Serum miRNAs Predicting Sustained HBs Antigen Reduction 48 Weeks after Pegylated Interferon Therapy in HBe Antigen-Negative Patients

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Study Cohort at Baseline
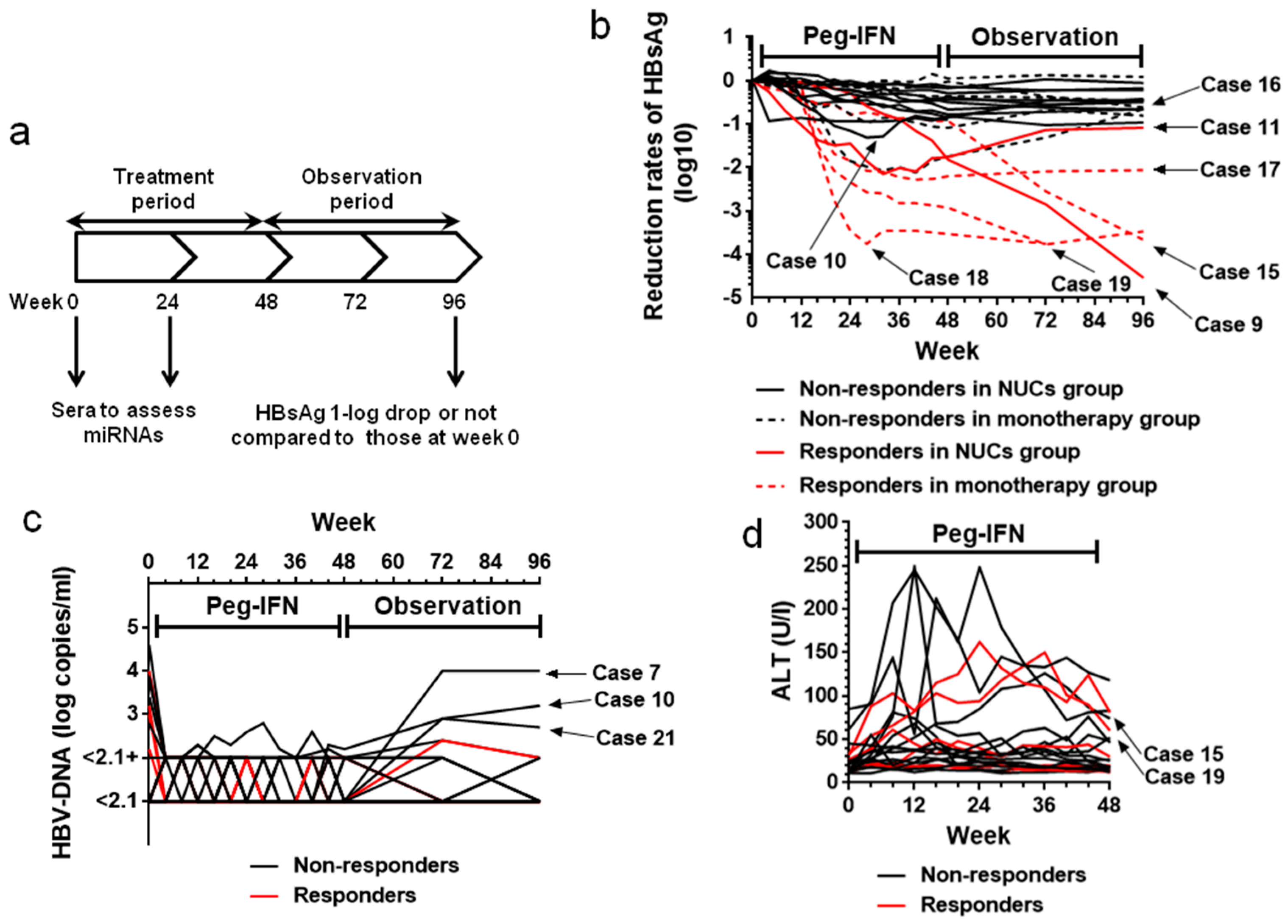
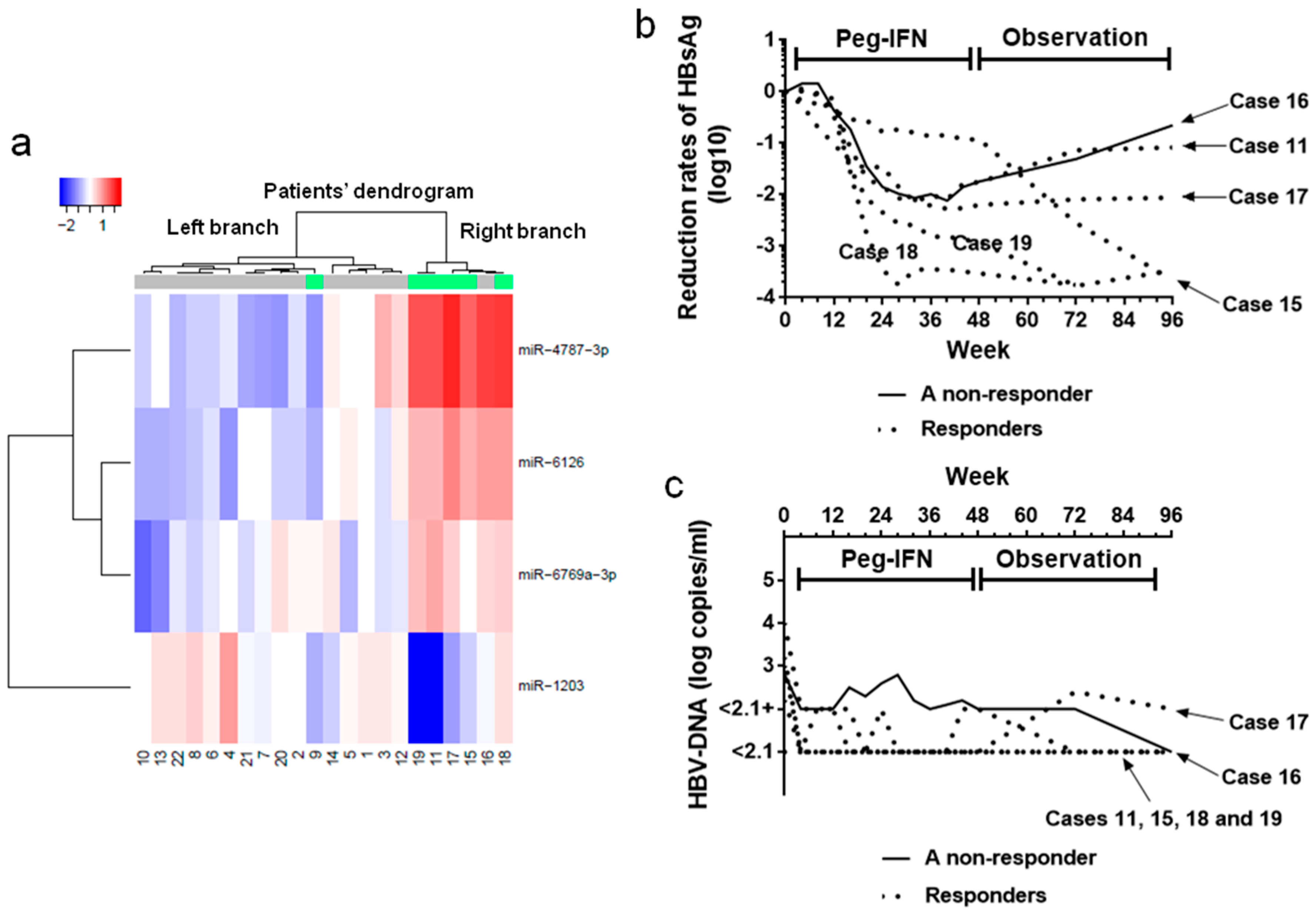
2.2. HBsAg Levels during the Treatment (Weeks 0–48) and Observation (Weeks 49–96) Periods
2.3. HBV-DNA Levels During Treatment (Weeks 0–48) and Observation (Weeks 49–96) Periods
2.4. Transient Increase in ALT Levels
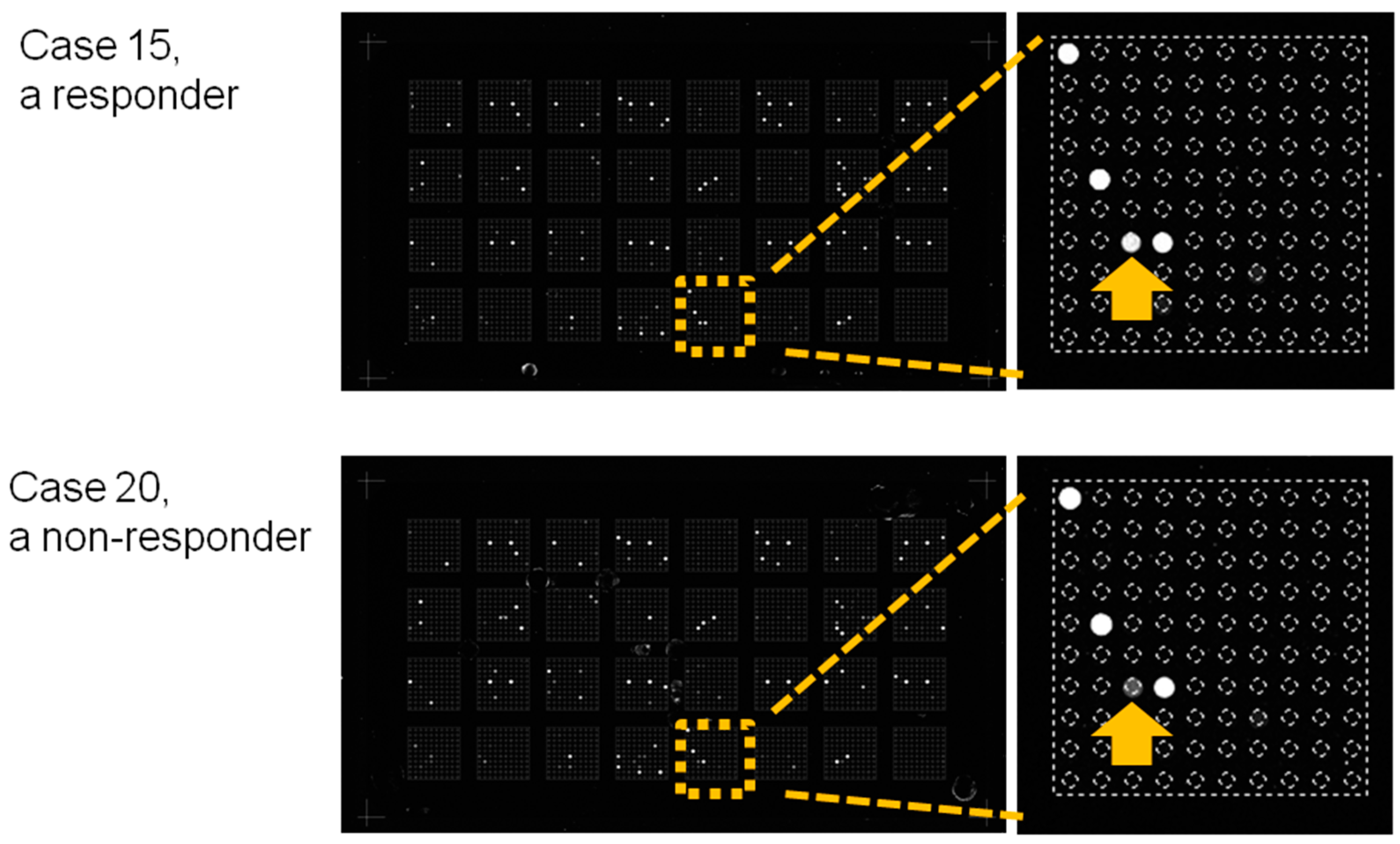
2.5. miRNA Expression Analysis
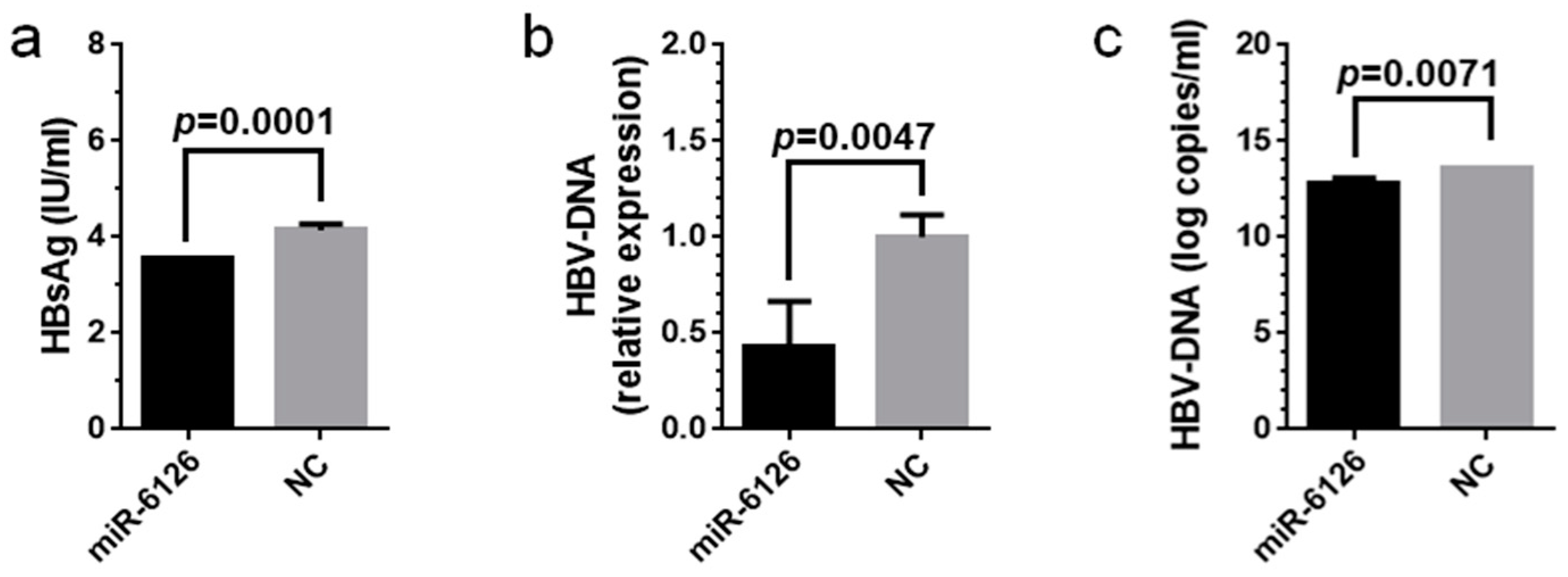
2.6. Virological Effects of miR-6126 against HBV
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Patients
4.3. Study Design
4.4. Efficacy of Treatment
4.5. miRNA Microarray Analysis
4.6. Real-Time qPCR for Serum miRNAs
4.7. Cell Line and Culture
4.8. HBV Infection
4.9. HBV Replication Analysis
4.10. Transfection of Cells with miRNA Mimics
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
| HBV | Hepatitis B virus |
| HBsAg | HBs antigen |
| HBeAg | HBe antigen |
| Peg-IFN | Pegylated interferon |
| NUCs | Nucleos(t)ide analogues |
| ETV | Entecavir |
| LMD | Lamivudine |
| ADV | Adefovir |
| miRNA | MicroRNA |
| FDR | False discovery rate |
| NTCP | Sodium taurocholate cotransporting polypeptide |
| NC | Negative control |
| Fib-4 index | Fibrosis-4 index |
References
- Lok, A.S.F.; McMahon, B.J. Chronic hepatitis B. Hepatology 2007, 45, 507–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, T.C.; Liu, C.J.; Yang, H.C.; Su, T.H.; Wang, C.C.; Chen, C.L.; Hsu, C.A.; Fang-Tzu, K.S.; Liu, C.H.; Chen, P.J.; et al. Serum hepatitis B surface antigen levels help predict disease progression in patients with low hepatitis B virus loads. Hepatology 2013, 57, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.M.; Liaw, Y.F. HBsAg seroclearance in asymptomatic carriers of high endemic areas: Appreciably high rates during a long-term follow-up. Hepatology 2007, 45, 1187–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, A.; Suzuki, F.; Kawamura, Y.; Sezaki, H.; Hosaka, T.; Akuta, N.; Kobayashi, M.; Suzuki, Y.; Saitou, S.; Arase, Y.; et al. Long-term continuous entecavir therapy in nucleos(t)ide-naïve chronic hepatitis B patients. J. Hepatol. 2012, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Marcellin, P.; Zoulim, F.; Hézode, C.; Causse, X.; Roche, B.; Truchi, R.; Pauwels, A.; Ouzan, D.; Dumortier, J.; Pageaux, G.P.; et al. Effectiveness and safety of tenofovir disoproxil fumarate in chronic hepatitis B: A 3-year, prospective, real-world study in France. Dig. Dis. Sci. 2016, 61, 3072–3083. [Google Scholar] [CrossRef] [PubMed]
- Lampertico, P.; Maini, M.; Papatheodoridis, G. Optimal management of hepatitis B virus infection—EASL special conference. J. Hepatol. 2015, 63, 1238–1253. [Google Scholar] [CrossRef] [PubMed]
- Terrault, N.A.; Bzowej, N.H.; Chang, K.M.; Hwang, J.P.; Jonas, M.M.; Murad, M.H. AASLD guidelines for treatment of chronic hepatitis B. Hepatology 2016, 63, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Kumar, M.; Lau, G.K.; Abbas, Z.; Chan, H.L.Y.; Chen, C.J.; Chen, D.S.; Chen, H.L.; Chen, P.J.; Chien, R.N.; et al. Asian-Pacific clinical practice guidelines on the management of hepatitis B: A 2015 update. Hepatol. Int. 2016, 10, 1–98. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.L.A.; van Zonneveld, M.; Senturk, H.; Zeuzem, S.; Akarca, U.S.; Cakaloglu, Y.; Simon, C.; So, T.M.K.; Gerken, G.; de Man, R.A.; et al. Pegylated interferon alfa-2b alone or in combination with lamivudine for HBeAg-positive chronic hepatitis B: A randomised trial. Lancet 2005, 365, 123–129. [Google Scholar] [CrossRef]
- Lau, G.K.K.; Piratvisuth, T.; Luo, K.X.; Marcellin, P.; Thongsawat, S.; Cooksley, G.; Gane, E.; Fried, M.W.; Chow, W.C.; Paik, S.W.; et al. Peginterferon alfa-2a, lamivudine, and the combination for HBeAg-positive chronic hepatitis B. N. Engl. J. Med. 2005, 352, 2682–2695. [Google Scholar] [CrossRef] [PubMed]
- Buster, E.H.C.J.; Schalm, S.W.; Janssen, H.L.A. Peginterferon for the treatment of chronic hepatitis B in the era of nucleos(t)ide analogues. Best Pract. Res. Clin. Gastroenterol. 2008, 22, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Bonino, F.; Marcellin, P.; Lau, G.K.K.; Hadziyannis, S.; Jin, R.; Piratvisuth, T.; Germanidis, G.; Yurdaydin, C.; Diago, M.; Gurel, S.; et al. Predicting response to peginterferon alfa-2a, lamivudine and the two combined for HBeAg-negative chronic hepatitis B. Gut 2007, 56, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moucari, R.; Mackiewicz, V.; Lada, O.; Ripault, M.P.; Castelnau, C.; Martinot-Peignoux, M.; Dauvergne, A.; Asselah, T.; Boyer, N.; Bedossa, P.; et al. Early serum HBsAg drop: A strong predictor of sustained virological response to pegylated interferon alfa-2a in HBeAg-negative patients. Hepatology 2009, 49, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Brunetto, M.R.; Moriconi, F.; Bonino, F.; Lau, G.K.K.; Farci, P.; Yurdaydin, C.; Piratvisuth, T.; Luo, K.; Wang, Y.; Hadziyannis, S.; et al. Hepatitis B virus surface antigen levels: A guide to sustained response to peginterferon alfa-2a in HBeAg-negative chronic hepatitis B. Hepatology 2009, 49, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Rijckborst, V.; Hansen, B.E.; Cakaloglu, Y.; Ferenci, P.; Tabak, F.; Akdogan, M.; Simon, K.; Akarca, U.S.; Flisiak, R.; Verhey, E.; et al. Early on-treatment prediction of response to peginterferon alfa-2a for HBeAg-negative chronic hepatitis B using HBsAg and HBV DNA levels. Hepatology 2010, 52, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcellin, P.; Bonino, F.; Yurdaydin, C.; Hadziyannis, S.; Moucari, R.; Kapprell, H.P.; Rothe, V.; Popescu, M.; Brunetto, M.R. Hepatitis B surface antigen levels: Association with 5-year response to peginterferon alfa-2a in hepatitis B e-antigen-negative patients. Hepatol. Int. 2013, 7, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Marcellin, P.; Ahn, S.H.; Ma, X.; Caruntu, F.A.; Tak, W.Y.; Elkashab, M.; Chuang, W.L.; Lim, S.G.; Tabak, F.; Mehta, R.; et al. Combination of tenofovir disoproxil fumarate and peginterferon α-2a increases loss of hepatitis B surface antigen in patients with chronic hepatitis B. Gastroenterology 2016, 150, 134.e10–144.e10. [Google Scholar] [CrossRef] [PubMed]
- Bourlière, M.; Rabiega, P.; Ganne-Carrie, N.; Serfaty, L.; Marcellin, P.; Barthe, Y.; Thabut, D.; Guyader, D.; Hezode, C.; Picon, M.; et al. Effect on HBs antigen clearance of addition of pegylated interferon alfa-2a to nucleos(t)ide analogue therapy versus nucleos(t)ide analogue therapy alone in patients with HBe antigen-negative chronic hepatitis B and sustained undetectable plasma hepatitis B virus DNA: A randomised, controlled, open-label trial. Lancet Gastroenterol. Hepatol. 2017, 2, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Li, W.C.; Wang, M.R.; Kong, L.B.; Ren, W.G.; Zhang, Y.G.; Nan, Y.M. Peginterferon alpha-based therapy for chronic hepatitis B focusing on HBsAg clearance or seroconversion: A meta-analysis of controlled clinical trials. BMC Infect. Dis. 2011, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Lujambio, A.; Lowe, S.W. The microcosmos of cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Morishita, A.; Masaki, T. miRNA in hepatocellular carcinoma. Hepatol. Res. 2015, 45, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Cermelli, S.; Ruggieri, A.; Marrero, J.A.; Ioannou, G.N.; Beretta, L. Circulating microRNAs in patients with chronic hepatitis C and non-alcoholic fatty liver disease. PLoS ONE 2011, 6, e23937. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Asano, M.; Matsumoto, A.; Sugiyama, M.; Nishida, N.; Tanaka, E.; Inoue, T.; Sakamoto, M.; Enomoto, N.; Shirasaki, T.; et al. Induction of IFN-λ3 as an additional effect of nucleotide, not nucleoside, analogues: A new potential target for HBV infection. Gut 2016, 67, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Lampertico, P.; Del Ninno, E.; Vigano, M.; Romeo, R.; Donato, M.F.; Sablon, E.; Morabito, A.; Colombo, M. Long-term suppression of hepatitis B e antigen-negative chronic hepatitis B by 24-month interferon therapy. Hepatology 2003, 37, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Winther, T.N.; Bang-Berthelsen, C.H.; Heiberg, I.L.; Pociot, F.; Hogh, B. Differential plasma microRNA profiles in HBeAg positive and HBeAg negative children with chronic hepatitis B. PLoS ONE 2013, 8, e58236. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ge, G.; Pan, T.; Wen, D.; Gan, J. Serum MiRNA panel as potential biomarkers for chronic hepatitis B with persistently normal alanine aminotransferase. Clin. Chim. Acta 2015, 451, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Brunetto, M.R.; Cavallone, D.; Oliveri, F.; Moriconi, F.; Colombatto, P.; Coco, B.; Ciccorossi, P.; Rastelli, C.; Romagnoli, V.; Cherubini, B.; et al. A serum microRNA signature is associated with the immune control of chronic hepatitis B virus infection. PLoS ONE 2014, 9, e110782. [Google Scholar] [CrossRef] [PubMed]
- Kanlikilicer, P.; Rashed, M.H.; Bayraktar, R.; Mitra, R.; Ivan, C.; Aslan, B.; Zhang, X.; Filant, J.; Silva, A.M.; Rodriguez-Aguayo, C.; et al. Ubiquitous release of exosomal tumor suppressor miR-6126 from Ovarian Cancer Cells. Cancer Res. 2016, 76, 7194–7207. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070.e6–1083.e6. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res. 1982, 42, 3858–3863. [Google Scholar] [PubMed]
- Knowles, B.; Howe, C.; Aden, D. Human hepatocellular carcinoma cell lines secrete the major plasma proteins and hepatitis B surface antigen. Science 1980, 209, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Yotsuyanagi, H.; Sugiyama, M.; Yatsuhashi, H.; Karino, Y.; Takikawa, Y.; Saito, T.; Arase, Y.; Imazeki, F.; Kurosaki, M.; et al. Geographic distribution and characteristics of genotype A hepatitis B virus infection in acute and chronic hepatitis B patients in Japan. J. Gastroenterol. Hepatol. 2016, 31, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Tanaka, Y.; Hige, S.; Yamada, G.; Murawaki, Y.; Komatsu, M.; Kuramitsu, T.; Kawata, S.; Tanaka, E.; Izumi, N.; et al. Distribution of hepatitis B virus genotypes among patients with chronic infection in Japan shifting toward an increase of genotype A. J. Clin. Microbiol. 2009, 47, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, H.; Ujino, S.; Shimizu, Y.; Harada, K.; Zhang, J.; Sugiyama, M.; Mizokami, M.; Shimotohno, K. Novel reporter system to monitor early stages of the hepatitis B virus life cycle. Cancer Sci. 2015, 106, 1616–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachogiannakos, J.; Papatheodoridis, G.V. HBeAg-negative chronic hepatitis B: why do I treat my patients with pegylated interferon-alfa? Liver Int. 2014, 34 (Suppl. 1), 127–132. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.H.; Hsu, C.W.; Chang, M.L.; Chen, Y.C.; Lai, M.W.; Yeh, C.T. Peginterferon is superior to Nucleos(t)ide analogues for prevention of hepatocellular carcinoma in chronic hepatitis B. J. Infect. Dis. 2016, 213, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Sterling, R.K.; Lissen, E.; Clumeck, N.; Sola, R.; Correa, M.C.; Montaner, J.; S. Sulkowski, M.; Torriani, F.J.; Dieterich, D.T.; Thomas, D.L.; et al. Development of a simple noninvasive index to predict significant fibrosis in patients with HIV/HCV coinfection. Hepatology 2006, 43, 1317–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gidlof, O.; van der Brug, M.; Ohman, J.; Gilje, P.; Olde, B.; Wahlestedt, C.; Erlinge, D. Platelets activated during myocardial infarction release functional miRNA, which can be taken up by endothelial cells and regulate ICAM1 expression. Blood 2013, 121, 3908–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age | Sex | Body Weight (kg) | Initial Peg-IFN Dose (μg) | *NUCs | HBsAg (U/mL) | HBV-DNA (Log Copies/mL) | Genotype | ALT (U/mL) | Plate-Let (×109/L) | Fib-4 Index |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 69 | M | 65 | 90 | ETV | 496.83 | <2.1 | C | 23 | 119 | 2.418 |
| 2 | 45 | M | 68 | 180 | ETV | 341.94 | <2.1 | C | 18 | 268 | 0.752 |
| 3 | 57 | M | 73 | 180 | ETV | 475.71 | <2.1 | C | 13 | 190 | 1.581 |
| 4 | 42 | M | 71 | 90 | ETV | 132.36 | <2.1 | C | 62 | 107 | 1.844 |
| 5 | 56 | F | 58 | 180 | ETV | 1255.75 | <2.1 | C | 12 | 193 | 1.423 |
| 6 | 56 | M | 77 | 180 | None | 35.01 | <2.1 | †N.T. | 45 | 239 | 1.048 |
| 7 | 43 | M | 66 | 180 | None | 6179.34 | 4.6 | C | 25 | 240 | 1.003 |
| 8 | 59 | M | 73 | 180 | ETV | 80.13 | <2.1 | C | 16 | 208 | 1.135 |
| 9 | 49 | M | 63 | 90 | LMD + ADV | 851.20 | <2.1 | †N.T. | 27 | 129 | 1.828 |
| 10 | 34 | M | 66 | 180 | None | 52.57 | 3.9 | B | 11 | 183 | 0.780 |
| 11 | 68 | F | 46 | 90 | ETV | 417.89 | <2.1 | C | 16 | 84 | 5.262 |
| 12 | 54 | F | 63 | 90 | ETV | 2261.67 | <2.1 | C | 11 | 158 | 1.855 |
| 13 | 52 | M | 70 | 180 | ETV | 923.92 | <2.1 | C | 22 | 166 | 1.469 |
| 14 | 64 | M | 56 | 90 | ETV | 4528.94 | <2.1+ | B | 23 | 163 | 1.965 |
| 15 | 45 | F | 83 | 180 | None | 45.86 | 2.2 | C | 32 | 127 | 2.722 |
| 16 | 41 | F | 63 | 90 | None | 4786.11 | 2.8 | C | 85 | 164 | 0.957 |
| 17 | 63 | M | 55 | 90 | None | 742.99 | 4.0 | C | 14 | 200 | 1.852 |
| 18 | 51 | F | 58 | 90 | None | 56.51 | 3.2 | C | 11 | 221 | 0.905 |
| 19 | 55 | M | 64 | 90 | None | 58.79 | 3.0 | C | 34 | 198 | 1.286 |
| 20 | 41 | F | 65 | 90 | ETV | 19099.89 | <2.1 | C | 15 | 243 | 0.828 |
| 21 | 66 | F | 44 | 90 | None | 88.81 | 3.4 | C | 15 | 179 | 1.809 |
| 22 | 63 | F | 44 | 90 | None | 5470.72 | 3.8 | C | 14 | 150 | 2.245 |
| Name | Fold-Change | SD | p | Chromosomal Location |
|---|---|---|---|---|
| Up-regulated | ||||
| miR-4787-3p | 2.881 | 1.270 | 0.0003 | 19 |
| miR-6126 | 1.884 | 0.657 | 0.0010 | 16 |
| miR-6769a-3p | 1.602 | 0.378 | 0.0013 | 16 |
| Down-regulated | ||||
| miR-1203 | 0.500 | 0.356 | 0.0015 | 17 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujita, K.; Mimura, S.; Iwama, H.; Nakahara, M.; Oura, K.; Tadokoro, T.; Nomura, T.; Tani, J.; Yoneyama, H.; Morishita, A.; et al. Serum miRNAs Predicting Sustained HBs Antigen Reduction 48 Weeks after Pegylated Interferon Therapy in HBe Antigen-Negative Patients. Int. J. Mol. Sci. 2018, 19, 1940. https://doi.org/10.3390/ijms19071940
Fujita K, Mimura S, Iwama H, Nakahara M, Oura K, Tadokoro T, Nomura T, Tani J, Yoneyama H, Morishita A, et al. Serum miRNAs Predicting Sustained HBs Antigen Reduction 48 Weeks after Pegylated Interferon Therapy in HBe Antigen-Negative Patients. International Journal of Molecular Sciences. 2018; 19(7):1940. https://doi.org/10.3390/ijms19071940
Chicago/Turabian StyleFujita, Koji, Shima Mimura, Hisakazu Iwama, Mai Nakahara, Kyoko Oura, Tomoko Tadokoro, Takako Nomura, Joji Tani, Hirohito Yoneyama, Asahiro Morishita, and et al. 2018. "Serum miRNAs Predicting Sustained HBs Antigen Reduction 48 Weeks after Pegylated Interferon Therapy in HBe Antigen-Negative Patients" International Journal of Molecular Sciences 19, no. 7: 1940. https://doi.org/10.3390/ijms19071940
APA StyleFujita, K., Mimura, S., Iwama, H., Nakahara, M., Oura, K., Tadokoro, T., Nomura, T., Tani, J., Yoneyama, H., Morishita, A., Oryu, M., Himoto, T., Nishitsuji, H., Shimotohno, K., Omata, M., & Masaki, T. (2018). Serum miRNAs Predicting Sustained HBs Antigen Reduction 48 Weeks after Pegylated Interferon Therapy in HBe Antigen-Negative Patients. International Journal of Molecular Sciences, 19(7), 1940. https://doi.org/10.3390/ijms19071940