An Insight into the Difficulties in the Discovery of Specific Biomarkers of Limbal Stem Cells


Abstract
:1. Introduction
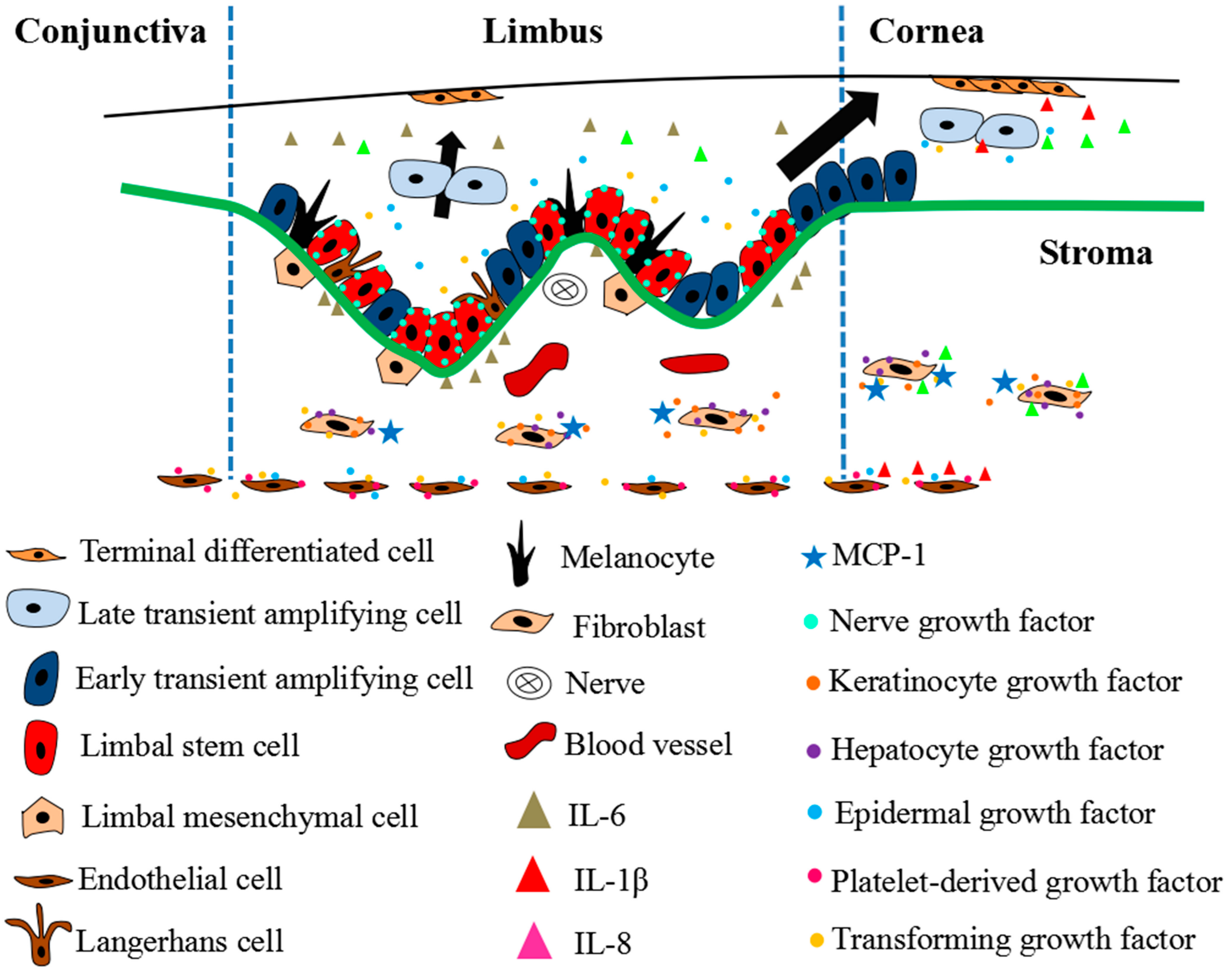
2. LSC and LSC Niche
2.1. Cells
2.2. Cell-Extracellular Matrix
2.3. Cytokines
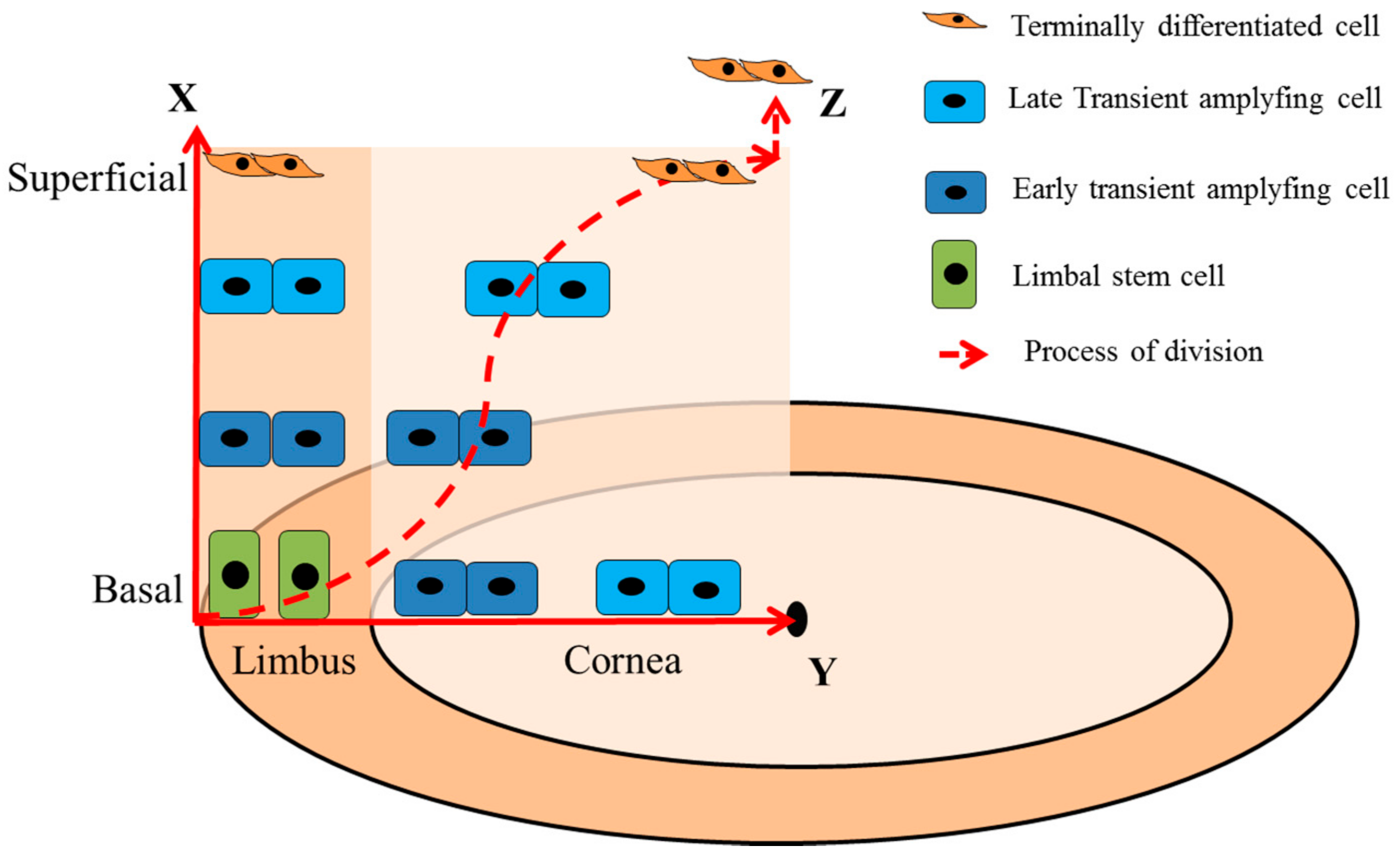
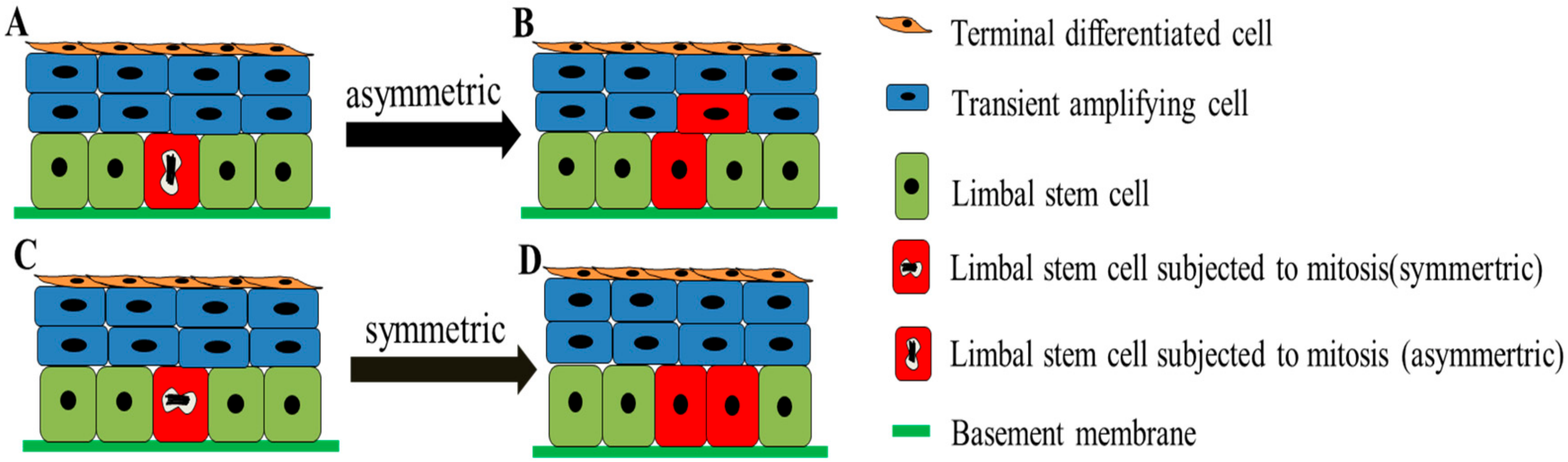
3. Dynamics of LSCs
4. Putative Biomarkers of LSC
4.1. Cell Cycle Regulators
4.1.1. ΔNp63α
4.1.2. C/EBPδ
4.2. ATP-Binding Cassette Transporters
4.2.1. ABCG2
4.2.2. ABCB5
4.3. Cytoskeletal Proteins
4.3.1. CK5 and CK14
4.3.2. Vimentin and Cytokeratin 19
4.4. Differentiation Associated Proteins
4.4.1. Cx 43
4.4.2. CK3 and CK12
5. Identification of LSCs
- The coexpression of LSC biomarkers: such as p63+, ABCG2+, integrin α9+, vimentin+, Cx43−, CK3−/CK12−, and involucrin−, etc.;
- N/C ≥ 0.7: Schlotzer-Schrehardt et al. demonstrated that the ration of N/C of LSC on the BM was higher than TACs and CECs, respectively [59]. Priya et al. successfully identified and quantified LSCs based on ABCG2 and N/C ≥ 0.7 [69]. Additionally, Kasinathan et al. established a two-step protocol by combining basal cell isolation and laser capture microdissection (LCM) of small cells with N/C ≥ 0.7 for LSC enrichment, which achieved 76–78% enrichment of LSC from 2% LSCs in total LECs [104];
- Label-retaining cell: bromodeoxyuridine (BrdU) can take the place of thymidine to be incorporated into the replicated DNA during the S-phase of the cell cycle. The BrdU-based “pulse-chase” experiment has been widely applied for the identification of stem cell. After a period of BrdU pulse, all cells with different degrees of differentiation can be labeled with BrdU. Slow cell cycling is a characteristic of LSCs, which means less division of BrdU labeled LSCs compared to the differentiated cells in the same time. Thus, after a period of BrdU chase, the BrdU-retaining cells can be considered as LSCs [105];
- Side population (SP) phenotype: SP is a sub-population of Hoechst blue−/Hoechst red− cells outside the main population based on staining with DNA-binding dye Hoechst 33342 and isolating by FACS. SP phenotype has become a characteristic of stem cells. Goodell et al. successfully isolated mice bone marrow stem cell by using FACS based on Hoechst-SP method, the Hoechst 33342 efflux activity resulting from the specific protein expression of stem cells, such as ATP binding cassette transporters [80]. Similarly, LSCs showed the ABCG2+/ABCB5+ phenotype. By this method, Shaharuddin et al. successfully isolated the LSCs [106].
6. Difficulties in the Study of LSC-Specific Biomarkers
6.1. Particularity of Limbal Histochemistry
6.2. Influences of Enzyme Digestion
6.3. Lack of Robust LSC Isolation Technology
6.4. Heterogeneity of LSCs
7. Prospectives
7.1. Development of mRNA- or microRNA-Based Biomarkers
7.2. Application of In Situ Sequencing Technology
7.3. Application of the Inducible Transgenic Animal System
7.4. Development of Virtual Simulation Technology
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
| 2-DE | Two-dimensional gel electrophoresis |
| AM | Amniotic membrane |
| bFGF | Basic fibroblast growth factor |
| BM | Basement membrane |
| BrdU: | Bromodeoxyuridine |
| C/EBP | CCAAT/enhancer-binding protein |
| CECs | Corneal epithelial cells |
| CK | Cytokeratin |
| Cx | Connexin |
| EGFR | Epidermal growth factor receptor |
| ESCs | embryonic stem cells |
| FACS | Fluorescence activated cell sorting |
| FFPE | Formalin-fixed paraffin-embedded |
| FGFR-1 | Fibroblast growth factor receptor-1 |
| FGF-β | Fibroblast growth factor-β |
| GJA1 | Gap junction alpha-1 |
| HCC | Hepatocellular carcinoma |
| HE | Hematoxylin and eosin |
| HPCs | Hepatic progenitor cells |
| iPSCs | Induced pluripotent stem cells |
| IGF-I | Insulin-like growth factor-I |
| IGFR | IGF receptor |
| KGF | Keratinocyte growth factor |
| LCM | Laser capture microdissection |
| LECs | Limbal epithelial cells |
| LSCs | Limbal stem cells |
| LSCD | Limbal stem cell deficiency |
| MACS | Magnetic-activated cell sorting |
| NGF | Nerve growth factor |
| NGFR | NGF receptors |
| NGS | Next-generation sequencing |
| N/C | Nuclear-to-cytoplasm |
| OCT | Optimal cutting temperature |
| PDGF | Platelet-derived growth factor |
| PSCs | Pluripotent stem cells |
| POVs | Palisades of Vogt |
| RCA | Rolling circle amplification |
| SD-OCT | Spectral-domain optical coherence tomography |
| SP | Side population |
| TACs | Transient amplifying cells |
| SESCs | Skin epithelial stem cells |
| TDCs | Terminally differentiated cells |
| TGF-α | Transforming growth factor-α |
| WHO | World Health Organization. |
References
- Dziasko, M.A.; Daniels, J.T. Anatomical features and cell-cell interactions in the human limbal epithelial stem cell niche. Ocul. Surf. 2016, 14, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Bourne, R.R.A.; Flaxman, S.R.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; Leasher, J.; Limburg, H.; et al. Magnitude, temporal trends, and projections of the global prevalence of blindness and distance and near vision impairment: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e888–e897. [Google Scholar] [CrossRef]
- Nguyen, K.N.; Bobba, S.; Richardson, A.; Park, M.; Watson, S.L.; Wakefield, D.; Di Girolamo, N. Native and synthetic scaffolds for limbal epithelial stem cell transplantation. Acta Biomater. 2018, 65, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Schermer, A.; Galvin, S.; Sun, T.T. Differentiation-related expression of a major 64 k corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R.M.; Sun, T.T. Epithelial stem cells: The eye provides a vision. Eye 2003, 17, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Shanmuganathan, V.A.; Powell-Richards, A.O.; Tighe, P.J.; Joseph, A. Limbal epithelial crypts: A novel anatomical structure and a putative limbal stem cell niche. Br. J. Ophthalmol. 2005, 89, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Gaddipati, S.; Tian, S.; Xue, L.; Kao, W.W.; Lu, Q.; Li, Q. Study of corneal epithelial progenitor origin and the yap1 requirement using keratin 12 lineage tracing transgenic mice. Sci. Rep. 2016, 6, 35202. [Google Scholar] [CrossRef] [PubMed]
- Ksander, B.R.; Kolovou, P.E.; Wilson, B.J.; Saab, K.R.; Guo, Q.; Ma, J.; McGuire, S.P.; Gregory, M.S.; Vincent, W.J.B.; Perez, V.L.; et al. Abcb5 is a limbal stem cell gene required for corneal development and repair. Nature 2014, 511, 353. [Google Scholar] [CrossRef] [PubMed]
- Di Girolamo, N.; Sarris, M.; Chui, J.; Cheema, H.; Coroneo, M.T.; Wakefield, D. Localization of the low-affinity nerve growth factor receptor p75 in human limbal epithelial cells. J. Cell. Mol. Med. 2008, 12, 2799–2811. [Google Scholar] [CrossRef] [PubMed]
- Barut Selver, O.; Yagci, A.; Egrilmez, S.; Gurdal, M.; Palamar, M.; Cavusoglu, T.; Ates, U.; Veral, A.; Guven, C.; Wolosin, J.M. Limbal stem cell deficiency and treatment with stem cell transplantation. Turkish J. Ophthalmol. 2017, 47, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Spaniol, K.; Witt, J.; Mertsch, S.; Borrelli, M.; Geerling, G.; Schrader, S. Generation and characterisation of decellularised human corneal limbus. Graefe’s Arch. Clin. Exp. Ophthalmol. Albrecht Graefes Arch. Klinische Exp. Ophthalmol. 2018, 256, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Joshi, J.; Farooqui, J.H.; Mathur, U. Results of simple limbal epithelial transplantation in unilateral ocular surface burn. Indian J. Ophthalmol. 2018, 66, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Azuara-Blanco, A. Limbal stem cells of the corneal epithelium. Surv. Ophthalmol. 2000, 44, 415–425. [Google Scholar] [CrossRef]
- Levis, H.J.; Daniels, J.T. Recreating the human limbal epithelial stem cell niche with bioengineered limbal crypts. Curr. Eye Res. 2016, 41, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.S.; Basu, S.; Vemuganti, G.K.; Sejpal, K.; Subramaniam, S.V.; Bandyopadhyay, S.; Krishnaiah, S.; Gaddipati, S.; Tiwari, S.; Balasubramanian, D. Clinical outcomes of xeno-free autologous cultivated limbal epithelial transplantation: A 10-year study. Br. J. Ophthalmol. 2011, 95, 1525–1529. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.S.; Sharp, J.A.H. Simple limbal epithelial transplantation. Curr. Opin. Ophthalmol. 2017, 28, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal stem-cell therapy and long-term corneal regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Latta, L.; Viestenz, A.; Stachon, T.; Colanesi, S.; Szentmary, N.; Seitz, B.; Kasmann-Kellner, B. Human aniridia limbal epithelial cells lack expression of keratins k3 and k12. Exp. Eye Res. 2018, 167, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Mikhailova, A.; Ilmarinen, T.; Ratnayake, A.; Petrovski, G.; Uusitalo, H.; Skottman, H.; Rafat, M. Human pluripotent stem cell-derived limbal epithelial stem cells on bioengineered matrices for corneal reconstruction. Exp. Eye Res. 2016, 146, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.; Arnarsson, A.; Hardarson, T.; Lindgard, A.; Daneshvarnaeini, M.; Ellerstrom, C.; Bruun, A.; Stenevi, U. Transplanting embryonic stem cells onto damaged human corneal endothelium. World J. Stem Cells 2017, 9, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.P.; Han, Y.M.; Cao, X.T. Induced pluripotent stem cell (IPSCS) and their application in immunotherapy. Cell. Mol. Immunol. 2014, 11, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Tian, Y.; Funderburgh, J.; Pellegrini, G.; Zhang, K.; Goldberg, J.L.; Ali, R.R.; Young, M.; Xie, Y.; Temple, S. Regenerating eye tissues to preserve and restore vision. Cell Stem Cell 2018, 22, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sareen, D.; Saghizadeh, M.; Ornelas, L.; Winkler, M.A.; Narwani, K.; Sahabian, A.; Funari, V.A.; Tang, J.; Spurka, L.; Punj, V.; et al. Differentiation of human limbal-derived induced pluripotent stem cells into limbal-like epithelium. Stem Cells Transl. Med. 2014, 3, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Casaroli-Marano, R.P.; Nieto-Nicolau, N.; Martinez-Conesa, E.M.; Edel, M.; B Álvarez-Palomo, A. Potential role of induced pluripotent stem cells (IPSCS) for cell-based therapy of the ocular surface. J. Clin. Med. 2015, 4, 318–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, E.J.; O’Neil, A.L.; Ng, S.Y.; Sun, C.C.; Rubin, L.L. Using intracellular markers to identify a novel set of surface markers for live cell purification from a heterogeneous HIPSC culture. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; de Paiva, C.S.; Luo, L.; Kretzer, F.L.; Pflugfelder, S.C.; Li, D.Q. Characterization of putative stem cell phenotype in human limbal epithelia. Stem Cells 2004, 22, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.C.; Espana, E.M.; Yoo, S.H.; Budak, M.T.; Wolosin, J.M.; Tseng, S.C. Different cell sizes in human limbal and central corneal basal epithelia measured by confocal microscopy and flow cytometry. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5125–5129. [Google Scholar] [CrossRef]
- Lehrer, M.S.; Sun, T.T.; Lavker, R.M. Strategies of epithelial repair: Modulation of stem cell and transit amplifying cell proliferation. J. Cell Sci. 1998, 1119, 2867–2875. [Google Scholar]
- Zhang, Y.; Sun, H.; Liu, Y.; Chen, S.; Cai, S.; Zhu, Y.; Guo, P. The limbal epithelial progenitors in the limbal niche environment. Int. J. Med. Sci. 2016, 13, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Eberwein, P.; Reinhard, T. Concise reviews: The role of biomechanics in the limbal stem cell niche: New insights for our understanding of this structure. Stem Cells 2015, 33, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Higa, K.; Shimmura, S.; Miyashita, H.; Shimazaki, J.; Tsubota, K. Melanocytes in the corneal limbus interact with k19-positive basal epithelial cells. Exp. Eye Res. 2005, 81, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, J.M.; Susskind, D.; Grub, M. The melanocyte and the eye: A review with special emphasis on the cornea. Klinische Monatsblatter Augenheilkd. 2012, 229, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Dziasko, M.A.; Tuft, S.J.; Daniels, J.T. Limbal melanocytes support limbal epithelial stem cells in 2d and 3d microenvironments. Exp. Eye Res. 2015, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R.; Yamato, M.; Sugiyama, H.; Sumide, T.; Yang, J.; Okano, T.; Tano, Y.; Nishida, K. N-cadherin is expressed by putative stem/progenitor cells and melanocytes in the human limbal epithelial stem cell niche. Stem Cells 2007, 25, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Saxena, T.; Kumar, K.; Sen, S.; Tandon, R. Langerhans cell histiocytosis presenting as a limbal nodule in an adult patient. Am. J. Ophthalmol. 2004, 138, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hara, K.; Tian, Q.; Zhao, K.; Yoshitomi, T. Existence of small slow-cycling langerhans cells in the limbal basal epithelium that express abcg2. Exp. Eye Res. 2007, 84, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Hamrah, P.; Huq, S.O.; Liu, Y.; Zhang, Q.; Dana, M.R. Corneal immunity is mediated by heterogeneous population of antigen-presenting cells. J. Leukoc. Biol. 2003, 74, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Gonzalez, S.; Deng, S.X. Extracellular matrix is an important component of limbal stem cell niche. J. Funct. Biomater. 2012, 3, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Al-Aqaba, M.A.; Anis, F.S.; Mohammed, I.; Dua, H.S. Nerve terminals at the human corneoscleral limbus. Br. J. Ophthalmol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.H.; Li, N.Y.; Wu, Z.; Wan, P.X.; Liang, X.W.; Zhang, W.; Wang, X.R.; Li, C.Y.; Xiao, J.H.; Zhou, Q.; et al. Using acellular porcine limbal stroma for rabbit limbal stem cell microenvironment reconstruction. Biomaterials 2011, 32, 7812–7821. [Google Scholar] [CrossRef] [PubMed]
- Espana, E.M.; Kawakita, T.; Romano, A.; Di Pascuale, M.; Smiddy, R.; Liu, C.Y.; Tseng, S.C. Stromal niche controls the plasticity of limbal and corneal epithelial differentiation in a rabbit model of recombined tissue. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5130–5135. [Google Scholar] [CrossRef]
- Higa, K.; Kato, N.; Yoshida, S.; Ogawa, Y.; Shimazaki, J.; Tsubota, K.; Shimmura, S. Aquaporin 1-positive stromal niche-like cells directly interact with n-cadherin-positive clusters in the basal limbal epithelium. Stem Cell Res. 2013, 10, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.T.; Chen, S.Y.; Li, G.G.; Tseng, S.C. Limbal epithelial stem/progenitor cells attract stromal niche cells by sdf-1/cxcr4 signaling to prevent differentiation. Stem Cells 2011, 29, 1874–1885. [Google Scholar] [CrossRef] [PubMed]
- Bray, L.J.; Heazlewood, C.F.; Munster, D.J.; Hutmacher, D.W.; Atkinson, K.; Harkin, D.G. Immunosuppressive properties of mesenchymal stromal cell cultures derived from the limbus of human and rabbit corneas. Cytotherapy 2014, 16, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Kameishi, S.; Sugiyama, H.; Yamato, M.; Sado, Y.; Namiki, H.; Kato, T.; Okano, T. Remodeling of epithelial cells and basement membranes in a corneal deficiency model with long-term follow-up. Lab. Investig. 2015, 95, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Schlotzer-Schrehardt, U.; Dietrich, T.; Saito, K.; Sorokin, L.; Sasaki, T.; Paulsson, M.; Kruse, F.E. Characterization of extracellular matrix components in the limbal epithelial stem cell compartment. Exp. Eye Res. 2007, 85, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Sorokin, L.; Okumura, N.; Koizumi, N.; Kinoshita, S.; Kruse, F.E.; Schlotzer-Schrehardt, U. Laminin-511 and-521-based matrices for efficient ex vivoexpansion of human limbal epithelial progenitor cells. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, J.; Kamiyama, K.; Iguchi, I.; Kita, M.; Sotozono, C.; Kinoshita, S. Growth factors: Importance in wound healing and maintenance of transparency of the cornea. Prog. Retin. Eye Res. 2000, 19, 113–129. [Google Scholar] [CrossRef]
- Qi, H.; Li, D.Q.; Shine, H.D.; Chen, Z.; Yoon, K.C.; Jones, D.B.; Pflugfelder, S.C. Nerve growth factor and its receptor trka serve as potential markers for human corneal epithelial progenitor cells. Exp. Eye Res. 2008, 86, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Tseng, S.C. Three patterns of cytokine expression potentially involved in epithelial-fibroblast interactions of human ocular surface. J. Cell. Physiol. 1995, 163, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Trosan, P.; Svobodova, E.; Chudickova, M.; Krulova, M.; Zajicova, A.; Holan, V. The key role of insulin-like growth factor I in limbal stem cell differentiation and the corneal wound-healing process. Stem Cells Dev. 2012, 21, 3341–3350. [Google Scholar] [CrossRef] [PubMed]
- Honma, Y.; Nishida, K.; Sotozono, C.; Kinoshita, S. Effect of transforming growth factor-beta1 and -beta2 on in vitro rabbit corneal epithelial cell proliferation promoted by epidermal growth factor, keratinocyte growth factor, or hepatocyte growth factor. Exp. Eye Res. 1997, 65, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Thoft, R.A.; Friend, J. The x, y, z hypothesis of corneal epithelial maintenance. Investig. Ophthalmol. Vis. Sci. 1983, 24, 1442–1443. [Google Scholar]
- Lamprecht, J. Symmetric and asymmetric cell division in rat corneal epithelium. Cell Tissue Kinet. 1990, 23, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Lobo, E.P.; Delic, N.C.; Richardson, A.; Raviraj, V.; Halliday, G.M.; Di Girolamo, N.; Myerscough, M.R.; Lyons, J.G. Self-organized centripetal movement of corneal epithelium in the absence of external cues. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.J.; Ismail, S.; Sherwin, T. Limbal stem cells: Central concepts of corneal epithelial homeostasis. World J. Stem Cells 2014, 6, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Schlotzer-Schrehardt, U.; Kruse, F.E. Identification and characterization of limbal stem cells. Exp. Eye Res. 2005, 81, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Ekici, K.; Temelli, O.; Parlakpinar, H.; Samdanci, E.; Polat, A.; Beytur, A.; Tanbek, K.; Ekici, C.; Dursun, I.H. Beneficial effects of aminoguanidine on radiotherapy-induced kidney and testis injury. Andrologia 2016, 48, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Sartaj, R.; Chao, Z.; Wan, P.X.; Pasha, Z.; Liu, A.H.; Fuchs, E.; Luo, Y.C.; Liu, J.B.; Rosenblatt, M. Characterization of slow cycling corneal limbal epithelial cells identifies putative stem cell markers. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3793. [Google Scholar]
- Poli, M.; Janin, H.; Justin, V.; Auxenfans, C.; Burillon, C.; Damour, O. Keratin 13 immunostaining in corneal impression cytology for the diagnosis of limbal stem cell deficiency. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9411–9415. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Kelly, J.G.; Trevisan, J.; Cooper, L.J.; Bentley, A.J.; Carmichael, P.L.; Scott, A.D.; Cotte, M.; Susini, J.; Martin-Hirsch, P.L.; et al. Microspectroscopy of spectral biomarkers associated with human corneal stem cells. Mol. Vis. 2010, 16, 359–368. [Google Scholar] [PubMed]
- Polisetti, N.; Zenkel, M.; Menzel-Severing, J.; Kruse, F.E.; Schlotzer-Schrehardt, U. Cell adhesion molecules and stem cell-niche-interactions in the limbal stem cell niche. Stem Cells 2016, 34, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.A.; Zieske, J.D. The corneal epithelial stem cell niche. Ocul. Surf. 2005, 3, 15–26. [Google Scholar] [CrossRef]
- Mei, H.; Nakatsu, M.N.; Baclagon, E.R.; Deng, S.X. Frizzled 7 maintains the undifferentiated state of human limbal stem/progenitor cells. Stem Cells 2014, 32, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Chee, K.Y.; Kicic, A.; Wiffen, S.J. Limbal stem cells: The search for a marker. Clin. Exp. Ophthalmol. 2006, 34, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, E.; Barbaro, V.; Ruzza, A.; Ponzin, D.; Pellegrini, G.; De Luca, M. Isoforms of delta np63 and the migration of ocular limbal cells in human corneal regeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 9523–9528. [Google Scholar] [CrossRef] [PubMed]
- Priya, C.G.; Prasad, T.; Prajna, N.V.; Muthukkaruppan, V. Identification of human corneal epithelial stem cells on the basis of high abcg2 expression combined with a large n/c ratio. Microsc. Res. Tech. 2013, 76, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, J.; Wang, C.; Tan, D.; Beuerman, R. Human limbal progenitor cell characteristics are maintained in tissue culture. Ann. Acad. Med. 2006, 35, 80–86. [Google Scholar]
- Bishop, J.A.; Teruya-Feldstein, J.; Westra, W.H.; Pelosi, G.; Travis, W.D.; Rekhtman, N. P40 (delta np63) is superior to p63 for the diagnosis of pulmonary squamous cell carcinoma. Modern Pathol. 2012, 25, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yuan, Q.; Niu, M.; Zhang, W.; Wen, L.; Fu, H.; Zhou, F.; He, Z. Transcriptional regulation of p63 on the apoptosis of male germ cells and three stages of spermatogenesis in mice. Cell Death Dis. 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, G.; Dellambra, E.; Golisano, O.; Martinelli, E.; Fantozzi, I.; Bondanza, S.; Ponzin, D.; McKeon, F.; De Luca, M. P63 identifies keratinocyte stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3156–3161. [Google Scholar] [CrossRef] [PubMed]
- Dohn, M.; Zhang, S.; Chen, X. P63alpha and deltanp63alpha can induce cell cycle arrest and apoptosis and differentially regulate p53 target genes. Oncogene 2001, 20, 3193–3205. [Google Scholar] [CrossRef] [PubMed]
- Saghizadeh, M.; Soleymani, S.; Harounian, A.; Bhakta, B.; Troyanovsky, S.M.; Brunken, W.J.; Pellegrini, G.; Ljubimov, A.V. Alterations of epithelial stem cell marker patterns in human diabetic corneas and effects of c-met gene therapy. Mol. Vis. 2011, 17, 2177–2190. [Google Scholar] [PubMed]
- Ramji, D.P.; Foka, P. Ccaat/enhancer-binding proteins: Structure, function and regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.F. Molecular stop signs: Regulation of cell-cycle arrest by c/ebp transcription factors. J. Cell Sci. 2005, 118, 2545–2555. [Google Scholar] [CrossRef] [PubMed]
- Rodier, G.; Montagnoli, A.; Di Marcotullio, L.; Coulombe, P.; Draetta, G.F.; Pagano, M.; Meloche, S. P27 cytoplasmic localization is regulated by phosphorylation on ser10 and is not a prerequisite for its proteolysis. TEMBO J. 2001, 20, 6672–6682. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, V.; Testa, A.; Di Iorio, E.; Mavilio, F.; Pellegrini, G.; De Luca, M. C/ebpdelta regulates cell cycle and self-renewal of human limbal stem cells. J. Cell Biol. 2007, 177, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Goodell, M.A. Stem cell identification and sorting using the Hoechst 33342 side population (sp). In Current Protocols in Cytometry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; pp. 9–18. [Google Scholar]
- De Paiva, C.S.; Chen, Z.; Corrales, R.M.; Pflugfelder, S.C.; Li, D.Q. Abcg2 transporter identifies a population of clonogenic human limbal epithelial cells. Stem Cells 2005, 23, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Taghi-Abadi, E.; Baharvand, H. Limbal stem cells in review. J. Ophthalmic Vis. Res. 2009, 4, 40–58. [Google Scholar] [PubMed]
- Wilson, B.J.; Schatton, T.; Zhan, Q.; Gasser, M.; Ma, J.; Saab, K.R.; Schanche, R.; Waaga-Gasser, A.M.; Gold, J.S.; Huang, Q.; et al. Abcb5 identifies a therapy-refractory tumor cell population in colorectal cancer patients. Cancer Res. 2011, 71, 5307–5316. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.; Lobo, E.P.; Delic, N.C.; Myerscough, M.R.; Lyons, J.G.; Wakefield, D.; Di Girolamo, N. Keratin-14-positive precursor cells spawn a population of migratory corneal epithelia that maintain tissue mass throughout life. Stem Cell Rep. 2017, 9, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Allinson, S.L.; Ma, A.; Bentley, A.J.; Martin, F.L.; Fullwood, N.J. Targeted cornea limbal stem/progenitor cell transfection in an organ culture model. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3395–3401. [Google Scholar] [CrossRef] [PubMed]
- Figueira, E.C.; Di Girolamo, N.; Coroneo, M.T.; Wakefield, D. The phenotype of limbal epithelial stem cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Meller, D.; Pires, R.T.; Tseng, S.C. Ex vivo preservation and expansion of human limbal epithelial stem cells on amniotic membrane cultures. Br. J. Ophthalmol. 2002, 86, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Mi, S.L.; Wright, B.; Connon, C.J. Investigation of k14/k5 as a stem cell marker in the limbal region of the bovine cornea. PLoS ONE 2010, 5, e13192. [Google Scholar] [CrossRef] [PubMed]
- Govaere, O.; Komuta, M.; Berkers, J.; Spee, B.; Janssen, C.; de Luca, F.; Katoonizadeh, A.; Wouters, J.; van Kempen, L.C.; Durnez, A.; et al. Keratin 19: A key role player in the invasion of human hepatocellular carcinomas. Gut 2014, 63, 674–685. [Google Scholar] [CrossRef] [PubMed]
- Larouche, D.; Lavoie, A.; Paquet, C.; Simard-Bisson, C.; Germain, L. Identification of epithelial stem cells in vivo and in vitro using keratin 19 and brdu. Methods Mol. Biol. 2010, 585, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Larouche, D.; Hayward, C.; Cuffley, K.; Germain, L. Keratin 19 as a stem cell marker in vivo and in vitro. Methods Mol. Biol. 2005, 289, 103–110. [Google Scholar] [PubMed]
- Ramirez-Miranda, A.; Nakatsu, M.N.; Zarei-Ghanavati, S.; Nguyen, C.V.; Deng, S.X. Keratin 13 is a more specific marker of conjunctival epithelium than keratin 19. Mol. Vis. 2011, 17, 1652–1661. [Google Scholar] [PubMed]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, J.E.; Dechat, T.; Grin, B.; Helfand, B.; Mendez, M.; Pallari, H.M.; Goldman, R.D. Introducing intermediate filaments: From discovery to disease. J. Clin. Investig. 2009, 119, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Vuoriluoto, K.; Haugen, H.; Kiviluoto, S.; Mpindi, J.P.; Nevo, J.; Gjerdrum, C.; Tiron, C.; Lorens, J.B.; Ivaska, J. Vimentin regulates emt induction by slug and oncogenic H-RAS and migration by governing axl expression in breast cancer. Oncogene 2011, 30, 1436–1448. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.G.; Kojima, S.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, J.M.; Bayless, K.J. Vimentin as an integral regulator of cell adhesion and endothelial sprouting. Microcirculation 2014, 21, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Ivaska, J.; Pallari, H.M.; Nevo, J.; Eriksson, J.E. Novel functions of vimentin in cell adhesion, migration, and signaling. Exp. Cell Res. 2007, 313, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Ghoubay-Benallaoua, D.; Basli, E.; Goldschmidt, P.; Pecha, F.; Chaumeil, C.; Laroche, L.; Borderie, V. Human epithelial cell cultures from superficial limbal explants. Mol. Vis. 2011, 17, 341–354. [Google Scholar] [PubMed]
- Donaldson, P.; Eckert, R.; Green, C.; Kistler, J. Gap junction channels: New roles in disease. Histol. Histopathol. 1997, 12, 219–231. [Google Scholar] [PubMed]
- Sohl, G.; Willecke, K. Gap junctions and the connexin protein family. Cardiovasc. Res. 2004, 62, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Evans, W.H.; Pflugfelder, S.C.; Li, D.Q. Gap junction protein connexin 43 serves as a negative marker for a stem cell-containing population of human limbal epithelial cells. Stem Cells 2006, 24, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Tanifuji-Terai, N.; Terai, K.; Hayashi, Y.; Chikama, T.; Kao, W.W. Expression of keratin 12 and maturation of corneal epithelium during development and postnatal growth. Investig. Ophthalmol. Vis. Sci. 2006, 47, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Kasinathan, J.R.; Namperumalsamy, V.P.; Veerappan, M.; Chidambaranathan, G.P. A novel method for a high enrichment of human corneal epithelial stem cells for genomic analysis. Microsc. Res. Tech. 2016, 79, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Arpitha, P.; Prajna, N.V.; Srinivasan, M.; Muthukkaruppan, V. A subset of human limbal epithelial cells with greater nucleus-to-cytoplasm ratio expressing high levels of p63 possesses slow-cycling property. Cornea 2008, 27, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Shaharuddin, B.; Harvey, I.; Ahmad, S.; Ali, S.; Meeson, A. Characterisation of human limbal side population cells isolated using an optimised protocol from an immortalised epithelial cell line and primary limbal cultures. Stem Cell. Rev. Rep. 2014, 10, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, I.; Maddileti, S.; Savy, S.; Tiwari, S.; Gaddipati, S.; Fatima, A.; Sangwan, V.S.; Balasubramanian, D.; Vemuganti, G.K. In vitro culture and expansion of human limbal epithelial cells. Nat. Protoc. 2010, 5, 1470–1479. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Mui, M.M.; Kao, W.W.; Liu, C.Y.; Tseng, S.C. Conjunctival epithelial cells do not transdifferentiate in organotypic cultures: Expression of k12 keratin is restricted to corneal epithelium. Curr. Eye Res. 1994, 13, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Puangsricharern, V.; Tseng, S.C. Cytologic evidence of corneal diseases with limbal stem cell deficiency. Ophthalmology 1995, 102, 1476–1485. [Google Scholar] [CrossRef]
- Poli, M.; Burillon, C.; Auxenfans, C.; Rovere, M.R.; Damour, O. Immunocytochemical diagnosis of limbal stem cell deficiency: Comparative analysis of current corneal and conjunctival biomarkers. Cornea 2015, 34, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Donisi, P.M.; Rama, P.; Fasolo, A.; Ponzin, D. Analysis of limbal stem cell deficiency by corneal impression cytology. Cornea 2003, 22, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Shirzadeh, E.; Keshel, S.H.; Ezzatizadeh, V.; Jabbehdari, S.; Baradaran-Rafii, A. Unrestricted somatic stem cells, as a novel feeder layer: Ex vivo culture of human limbal stem cells. J. Cell. Biochem. 2018, 119, 2666–2678. [Google Scholar] [CrossRef] [PubMed]
- Espana, E.M.; Romano, A.C.; Kawakita, T.; Di Pascuale, M.; Smiddy, R.; Tseng, S.C. Novel enzymatic isolation of an entire viable human limbal epithelial sheet. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4275–4281. [Google Scholar] [CrossRef]
- Chen, S.Y.; Hayashida, Y.; Chen, M.Y.; Xie, H.T.; Tseng, S.C. A new isolation method of human limbal progenitor cells by maintaining close association with their niche cells. Tissue Eng. Part C Methods 2011, 17, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Stasi, K.; Goings, D.; Huang, J.; Herman, L.; Pinto, F.; Addis, R.C.; Klein, D.; Massaro-Giordano, G.; Gearhart, J.D. Optimal isolation and xeno-free culture conditions for limbal stem cell function. Investig. Ophthalmol. Vis. Sci. 2014, 55, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Paniagua, M.; Nieto-Miguel, T.; de la Mata, A.; Dziasko, M.; Galindo, S.; Rey, E.; Herreras, J.M.; Corrales, R.M.; Daniels, J.T.; Calonge, M. Comparison of functional limbal epithelial stem cell isolation methods. Exp. Eye Res. 2016, 146, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Schofield, R. The stem cell system. Biomed. Pharmacother. Biomed. Pharmacother. 1983, 37, 375–380. [Google Scholar] [PubMed]
- Atallah, M.R.; Palioura, S.; Perez, V.L.; Amescua, G. Limbal stem cell transplantation: Current perspectives. Clin. Ophthalmol. 2016, 10, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Haagdorens, M.; Van Acker, S.I.; Van Gerwen, V.; Ni Dhubhghaill, S.; Koppen, C.; Tassignon, M.J.; Zakaria, N. Limbal stem cell deficiency: Current treatment options and emerging therapies. Stem Cells Int. 2016, 2016, 9798374. [Google Scholar] [CrossRef] [PubMed]
- Krulova, M.; Pokorna, K.; Lencova, A.; Fric, J.; Zajicova, A.; Filipec, M.; Forrester, J.V.; Holan, V. A rapid separation of two distinct populations of mouse corneal epithelial cells with limbal stem cell characteristics by centrifugation on percoll gradient. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3903–3908. [Google Scholar] [CrossRef] [PubMed]
- Shaharuddin, B.; Ahmad, S.; Latar, N.M.; Ali, S.; Meeson, A. A human corneal epithelial cell line model for limbal stem cell biology and limbal immunobiology. Stem Cells Transl. Med. 2017, 6, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Espina, V.; Wulfkuhle, J.D.; Calvert, V.S.; VanMeter, A.; Zhou, W.; Coukos, G.; Geho, D.H.; Petricoin, E.F., 3rd; Liotta, L.A. Laser-capture microdissection. Nat. Protoc. 2006, 1, 586–603. [Google Scholar] [CrossRef] [PubMed]
- Emmert-Buck, M.R.; Bonner, R.F.; Smith, P.D.; Chuaqui, R.F.; Zhuang, Z.; Goldstein, S.R.; Weiss, R.A.; Liotta, L.A. Laser capture microdissection. Science 1996, 274, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Espina, V.; Heiby, M.; Pierobon, M.; Liotta, L.A. Laser capture microdissection technology. Expert Rev. Mol. Diagn. 2007, 7, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Bath, C.; Muttuvelu, D.; Emmersen, J.; Vorum, H.; Hjortdal, J.; Zachar, V. Transcriptional dissection of human limbal niche compartments by massive parallel sequencing. PLoS ONE 2013, 8, e64244. [Google Scholar] [CrossRef]
- Datta, S.; Malhotra, L.; Dickerson, R.; Chaffee, S.; Sen, C.K.; Roy, S. Laser capture microdissection: Big data from small samples. Histol. Histopathol. 2015, 30, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Fend, F.; Emmert-Buck, M.R.; Chuaqui, R.; Cole, K.; Lee, J.; Liotta, L.A.; Raffeld, M. Immuno-lcm: Laser capture microdissection of immunostained frozen sections for MRNA analysis. Am. J. Pathol. 1999, 154, 61–66. [Google Scholar] [CrossRef]
- Gautam, V.; Singh, A.; Singh, S.; Sarkar, A.K. An efficient lcm-based method for tissue specific expression analysis of genes and MIRNAs. Sci. Rep. 2016, 6, 21577. [Google Scholar] [CrossRef] [PubMed]
- Craven, R.A.; Totty, N.; Harnden, P.; Selby, P.J.; Banks, R.E. Laser capture microdissection and two-dimensional polyacrylamide gel electrophoresis—Evaluation of tissue preparation and sample limitations. Am. J. Pathol. 2002, 160, 815–822. [Google Scholar] [CrossRef]
- Liu, A. Laser capture microdissection in the tissue biorepository. J. Biomol. Tech. 2010, 21, 120–125. [Google Scholar] [PubMed]
- Luo, Y.; He, J.; Xu, X.; Sun, M.A.; Wu, X.; Lu, X.; Xie, H. Integrative single-cell omics analyses reveal epigenetic heterogeneity in mouse embryonic stem cells. PLoS Comput. Biol. 2018, 14, e1006034. [Google Scholar] [CrossRef] [PubMed]
- Du, W.J.; Chi, Y.; Yang, Z.X.; Li, Z.J.; Cui, J.J.; Song, B.Q.; Li, X.; Yang, S.G.; Han, Z.B.; Han, Z.C. Heterogeneity of proangiogenic features in mesenchymal stem cells derived from bone marrow, adipose tissue, umbilical cord, and placenta. Stem Cell Res. Ther. 2016, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, Y.; Li, W.; Chen, Y.T.; He, H.; Chen, S.Y.; Kheirkah, A.; Zhu, Y.T.; Matsumoto, Y.; Tseng, S.C. Heterogeneity of limbal basal epithelial progenitor cells. Cornea 2010, 29 (Suppl. 1), S32–S40. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Tew, T.B.; Hsieh, Y.T.; Lin, S.Y.; Chang, H.W.; Hu, F.R.; Chen, W.L. Using optical coherence tomography to assess the role of age and region in corneal epithelium and palisades of vogt. Medicine 2016, 95. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Shortt, A.J.; O’Callaghan, A.R.; Daniels, J.T. The impact of age on the physical and cellular properties of the human limbal stem cell niche. Age 2013, 35, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Grieve, K.; Ghoubay, D.; Georgeon, C.; Thouvenin, O.; Bouheraoua, N.; Paques, M.; Borderie, V.M. Three-dimensional structure of the mammalian limbal stem cell niche. Exp. Eye Res. 2015, 140, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the limbal epithelial stem cell niche: Novel imaging techniques permit in vivo observation and targeted biopsy of limbal epithelial stem cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.R.; Daniels, J.T. Concise review: Limbal epithelial stem cell therapy: Controversies and challenges. Stem Cells 2011, 29, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Mo, V.; Nagasaki, T. Distribution of label-retaining cells in the limbal epithelium of a mouse eye. J. Histochem. Cytochem. 2009, 57, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Howlett, P.; Cleal, J.K.; Wu, H.; Shah, N.; Horton, A.; Curzen, N.; Mahmoudi, M. Microrna 8059 as a marker for the presence and extent of coronary artery calcification. Open Heart 2018, 5, e000678. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Lu, S.; Dong, L.; Xue, Y.; Yao, C.; Tong, C.; Wang, C.; Shu, X. Hsa-mir-320d and hsa-mir-582, mirna biomarkers of aortic dissection regulate apoptosis of vascular smooth muscle cells. J. Cardiovasc. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ou, Q.; Braciak, T.A.; Chen, G.; Oduncu, F.S. Microrna-192-5p is a predictive biomarker of survival for stage IIIB colon cancer patients. Jpn. J. Clin. Oncol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Mignardi, M.; Pacureanu, A.; Svedlund, J.; Botling, J.; Wahlby, C.; Nilsson, M. In situ sequencing for RNA analysis in preserved tissue and cells. Nat. Methods 2013, 10, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Sartaj, R.; Zhang, C.; Wan, P.; Pasha, Z.; Guaiquil, V.; Liu, A.; Liu, J.; Luo, Y.; Fuchs, E.; Rosenblatt, M.I. Characterization of slow cycling corneal limbal epithelial cells identifies putative stem cell markers. Sci. Rep. 2017, 7, 3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasser, W.; Amitai-Lange, A.; Soteriou, D.; Hanna, R.; Tiosano, B.; Fuchs, Y.; Shalom-Feuerstein, R. Corneal-committed cells restore the stem cell pool and tissue boundary following injury. Cell Rep. 2018, 22, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Mirulla, A.I.; Bragonzoni, L.; Zaffagnini, S.; Bontempi, M.; Nigrelli, V.; Ingrassia, T. Virtual simulation of an osseointegrated trans-humeral prosthesis: A falling scenario. Injury 2018. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.; Jung, S.W.; Park, H.S. In vivo biomechanical measurement and haptic simulation of portal placement procedure in shoulder arthroscopic surgery. PLoS ONE 2018, 13, e0193736. [Google Scholar] [CrossRef] [PubMed]
- Molvaer, R.K.; Andreasen, A.; Heegaard, S.; Thomsen, J.S.; Hjortdal, J.; Urbak, S.F.; Nielsen, K. Interactive 3d computer model of the human corneolimbal region: Crypts, projections and stem cells. Acta Ophthalmol. 2013, 91, 457–462. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| BM Components | Cornea | Limbus | References |
|---|---|---|---|
| α1(IV) collagen chain | − | ++ | [47] |
| α2(IV) collagen chain | − | ++ | [47] |
| α3(IV) collagen chain | − | − | [47] |
| α4(IV) collagen chain | − | + | [47] |
| α5(IV) collagen chain | ++ | ++ | [47] |
| α6(IV) collagen chain | ++ | ++ | [48] |
| Type V collagen | + | − | [48] |
| Type VI collagen | − | − | [48] |
| Type VII collagen | ++ | ++ | [48] |
| Laminin α1chain | + | + | [49] |
| Laminin α2 chain | ± | + | [49] |
| Laminin α3 chain | ++ | ++ | [49] |
| Laminin α4 chain | − | ± | [49] |
| Laminin α5 chain | − | + | [49] |
| Laminin β1 chain | ++ | ++ | [49] |
| Laminin β2 chain | + | ++ | [49] |
| Laminin β3 chain | + | ++ | [49] |
| Laminin γ1 chain | + | ++ | [49] |
| Laminin γ2 chain | + | ++ | [49] |
| Laminin γ3 chain | ± | + | [49] |
| Group of Putative Biomarkers | Putative Biomarkers | Corneal | Limbal | References | ||
|---|---|---|---|---|---|---|
| Basal | Suprabasal | Basal | Suprabasal | |||
| Cell structural proteins | Vimentin | − | − | ++ | + | [59] |
| CK5/14 | − | − | + | + | [61] | |
| CK19 | +++ | +++ | +++ | + | [62] | |
| CK15 | + | − | ++ | − | [63] | |
| Cell adhesion molecules | Integrinα2 | +++ | +++ | +++ | +++ | [64] |
| Integrinα3 | +++ | + | +++ | ± | [64] | |
| Integrinα4 | − | − | + | ± | [64] | |
| Integrinα6 | ++ | + | − | +++ | [64] | |
| Integrinα8 | − | − | ± | ± | [64] | |
| Integrinα9 | − | − | +++ | ± | [65] | |
| Integrinβ1 | +++ | ++ | +++ | + | [27] | |
| Integrinβ4 | ++ | + | − | + | [59] | |
| P−cadherin | ± | − | ± | − | [59] | |
| E−cadherin | + | +++ | − | +++ | [36] | |
| N−cadherin | − | − | + | ± | [36] | |
| Frizzled7 | + | − | +++ | ++ | [66] | |
| Enzymes | α−enolase | ++ | + | +++ | + | [65] |
| cytochrome oxidase | ++ | + | +++ | + | [67] | |
| Na+/K+−ATPase | ++ | + | +++ | + | [67] | |
| Growth factors and its receptors | EGF−R | +++ | +++ | +++ | + | [27] |
| KGF−R bek | ± | − | − | − | [59] | |
| NGF−R TrkA | ± | − | + | − | [51] | |
| NGF−R p75 | ++ | − | ++ | − | [51] | |
| NGF | + | ± | ++ | − | [51] | |
| Cell cycle regulators | ΔNp63α | − | − | +++ | ± | [68] |
| ATP−binding cassette transporters | ABCG2 | − | − | +++ | ± | [69] |
| ABCB5 | − | − | +++ | ++ | [8] | |
| Differentiation associated proteins | Cx43 | + | +++ | − | +++ | [69] |
| CK3/12 | +++ | +++ | − | +++ | [59] | |
| Involucrin | + | +++ | − | +++ | [70] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.H.; Zhang, W.; Jia, Y.Y.S.; Liu, Q.X.; Li, Z.F.; Lin, J.S. An Insight into the Difficulties in the Discovery of Specific Biomarkers of Limbal Stem Cells. Int. J. Mol. Sci. 2018, 19, 1982. https://doi.org/10.3390/ijms19071982
Guo ZH, Zhang W, Jia YYS, Liu QX, Li ZF, Lin JS. An Insight into the Difficulties in the Discovery of Specific Biomarkers of Limbal Stem Cells. International Journal of Molecular Sciences. 2018; 19(7):1982. https://doi.org/10.3390/ijms19071982
Chicago/Turabian StyleGuo, Zhi Hou, Wei Zhang, Yang Yan Sheng Jia, Qing Xiu Liu, Zhao Fa Li, and Jun Sheng Lin. 2018. "An Insight into the Difficulties in the Discovery of Specific Biomarkers of Limbal Stem Cells" International Journal of Molecular Sciences 19, no. 7: 1982. https://doi.org/10.3390/ijms19071982
APA StyleGuo, Z. H., Zhang, W., Jia, Y. Y. S., Liu, Q. X., Li, Z. F., & Lin, J. S. (2018). An Insight into the Difficulties in the Discovery of Specific Biomarkers of Limbal Stem Cells. International Journal of Molecular Sciences, 19(7), 1982. https://doi.org/10.3390/ijms19071982