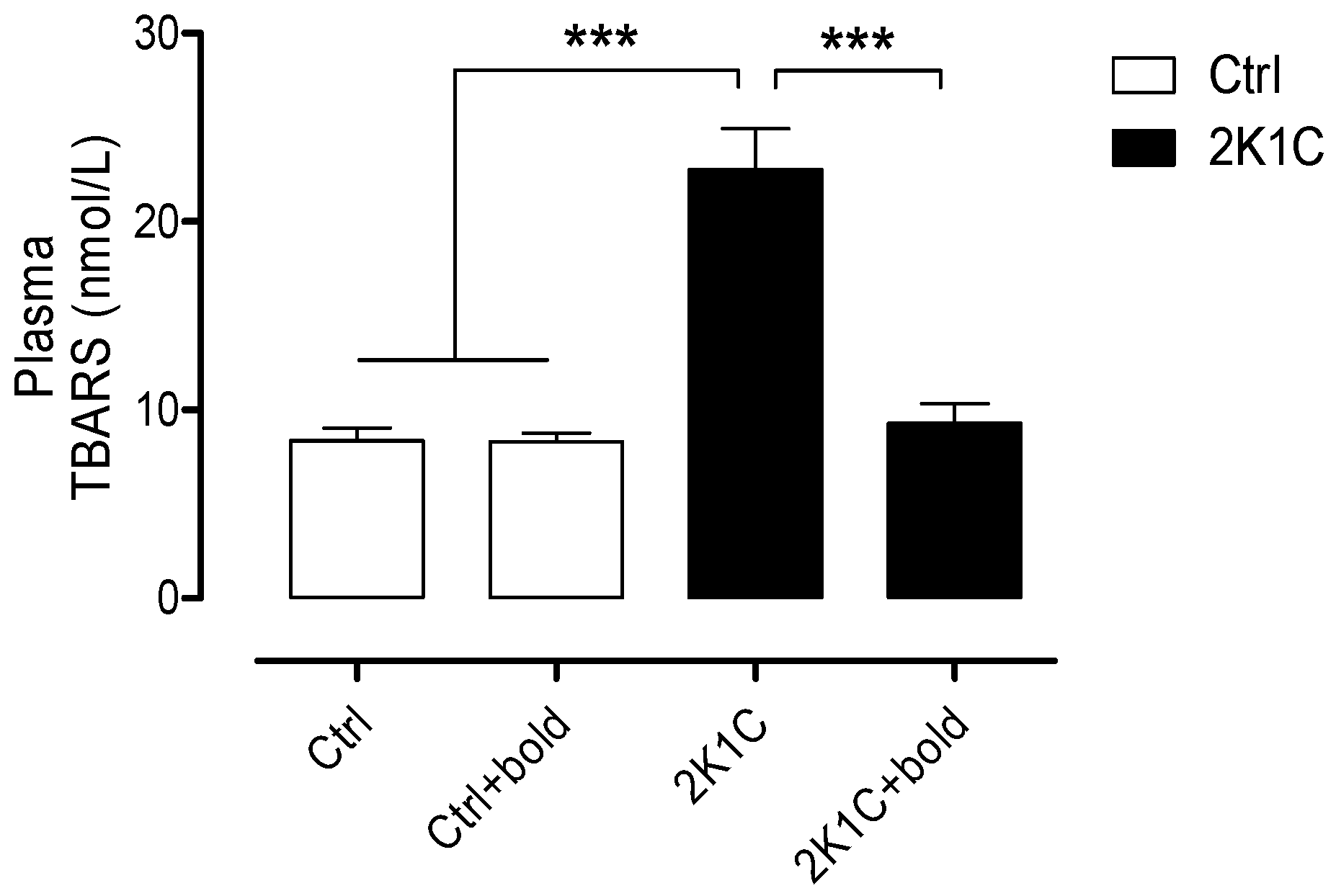
2.1. Boldine Improves Kidney Function and Decreases OS without Decreasing Systolic Blood Pressure (SBP)
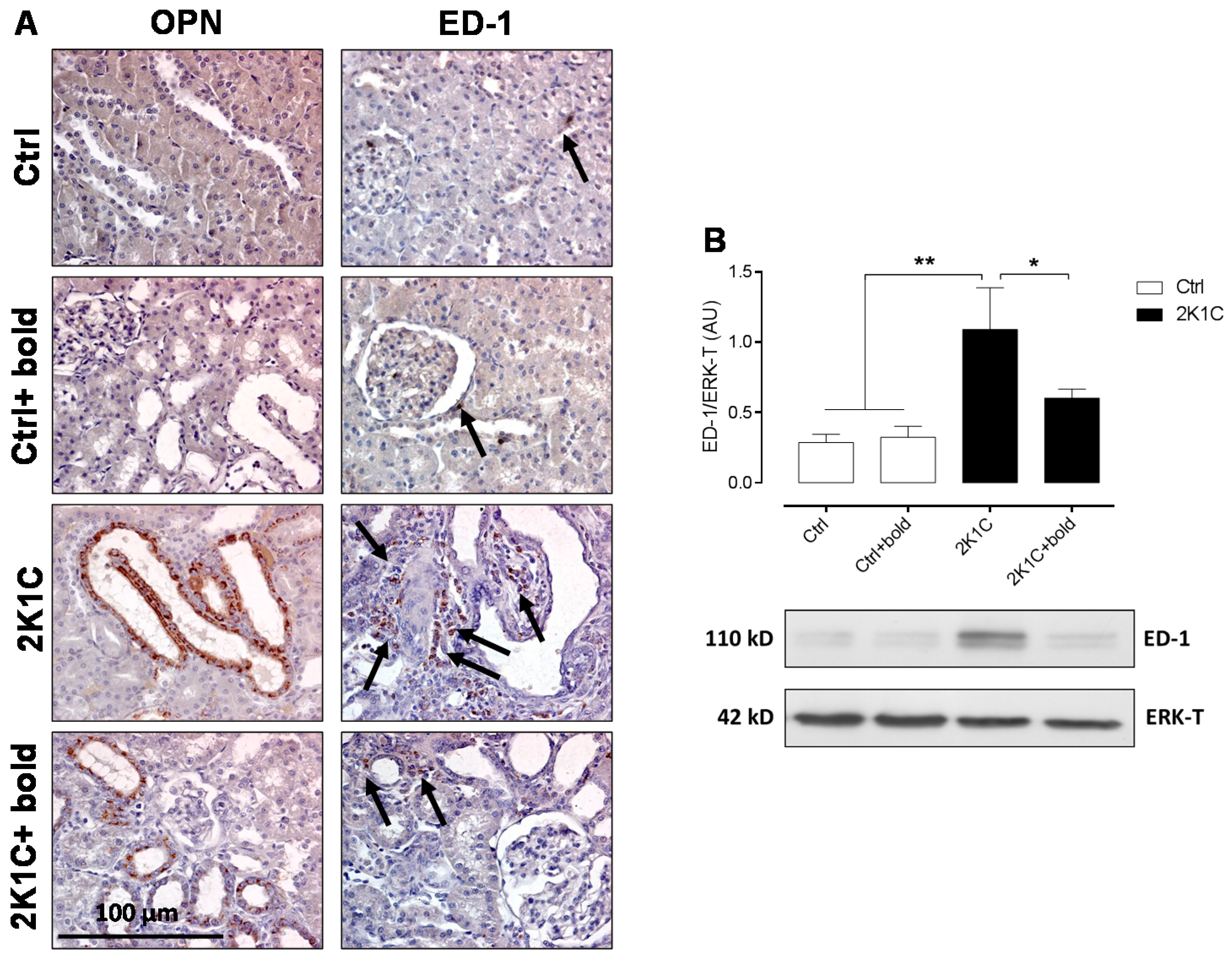
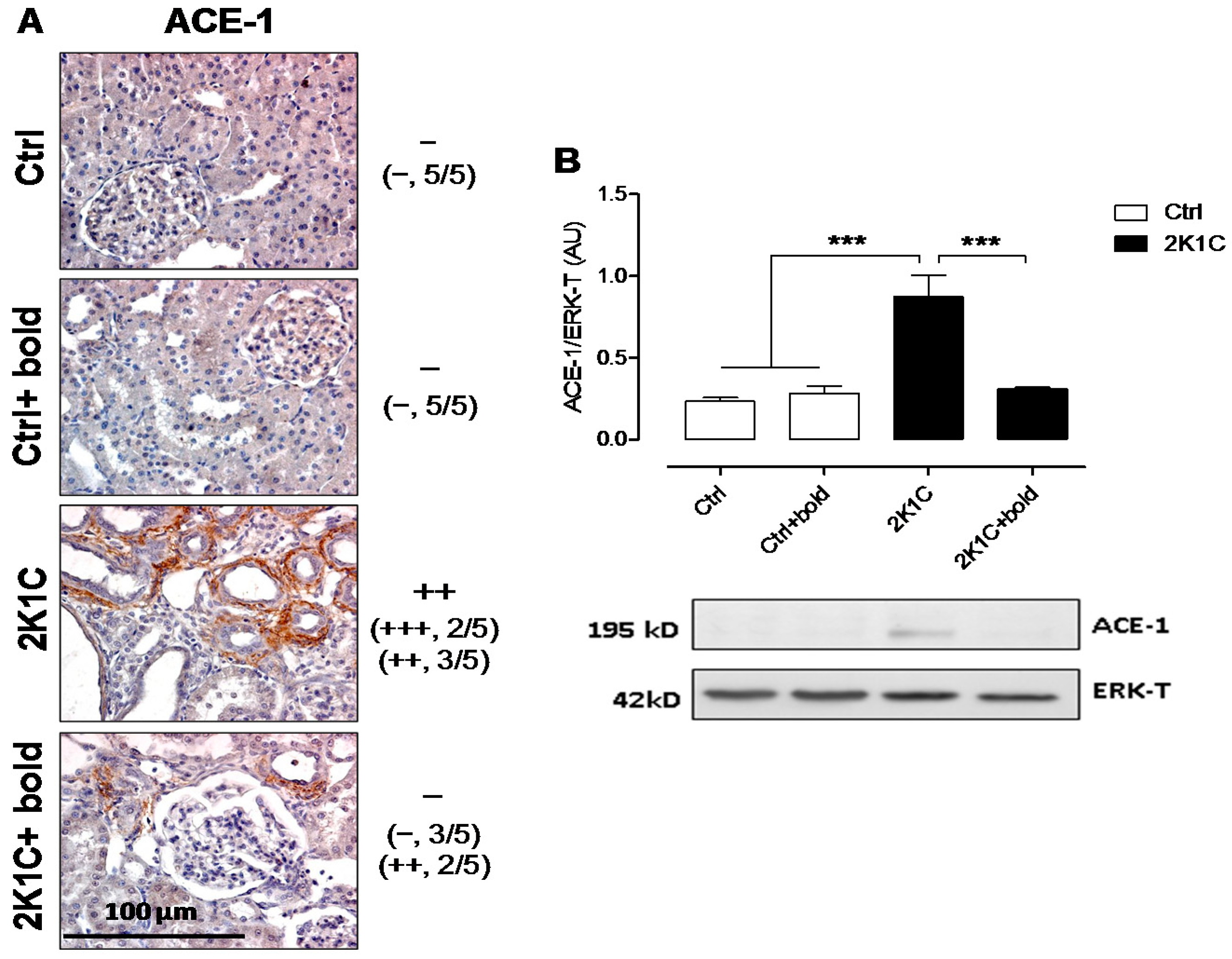
2.2. Boldine Reduces Renal Tissue Damage and Inflammation in Hypertensive 2K1C Rats
2.3. TGF-β, Which Has Been Linked to Kidney Damage, Was Reduced by Boldine Treatment
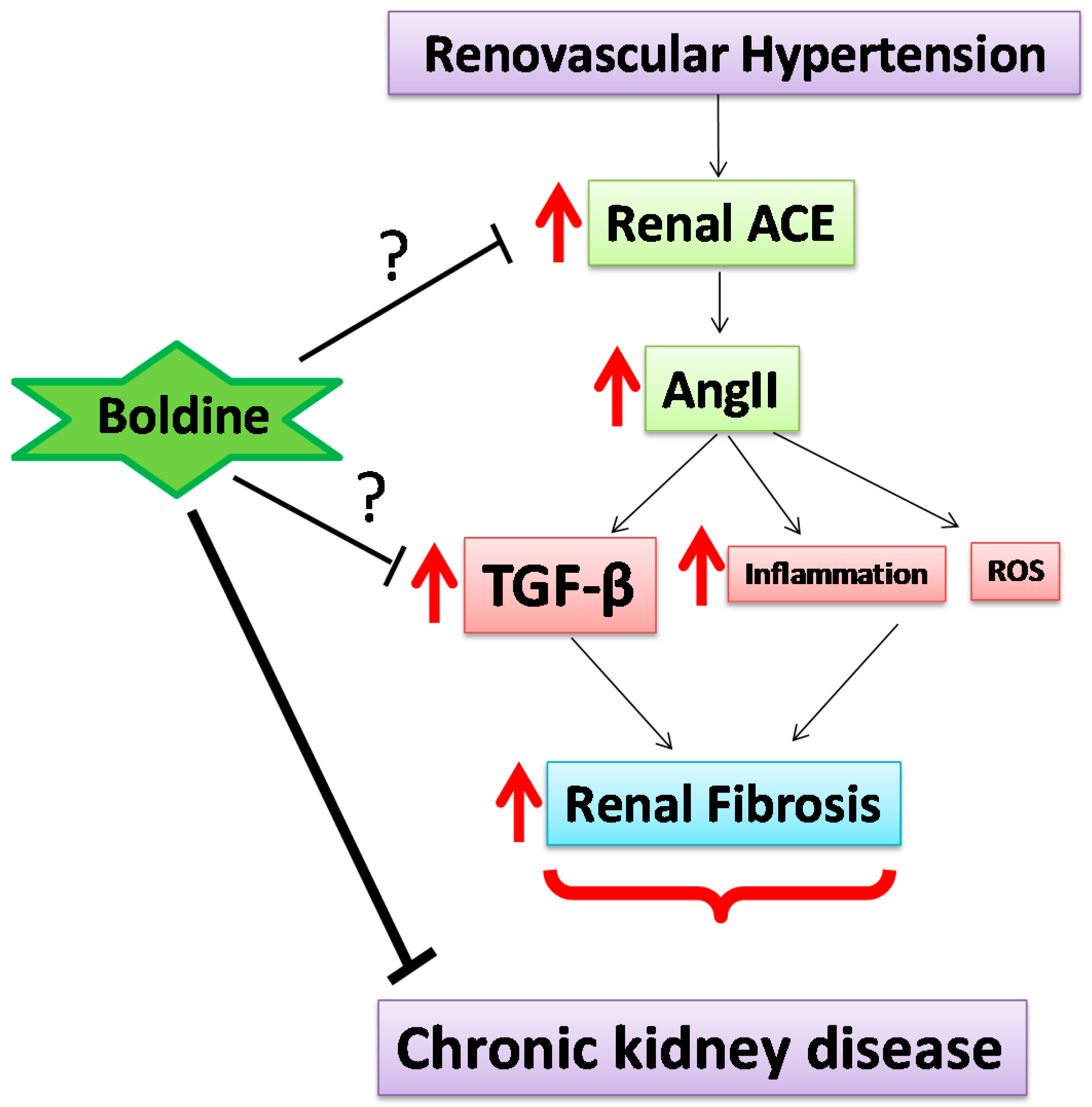
3. Discussion
4. Conclusions
5. Methods
5.1. Antibodies and Chemicals
5.2. Animals and Experimental Protocol
5.3. Blood Pressure Measurements
5.4. Renal Function Measurements
5.5. Thiobarbituric Acid Reactive Substances (TBARS) Measurement
5.6. Histological Damage Assessment
5.7. Tissue Processing and Immunohistochemical Analysis
5.8. Western Blot
5.9. Enzyme-Linked Immunosorbent Assay
5.10. Statistical Analysis.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Renin-Angiotensin System | RAS |
| Chronic kidney disease | CDK |
| Glomerular filtration rate | GFR |
| Goldblatt two-kidney one clip | 2K1C |
| Renovascular hypertension | RVH |
| transforming growth factor-β | TGF-β |
| Extracellular matrix | ECM |
| Alpha-smooth muscle actin | α-SMA |
| Fibronectin | FN |
| Collagen type III | Col III |
| infiltration of macrophages | ED-1 |
| Osteopontin | OPN |
| Reactive oxygen species | ROS |
| Oxidative stress | OS |
| Angiotensin converter enzyme | ACE |
| Systolic blood pressure | SBP |
| Enzyme-Linked Immunosorbent Assay | ELISA |
References
- Ozawa, Y.; Kobori, H.; Suzaki, Y.; Navar, L.G. Sustained renal interstitial macrophage infiltration following chronic angiotensin II infusions. Am. J. Physiol. Renal Physiol. 2007, 292, F330–F339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.J.; Alpers, C.E.; Yoshimura, A.; Lombardi, D.; Pritzl, P.; Floege, J.; Schwartz, S.M. Renal injury from angiotensin II-mediated hypertension. Hypertension (Dallas Tex. 1979) 1992, 19, 464–474. [Google Scholar] [CrossRef]
- Cheng, J.; Zhou, W.; Warner, G.M.; Knudsen, B.E.; Garovic, V.D.; Gray, C.E.; Lerman, L.O.; Platt, J.L.; Romero, J.C.; Textor, S.C.; et al. Temporal analysis of signaling pathways activated in a murine model of two-kidney, one-clip hypertension. Am. J. Physiol. Renal Physiol. 2009, 297, F1055–F1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Saura, M.F.; Galisteo, M.; Villar, I.C.; Bermejo, A.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of chronic quercetin treatment in experimental renovascular hypertension. Mol. Cell. Biochem. 2005, 270, 147–155. [Google Scholar] [CrossRef] [PubMed]
- López-Novoa, J.M.; Martínez-Salgado, C.; Rodríguez-Peña, A.B.; López-Hernández, F.J. Common pathophysiological mechanisms of chronic kidney disease: Therapeutic perspectives. Pharmacol. Ther. 2010, 128, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Coresh, J.; Byrd-Holt, D.; Astor, B.C.; Briggs, J.P.; Eggers, P.W.; Lacher, D.A.; Hostetter, T.H. Chronic Kidney Disease Awareness, Prevalence, and Trends among U.S. Adults, 1999 to 2000. J. Am. Soc. Nephrol. 2004, 16, 180–188. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, D.; Hillege, H.L.; de Jong, P.E. The kidney, a cardiovascular risk marker, and a new target for therapy. Kidney Int. Suppl. 2005, 68, S25–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldblatt, H.; Kahn, J.R.; Hanzal, R.F. Studies on experimental hypertension: IX. The effect on blood pressure of constriction of the abdominal aorta above and below the site of origin of both main renal arteries. J. Exp. Med. 1939, 69, 649–674. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; Fox, J.; Mitchell, K.D.; Navar, L.G. Angiotensin and angiotensin converting enzyme tissue levels in two-kidney, one clip hypertensive rats. Hypertension (Dallas Tex. 1979) 1992, 20, 763–767. [Google Scholar] [CrossRef]
- Border, W.A.; Noble, N.A. Interactions of transforming growth factor-beta and angiotensin II in renal fibrosis. Hypertension (Dallas Tex. 1979) 1998, 31, 181–188. [Google Scholar] [CrossRef]
- Yang, F.; Chung, A.C.K.; Huang, X.R.; Lan, H.Y. Angiotensin II induces connective tissue growth factor and collagen I expression via transforming growth factor-beta-dependent and -independent Smad pathways: The role of Smad3. Hypertension (Dallas Tex. 1979) 2009, 54, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Mezzano, S.A.; Ruiz-Ortega, M.; Egido, J. Angiotensin II and renal fibrosis. Hypertension (Dallas Tex. 1979) 2001, 38, 635–638. [Google Scholar] [CrossRef]
- Wolf, G.; Schneider, A.; Wenzel, U.; Helmchen, U.; Stahl, R.A. Regulation of glomerular TGF-beta expression in the contralateral kidney of two-kidney, one-clip hypertensive rats. J. Am. Soc. Nephrol. 1998, 9, 763–772. [Google Scholar] [PubMed]
- Mezzano, S.A.; Aros, C.A.; Droguett, A.; Burgos, M.E.; Ardiles, L.G.; Flores, C.A.; Carpio, D.; Vío, C.P.; Ruiz-Ortega, M.; Egido, J. Renal angiotensin II up-regulation and myofibroblast activation in human membranous nephropathy. Kidney Int. Suppl. 2003, 64, S39–S45. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, S.S.; Chen, Y.; Ahokas, R.A.; Sun, Y. Kidney fibrosis in hypertensive rats: Role of oxidative stress. Am. J. Nephrol. 2008, 28, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Clermont, G.; Lecour, S.; Lahet, J.; Siohan, P.; Vergely, C.; Chevet, D.; Rifle, G.; Rochette, L. Alteration in plasma antioxidant capacities in chronic renal failure and hemodialysis patients: A possible explanation for the increased cardiovascular risk in these patients. Cardiovasc. Res. 2000, 47, 618–623. [Google Scholar] [CrossRef]
- Forni, C.; Braglia, R.; Mulinacci, N.; Urbani, A.; Ronci, M.; Gismondi, A.; Tabolacci, C.; Provenzano, B.; Lentini, A.; Beninati, S. Antineoplastic activity of strawberry (Fragaria × ananassa Duch.) crude extracts on B16-F10 melanoma cells. Mol. BioSyst. 2014, 10, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Impei, S.; Gismondi, A.; Canuti, L.; Canini, A. Metabolic and biological profile of autochthonous Vitis vinifera L. ecotypes. Food Funct. 2015, 6, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.; Carrasco-Pozo, C.; Speisky, H. Boldine and its antioxidant or health-promoting properties. Chem. Biol. Interact. 2006, 159, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Speisky, H.; Cassels, B.K. Boldo and boldine: An emerging case of natural drug development. Pharmacol. Res. 1994, 29, 1–12. [Google Scholar] [CrossRef]
- Methven, S.; MacGregor, M.S.; Traynor, J.P.; O’Reilly, D.S.J.; Deighan, C.J. Assessing proteinuria in chronic kidney disease: Protein-creatinine ratio versus albumin-creatinine ratio. Nephrol. Dial. Transplant. 2010, 25, 2991–2996. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, C.W.; Park, J.H.; Lim, S.W.; Sun, B.K.; Jung, J.Y.; Kim, S.B.; Kim, Y.S.; Kim, J.; Bang, B.K. Pravastatin treatment attenuates interstitial inflammation and fibrosis in a rat model of chronic cyclosporine-induced nephropathy. Am. J. Physiol. Renal Physiol. 2004, 286, F46–F57. [Google Scholar] [CrossRef] [PubMed]
- Correa-Rotter, R.; Cusumano, A.M. Present, Prevention, and Management of Chronic Kidney Disease in Latin America. Blood Purif. 2008, 26, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Vilayur, E.; Harris, D.C.H. Emerging therapies for chronic kidney disease: What is their role? Nat. Rev. Nephrol. 2009, 5, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.K.; Griendling, K.K. Angiotensin II cell signaling: Physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 2007, 292, C82–C97. [Google Scholar] [CrossRef] [PubMed]
- Price, C.P.; Newall, R.G.; Boyd, J.C. Use of protein:creatinine ratio measurements on random urine samples for prediction of significant proteinuria: A systematic review. Clin. Chem. 2005, 51, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Wilmer, W.A.; Rovin, B.H.; Hebert, C.J.; Rao, S.V.; Kumor, K.; Hebert, L.A. Management of glomerular proteinuria: A commentary. J. Am. Soc. Nephrol. 2003, 14, 3217–3232. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.J.; Onstead-Haas, L.; Lee, T.; Torfah, M.; Mooradian, A.D. Angiotensin II receptor one (AT1) mediates dextrose induced endoplasmic reticulum stress and superoxide production in human coronary artery endothelial cells. Int. J. Cardiol. 2016, 220, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Wang, W.; Wang, Y.; Du, T.; Shen, W.; Tang, H.; Wang, Y.; Yin, H. Bletilla striata polysaccharide inhibits angiotensin II-induced ROS and inflammation via NOX4 and TLR2 pathways. Int. J. Biol. Macromol. 2016, 89, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Kao, M.P.C.; Ang, D.S.C.; Pall, A.; Struthers, A.D. Oxidative stress in renal dysfunction: Mechanisms, clinical sequelae and therapeutic options. J. Hum. Hypertens. 2010, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, E.; Rodriguez-Vita, J.; Cartier, C.; Rupérez, M.; Esteban, V.; Carvajal, G.; Rodrígues-Díez, R.; Plaza, J.J.; Egido, J.; Ruiz-Ortega, M. Inhibitory effect of interleukin-1beta on angiotensin II-induced connective tissue growth factor and type IV collagen production in cultured mesangial cells. Am. J. Physiol. Renal Physiol. 2008, 294, F149–F160. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Bahrami, L.; Castillo, A.; Majid, D.S.A. TNF-α type 2 receptor mediates renal inflammatory response to chronic angiotensin II administration with high salt intake in mice. Am. J. Physiol. Renal Physiol. 2013, 304, F991–F999. [Google Scholar] [CrossRef] [PubMed]
- Zagorova, M.; Prasnicka, A.; Kadova, Z.; Dolezelova, E.; Kazdova, L.; Cermanova, J.; Rozkydalova, L.; Hroch, M.; Mokry, J.; Micuda, S. Boldine attenuates cholestasis associated with nonalcoholic fatty liver disease in hereditary hypertriglyceridemic rats fed by high-sucrose diet. Physiol. Res. 2015, 64 (Suppl. 4), S467–S476. [Google Scholar] [PubMed]
- Lanhers, M.C.; Joyeux, M.; Soulimani, R.; Fleurentin, J.; Sayag, M.; Mortier, F.; Younos, C.; Pelt, J.M. Hepatoprotective and anti-inflammatory effects of a traditional medicinal plant of Chile, Peumus boldus. Planta Med. 1991, 57, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Lagos, P.; Rivera, P.; Zamorano-Ponce, E. Effect of boldo (Peumus boldus Molina) infusion on lipoperoxidation induced by cisplatin in mice liver. Phytother. Res. 2009, 23, 1024–1027. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Rivera, J.; Ortiz, A.; Egido, J. Antioxidants in kidney diseases: The impact of bardoxolone methyl. Int. J. Nephrol. 2012, 2012, 321714. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yokozawa, T.; Cho, E.J.; Oowada, S.; Kim, M. Protective role of gamma-aminobutyric acid against chronic renal failure in rats. J. Pharm. Pharmacol. 2006, 58, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Toklu, H.Z.; Sehirli, O.; Erşahin, M.; Süleymanoğlu, S.; Yiğiner, O.; Emekli-Alturfan, E.; Yarat, A.; Yeğen, B.Ç.; Sener, G. Resveratrol improves cardiovascular function and reduces oxidative organ damage in the renal, cardiovascular and cerebral tissues of two-kidney, one-clip hypertensive rats. J. Pharm. Pharmacol. 2010, 62, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.-S.; Machha, A.; Achike, F.I.; Murugan, D.; Mustafa, M.R. The aporphine alkaloid boldine improves endothelial function in spontaneously hypertensive rats. Exp. Biol. Med. (Maywood) 2012, 237, 93–98. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Lee, L.; Yang, J.; Wang, X. Preventive effects and mechanisms of rhein on renal interstitial fibrosis in obstructive nephropathy. Biol. Pharm. Bull. 2011, 34, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Giachelli, C.M.; Pichler, R.; Lombardi, D.; Denhardt, D.T.; Alpers, C.E.; Schwartz, S.M.; Johnson, R.J. Osteopontin expression in angiotensin II-induced tubulointerstitial nephritis. Kidney Int. 1994, 45, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, L.F. Effects of HMG-CoA reductase inhibitors (statins) on progression of kidney disease. Kidney Int. 2008, 74, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Satoh, T.; Hibi, D.; Ohno, Y.; Kohda, Y.; Miura, K.; Gemba, M. The effect of antioxidant on development of fibrosis by cisplatin in rats. J. Pharmacol. Sci. 2009, 111, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Onozato, M.L.; Tojo, A.; Kobayashi, N.; Goto, A.; Matsuoka, H.; Fujita, T. Dual blockade of aldosterone and angiotensin II additively suppresses TGF-beta and NADPH oxidase in the hypertensive kidney. Nephrol. Dial. Transplant. 2007, 22, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.-X.; Zhang, A.-H.; Huang, S.-M.; Pan, X.-Q.; Chen, R.-H. SP600125, an inhibitor of c-Jun NH2-terminal kinase, blocks expression of angiotensin II-induced monocyte chemoattractant protein-1 in human mesangial cells. World J. Pediatr. 2010, 6, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, S.; Cespedes, C.; Gonzalez, A.; Vio, C.P. bFGF induces an earlier expression of nephrogenic proteins after ischemic acute renal failure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1677–R1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, S.P.; Giacaman, A.; Vío, C.P. Pregnant rats with 5/6 nephrectomy have normal volume expansion despite lower renin and kallikrein. Hypertension (Dallas Tex. 1979) 2003, 42, 744–748. [Google Scholar] [CrossRef] [PubMed]
- López-De León, A.; Rojkind, M. A simple micromethod for collagen and total protein determination in formalin-fixed paraffin-embedded sections. J. Histochem. Cytochem. 1985, 33, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Vio, C.P.; An, S.J.; Céspedes, C.; McGiff, J.C.; Ferreri, N.R. Induction of cyclooxygenase-2 in thick ascending limb cells by adrenalectomy. J. Am. Soc. Nephrol. 2001, 12, 649–658. [Google Scholar] [PubMed]
- Vio, C.P.; Quiroz-Munoz, M.; Cuevas, C.A.; Cespedes, C.; Ferreri, N.R. Prostaglandin E2 EP3 receptor regulates cyclooxygenase-2 expression in the kidney. Am. J. Physiol. Cell Physiol. 2012, 303, F449–F457. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Weight (gr) | Proteinuria (mg/day) | Urine Creatinine (mg/day) | Plasma Creatinine (mg/mL) | Creatinine clearence (mL/min) | FE Na+ (%) | FE K+ (%) |
|---|---|---|---|---|---|---|---|
| Ctrl (n = 5) | 410 ± 10.5 | 11.6 ± 2.4 *** | 6.4 ± 1.8 | 0.3 ± 0.0 *** | 0.4 ± 0.1 * | 0.1 ± 0.0 ** | 29.6 ± 6.2 ** |
| Ctrl + bold (n = 5) | 412 ± 20.3 | 15.9 ± 6.0 *** | 6.6 ± 1.0 | 0.2 ± 0.0 *** | 0.4 ± 0.1 * | 0.1 ± 0.0 ** | 21.6 ± 1.1 ** |
| 2K1C (n = 5) | 383 ± 8.7 | 132.2 ± 11.7 | 4.3 ± 0.4 | 0.8 ± 0.1 | 0.1 ± 0.0 | 0.6 ± 0.1 | 101.6 ± 23.1 |
| 2K1C + bold (n = 5) | 390 ± 13.4 | 26.4 ± 5.8 *** | 4.4 ± 0.3 | 0.3 ± 0.1 *** | 0.3 ± 0.1 | 0.2 ± 0.1 * | 30.6 ± 6.0 ** |
| Score | |||
|---|---|---|---|
| Groups | Picric-Sirius | Col III | α-SMA |
| Ctrl (n = 5) | − (−, 5/5) | − (−, 4/5; +, 1/5) | − (−, 5/5) |
| Ctrl + bold (n = 5) | − (−, 4/5; +, 1/5) | − (−, 4/5; +, 1/5) | − (−, 5/5) |
| 2K1C (n = 5) | +++ (+++, 4/5; ++, 1/5) | ++ (+++, 2/5; ++, 2/5; +, 1/5) | + (+, 4/5; −, 1/35) |
| 2K1C + bold (n = 5) | + (+, 5/5) | − (−, 3/5; +, 2/5) | − (−, 5/5) |
| Score | ||
|---|---|---|
| Groups | OPN | ED-1 |
| Ctrl (n = 5) | − (−, 5/5) | − (−, 5/5) |
| Ctrl + bold (n = 5) | − (−, 3/5; +, 2/5) | − (−, 4/5; +, 1/5) |
| 2K1C (n = 5) | +++ (+++, 3/5; ++, 2/5) | +++ (+++, 3/5; ++, 2/5) |
| 2K1C + bold (n = 5) | + (+, 4/5; ++, 1/5) | + (−, 2/5; +, 3/5) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, G.I.; Velarde, V. Boldine Improves Kidney Damage in the Goldblatt 2K1C Model Avoiding the Increase in TGF-β. Int. J. Mol. Sci. 2018, 19, 1864. https://doi.org/10.3390/ijms19071864
Gómez GI, Velarde V. Boldine Improves Kidney Damage in the Goldblatt 2K1C Model Avoiding the Increase in TGF-β. International Journal of Molecular Sciences. 2018; 19(7):1864. https://doi.org/10.3390/ijms19071864
Chicago/Turabian StyleGómez, Gonzalo I., and Victoria Velarde. 2018. "Boldine Improves Kidney Damage in the Goldblatt 2K1C Model Avoiding the Increase in TGF-β" International Journal of Molecular Sciences 19, no. 7: 1864. https://doi.org/10.3390/ijms19071864
APA StyleGómez, G. I., & Velarde, V. (2018). Boldine Improves Kidney Damage in the Goldblatt 2K1C Model Avoiding the Increase in TGF-β. International Journal of Molecular Sciences, 19(7), 1864. https://doi.org/10.3390/ijms19071864